The Reaction of Oxy Hemoglobin with Nitrite: Mechanism, Antioxidant-Modulated Effect, and Implications for Blood Substitute Evaluation



Abstract
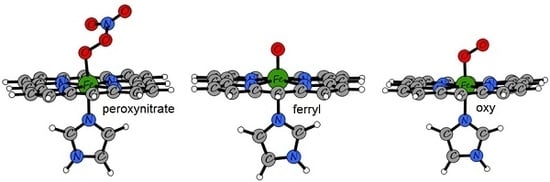
:
1. Introduction
2. Results and Discussion
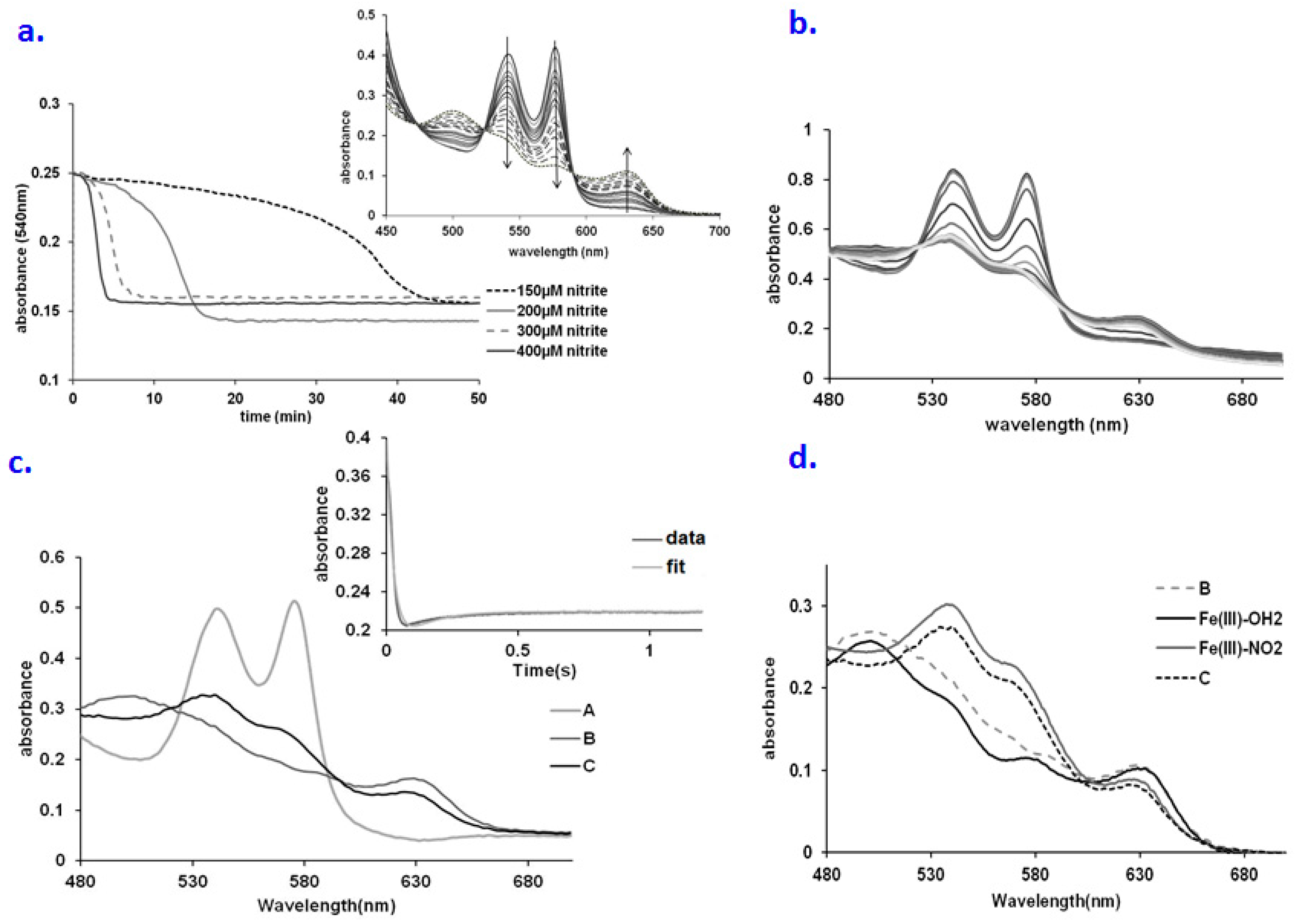
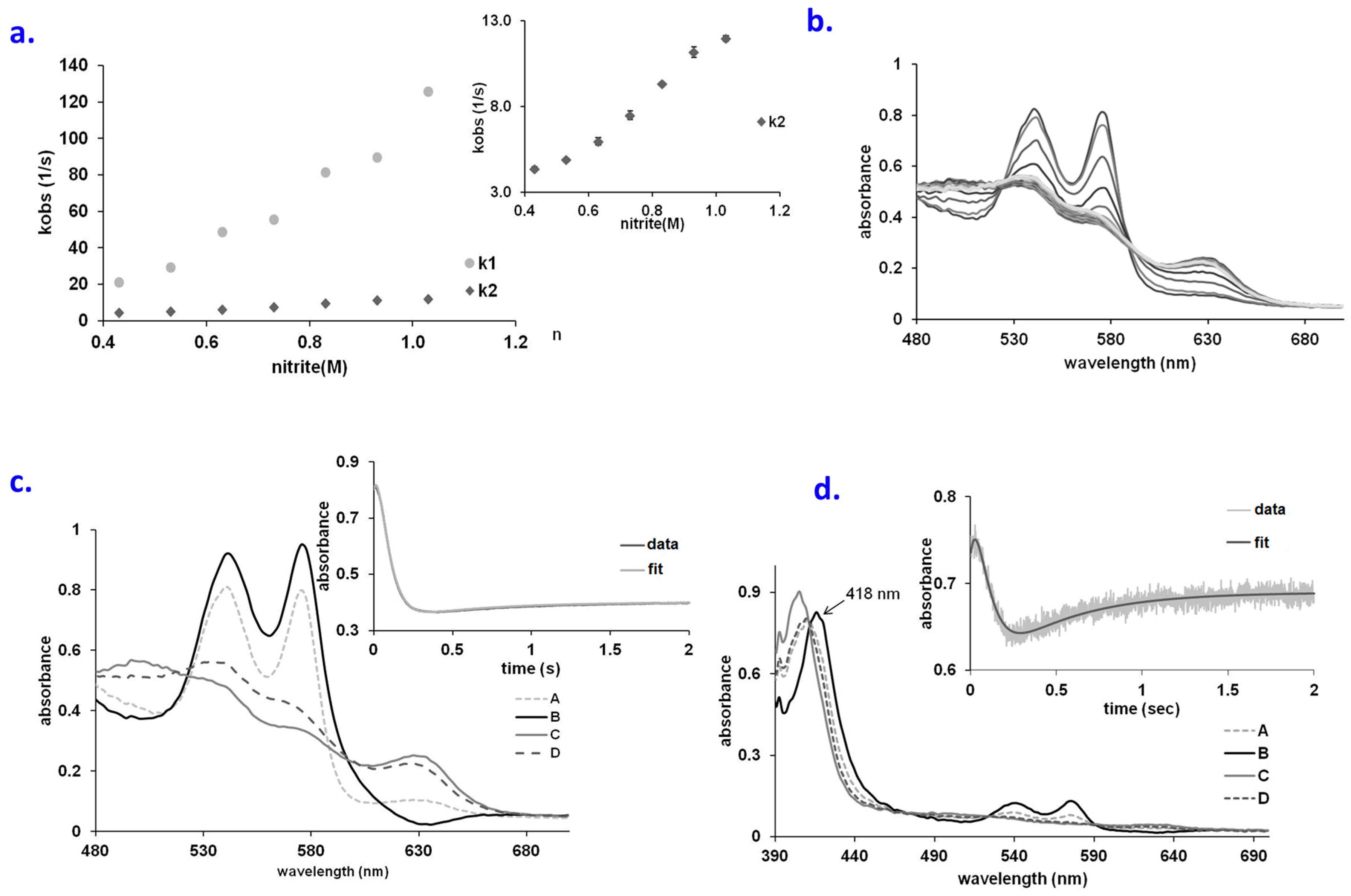
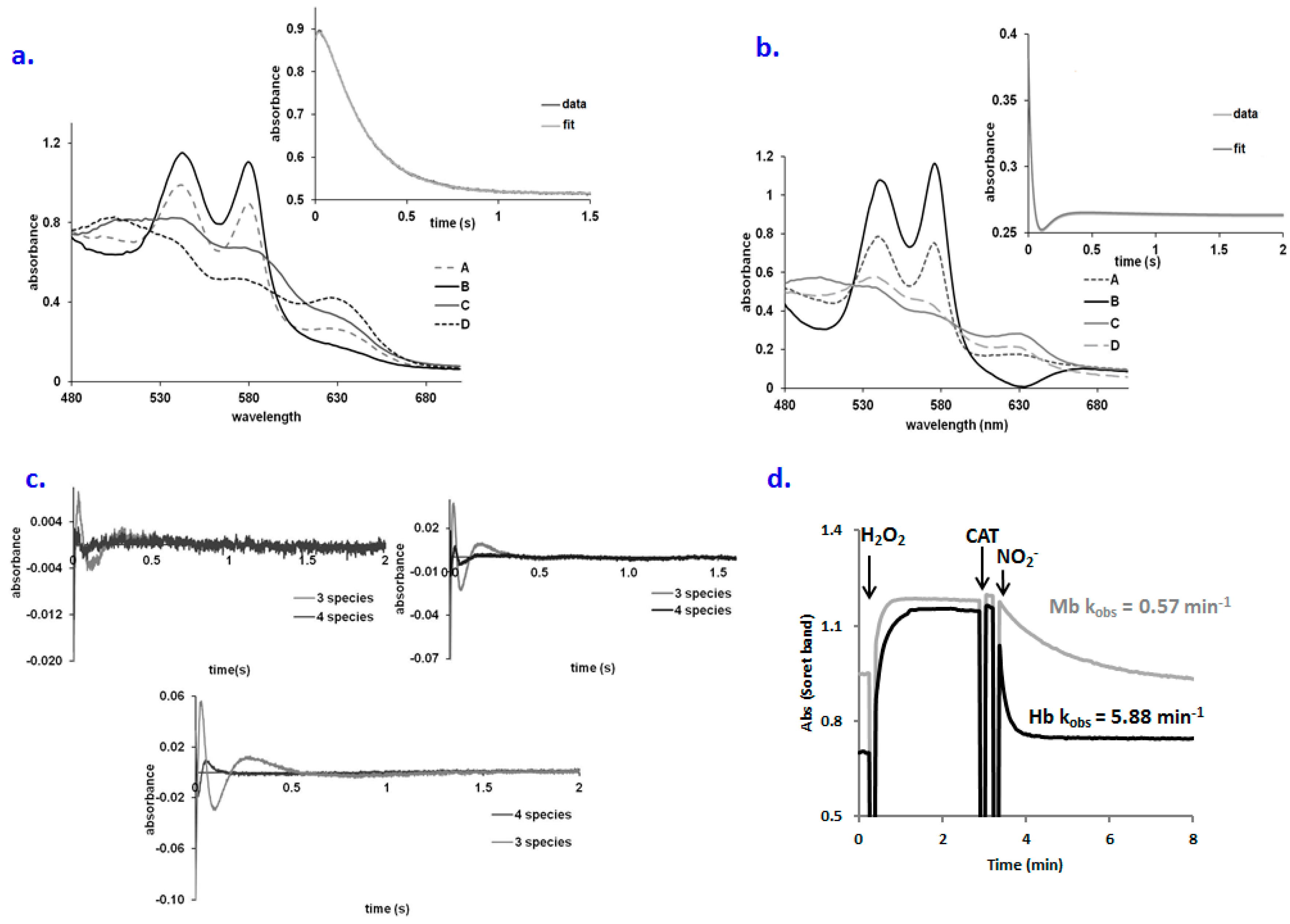
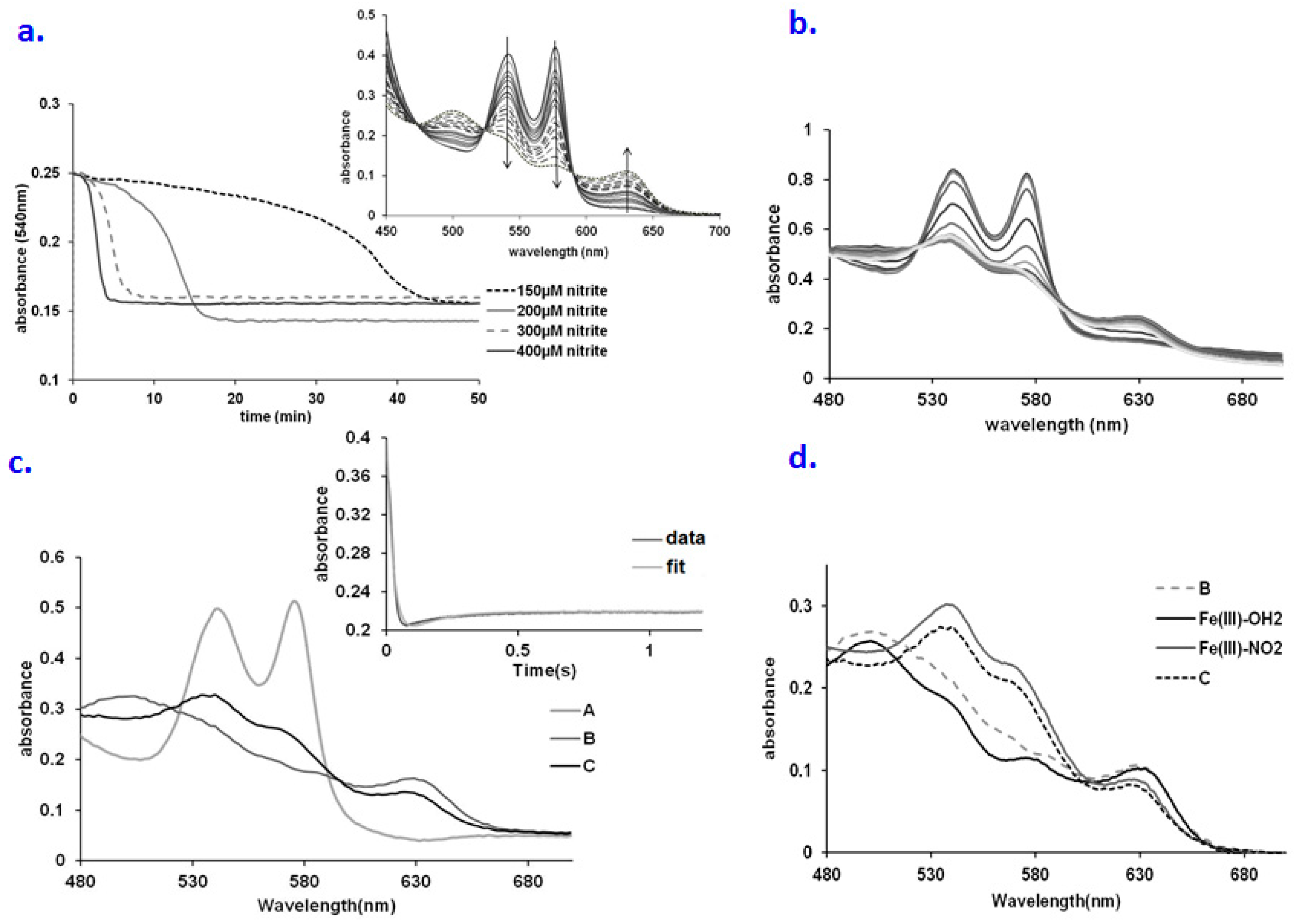
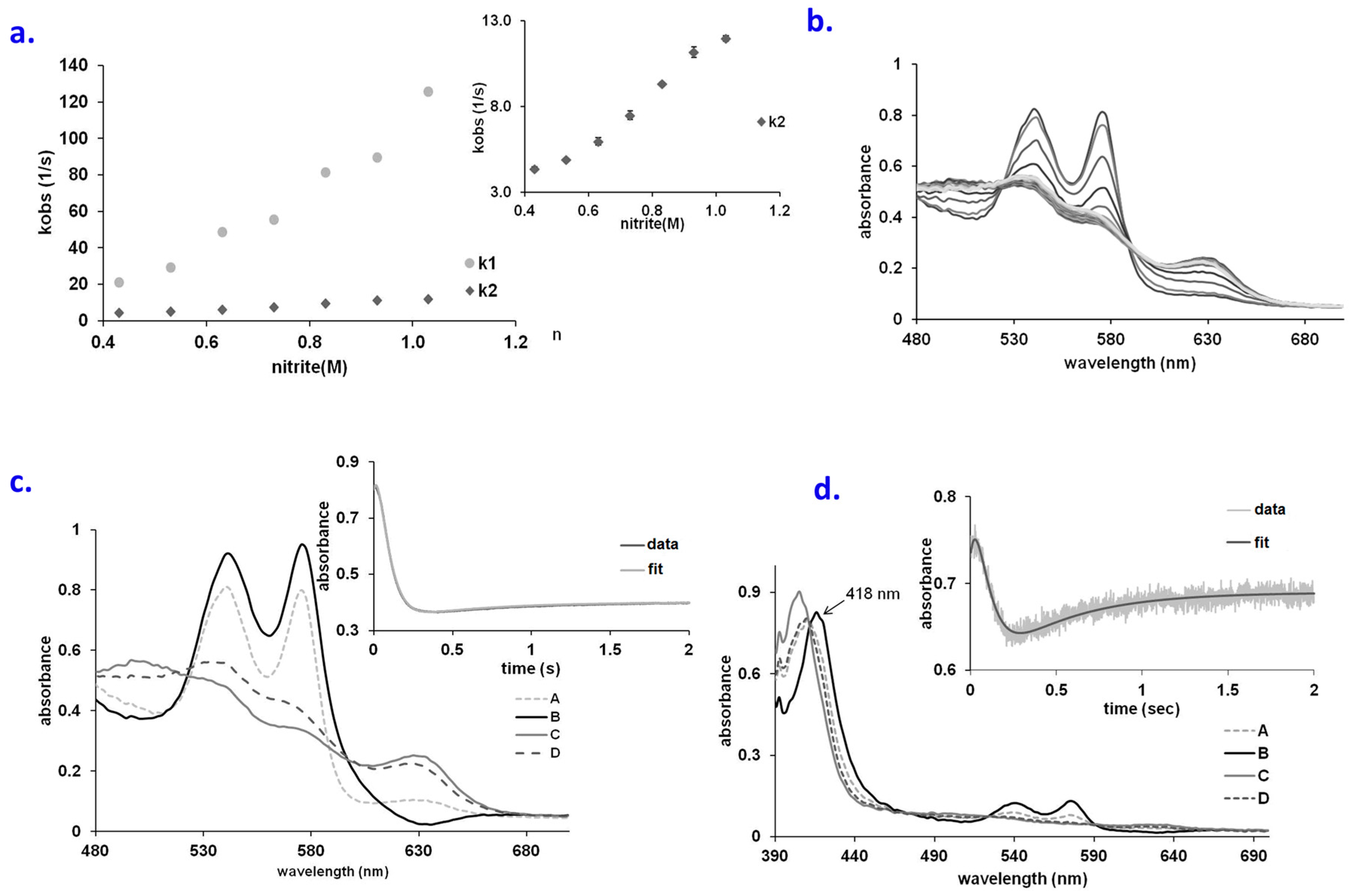
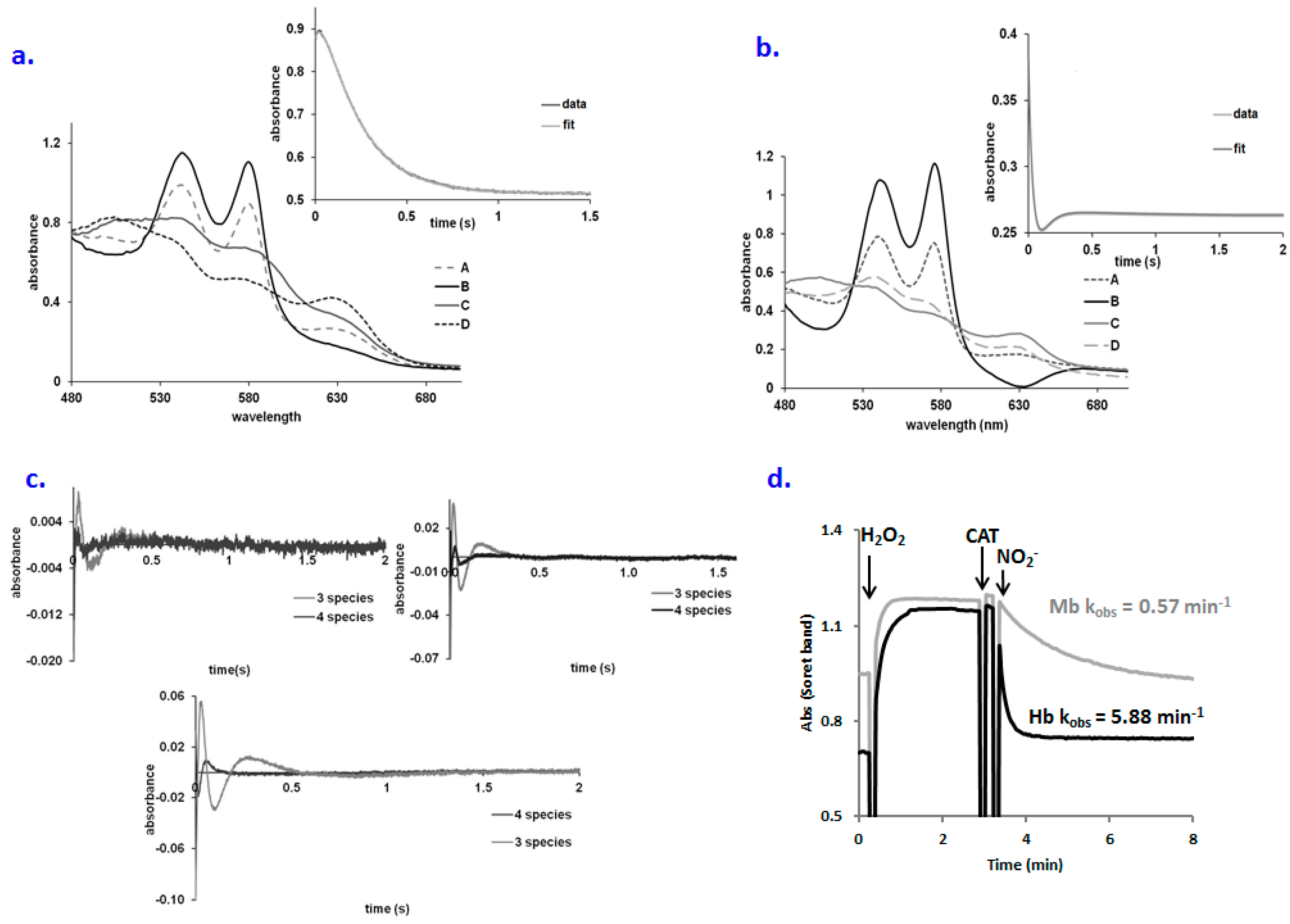
2.1. No Significant Accumulation of Ferryl in the OxyHb–Nitrite Reaction at Supraphysiological Nitrite Concentration
2.2. An Elusive Minor Component in the Kinetics: The Heme-Peroxynitrate Adduct?
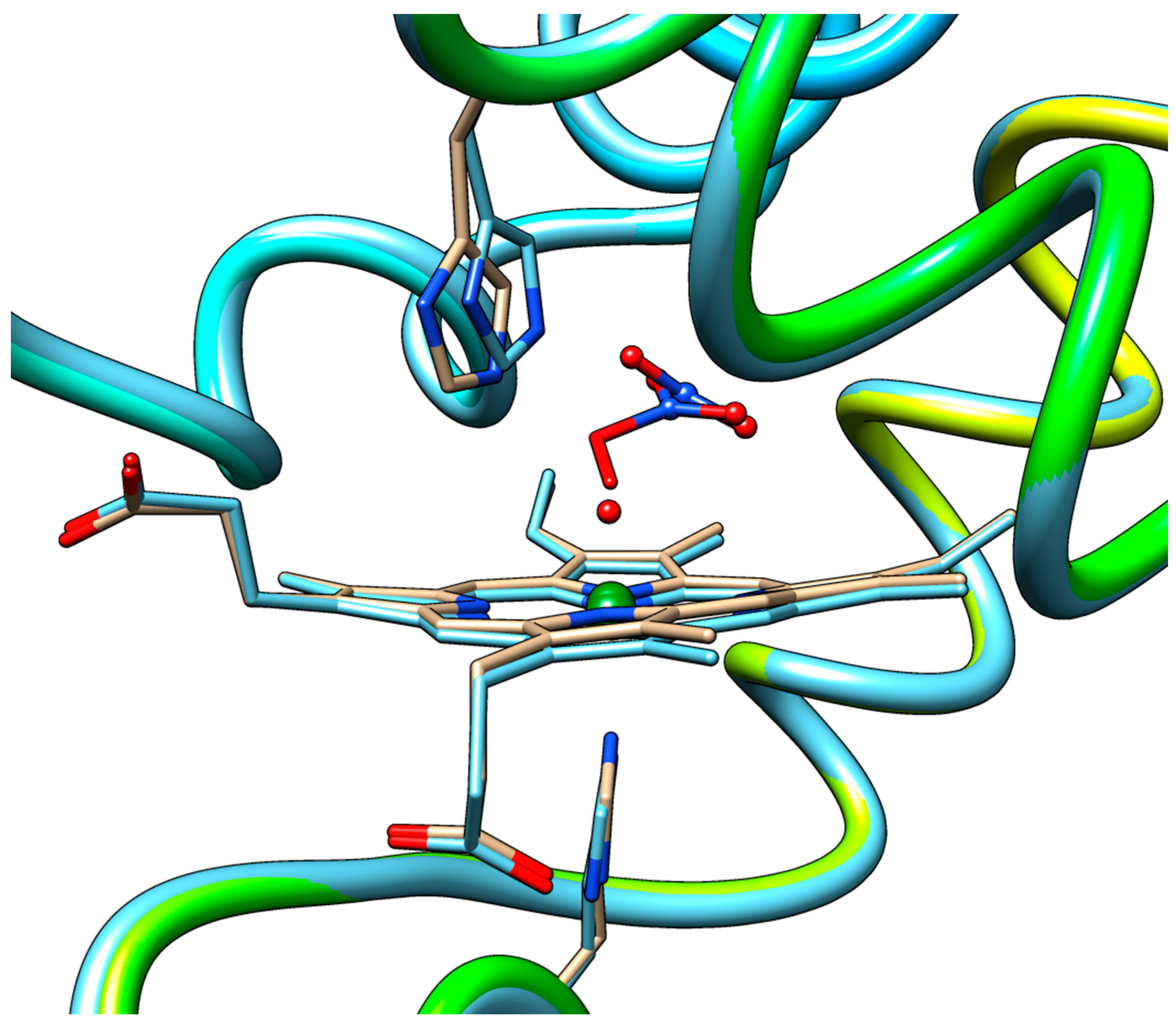
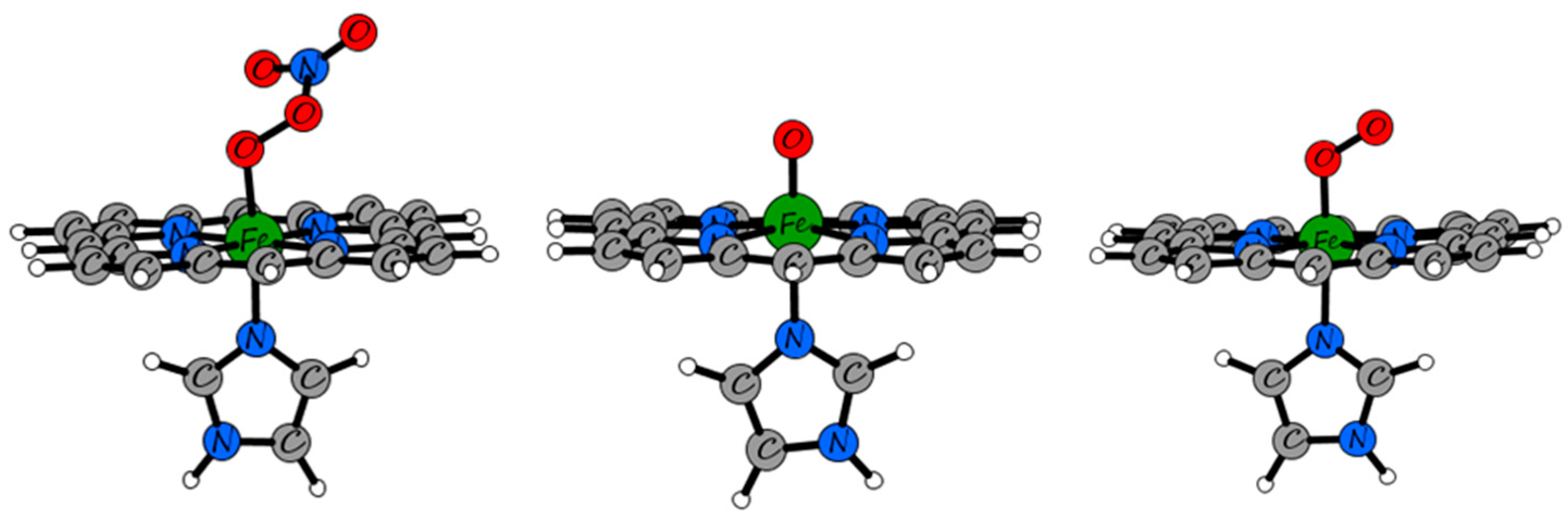
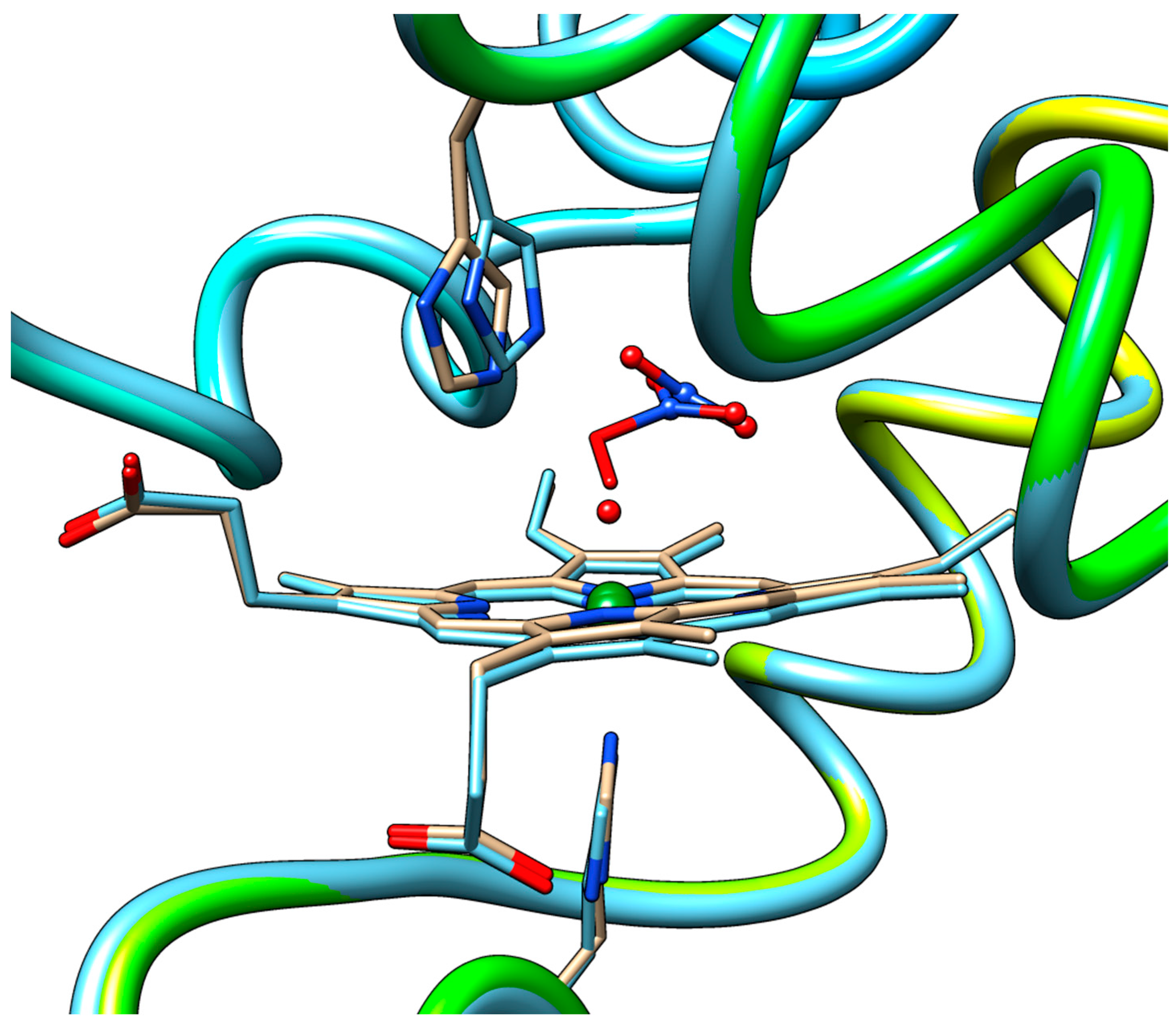
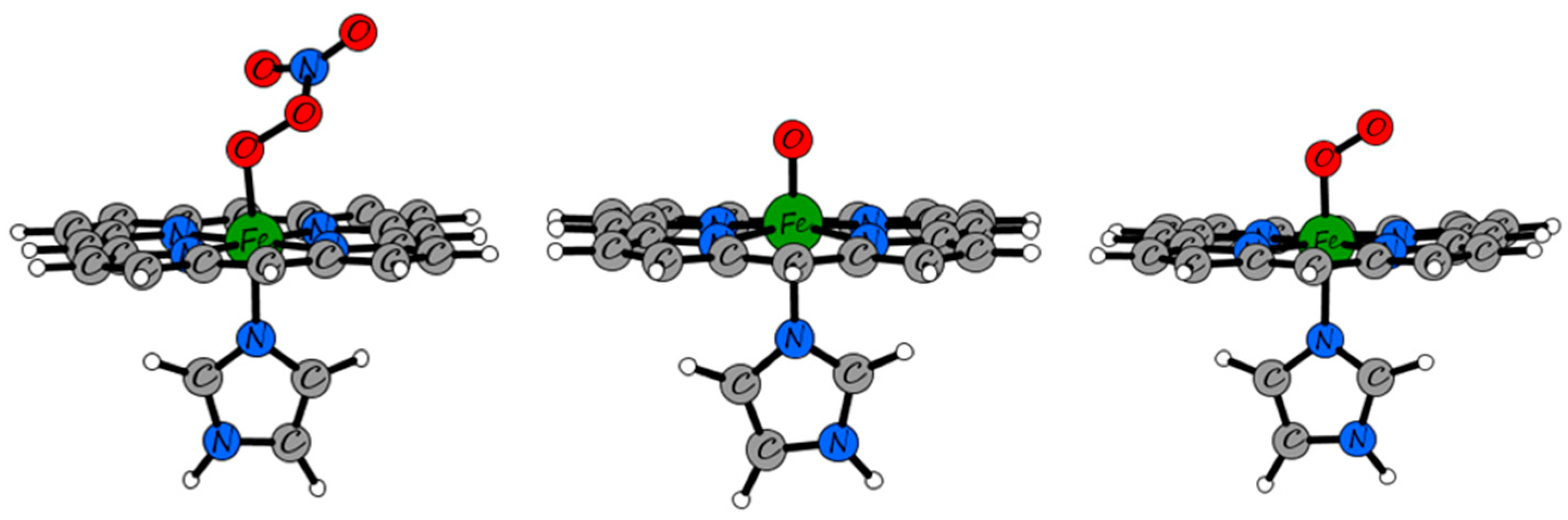
2.3. QM/MM and TD-DFT Simulations
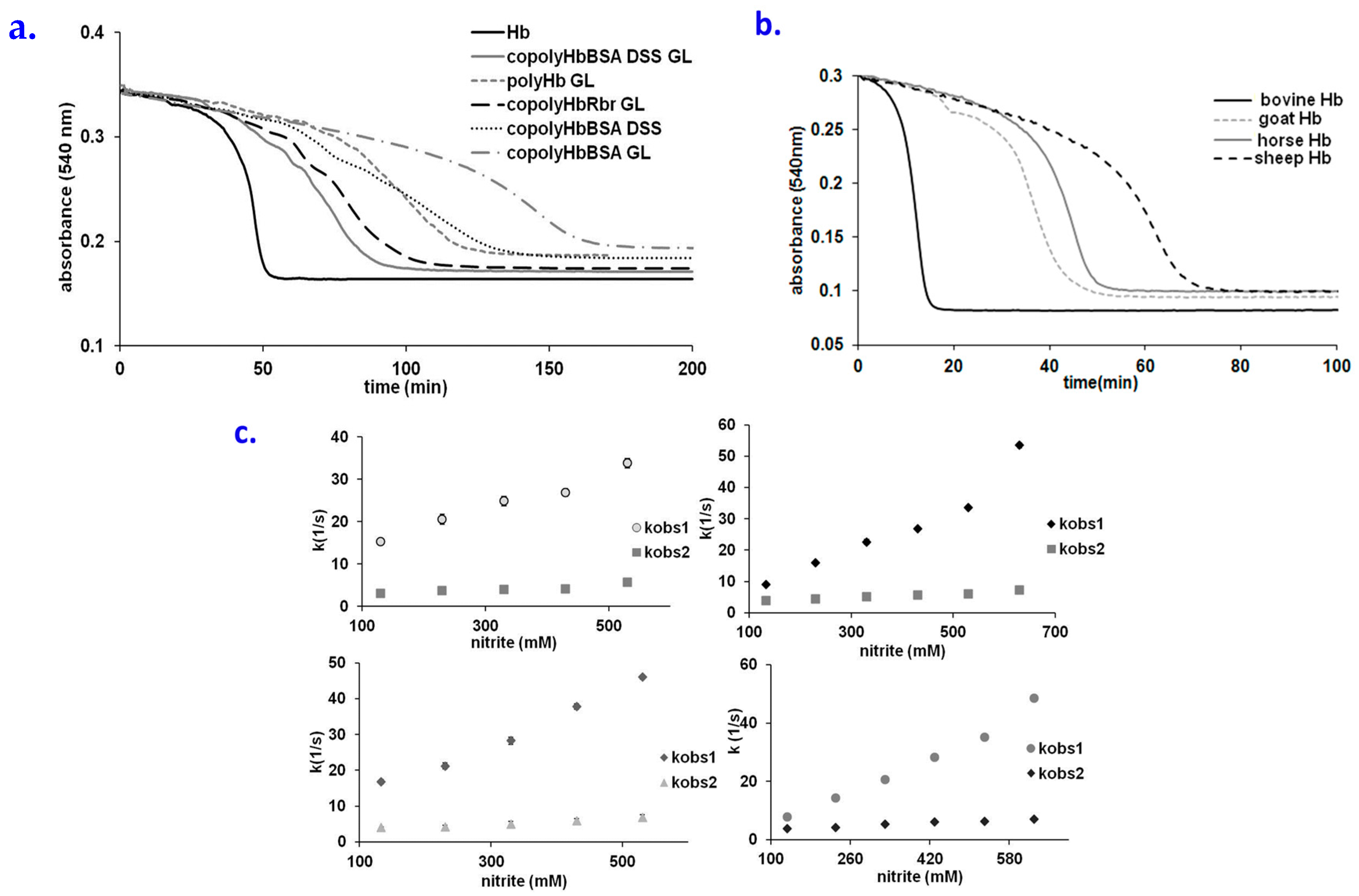
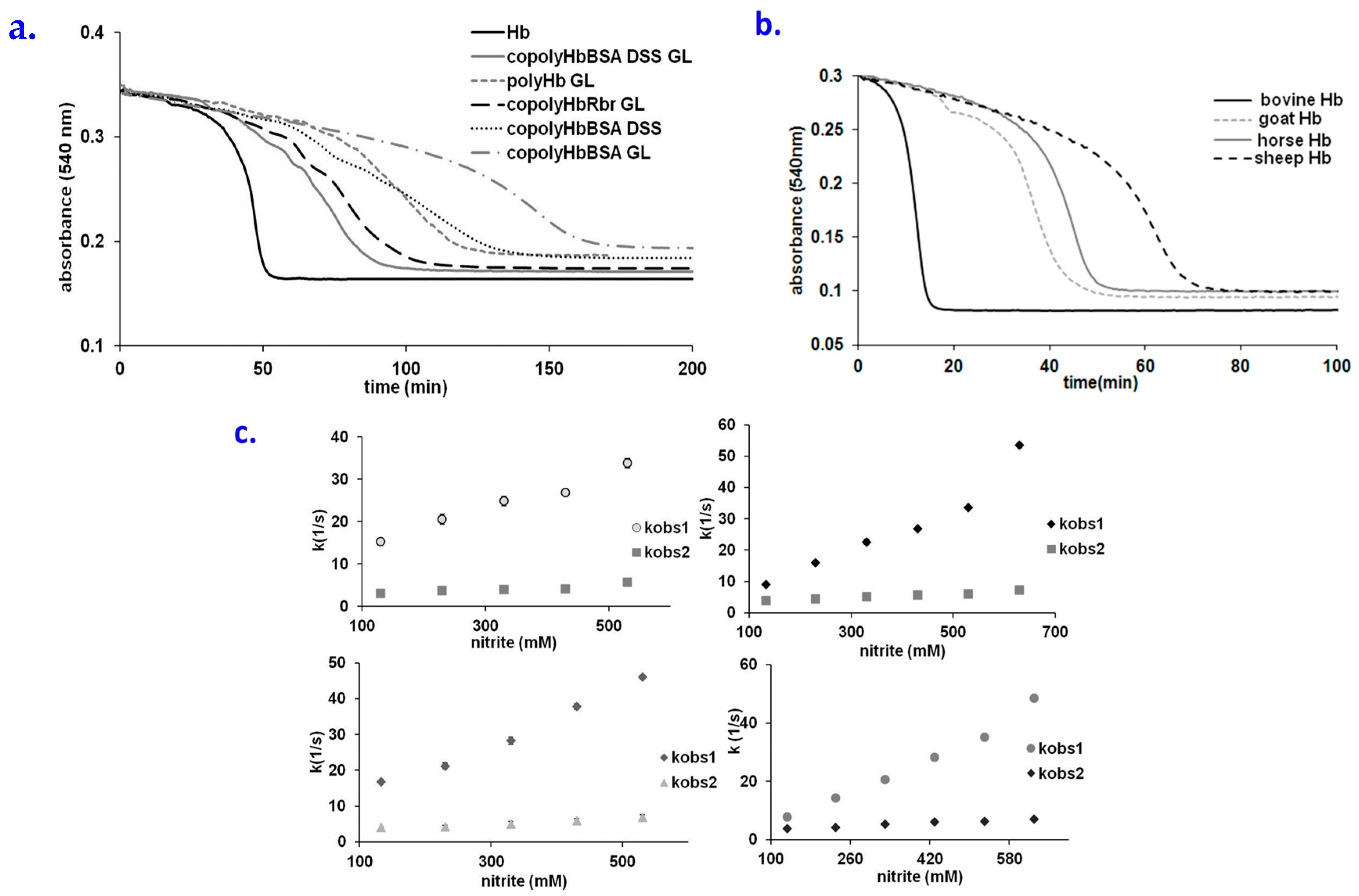
2.4. Behavior of Polymerized Hemoglobin Derivatives
3. Materials and Methods
4. Conclusions
Supplementary Materials
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Silaghi-Dumitrescu, R.; Reeder, B.J.J.; Nicholls, P.; Cooper, C.E.E.; Wilson, M.T.T. Ferryl haem protonation gates peroxidatic reactivity in globins. Biochem. J. 2007, 403, 391–395. [Google Scholar] [CrossRef] [PubMed]
- Vollaard, N.B.; Shearman, J.P.; Cooper, C.E. Exercise-induced oxidative stress: Myths, realities and physiological relevance. Sports Med. 2005, 35, 1045–1062. [Google Scholar] [CrossRef] [PubMed]
- Cooper, C.E.; Vollaard, N.B.; Choueiri, T.; Wilson, M.T. Exercise, free radicals and oxidative stress. Biochem. Soc. Trans. Trans. 2002, 30, 280–285. [Google Scholar] [CrossRef]
- Vollaard, N.B.; Reeder, B.J.; Shearman, J.P.; Menu, P.; Wilson, M.T.; Cooper, C.E. A new sensitive assay reveals that hemoglobin is oxidatively modified in vivo. Free Radic. Biol. Med. 2005, 39, 1216–1228. [Google Scholar] [CrossRef] [PubMed]
- Reeder, B.J.; Svistunenko, D.A.; Cooper, C.E.; Wilson, M.T. The radical and redox chemistry of myoglobin and hemoglobin: From in vitro studies to human pathology. Antioxid. Redox Sign. 2004, 6, 954–966. [Google Scholar] [CrossRef] [PubMed]
- Scurtu, F.; Zolog, O.; Iacob, B.; Silaghi-Dumitrescu, R. Hemoglobin-albumin cross-linking with disuccinimidyl suberate (DSS) and/or glutaraldehyde for blood substitutes. Artif. Cells Nanomed. Biotechnol. 2014, 42, 13–17. [Google Scholar] [CrossRef] [PubMed]
- Iacob, B.; Deac, F.; Cioloboc, D.; Damian, G.; Silaghi-Dumitrescu, R. Hemoglobin-albumin Crosslinked Copolymers: Reduced Prooxidant Reactivity. Artif. Cells Blood Substit. Biotechnol. 2011, 39, 293–297. [Google Scholar] [CrossRef] [PubMed]
- Alayash, A.I. Oxygen therapeutics: Can we tame haemoglobin? Nat. Rev. Drug Discov. 2004, 3, 152–159. [Google Scholar] [CrossRef] [PubMed]
- Hathazi, D.; Mahuţ, S.D.; Scurtu, F.V.; Bischin, C.; Stanciu, C.; Attia, A.A.; Damian, G.; Silaghi-Dumitrescu, R. Involvement of ferryl in the reaction between nitrite and the oxy forms of globins. J. Biol. Inorg. Chem. 2014, 19, 1233–1239. [Google Scholar] [CrossRef] [PubMed]
- Huang, K.T.; Keszler, A.; Patel, N.; Patel, R.P.; Gladwin, M.T.; Kim-Shapiro, D.B.; Hogg, N. The reaction between nitrite and deoxyhemoglobin. Reassessment of reaction kinetics and stoichiometry. J. Biol. Chem. 2005, 280, 31126–31131. [Google Scholar] [CrossRef] [PubMed]
- Keszler, A.; Piknova, B.; Schechter, A.N.; Hogg, N. The reaction between nitrite and oxyhemoglobin: A mechanistic study. J. Biol. Chem. 2008, 283, 9615–9622. [Google Scholar] [CrossRef] [PubMed]
- Kosaka, H.; Tyuma, I.; Imaizumi, K. Mechanism of autocatalytic oxidation of oxyhemoglobin by nitrite. Biomed. Biochim. Acta 1983, 42, S144–S148. [Google Scholar] [CrossRef] [PubMed]
- Imaizumi, K.; Tyuma, I.; Imai, K.; Kosaka, H.; Ueda, Y. In vivo studies on methemoglobin formation by sodium nitrite. Int. Arch. Occup. Environ. Health 1980, 45, 97–104. [Google Scholar] [CrossRef] [PubMed]
- Kosaka, H.; Imaizumi, K.; Tyuma, I. Mechanism of autocatalytic oxidation of oxyhemoglobin by nitrite. An intermediate detected by electron spin resonance. Biochim. Biophys. Acta 1982, 702, 237–241. [Google Scholar] [CrossRef]
- Kosaka, H.; Imaizumi, K.; Imai, K.; Tyuma, I. Stoichiometry of the reaction of oxyhemoglobin with nitrite. Biochim. Biophys. Acta 1979, 581, 184–188. [Google Scholar] [CrossRef]
- Kosaka, H.; Tyuma, I. Mechanism of autocatalytic oxidation of oxyhemoglobin by nitrite. Environ. Health Perspect. 1987, 73, 147–151. [Google Scholar] [CrossRef] [PubMed]
- Spagnuolo, C.; Rinelli, P.; Coletta, M.; Chiancone, E.; Ascoli, F. Oxidation reaction of human oxyhemoglobin with nitrite: A reexamination. Biochim. Biophys. Acta 1987, 911, 59–65. [Google Scholar] [CrossRef]
- Tomoda, A.; Tsuji, A.; Yoneyama, Y. Mechanism of hemoglobin oxidation by ferricytochrome C and nitrite. Acta Biol. Med. Ger. 1981, 40, 943–954. [Google Scholar] [PubMed]
- Lissi, E. Autocatalytic oxidation of hemoglobin by nitrite: A possible mechanism. Free Radic. Biol. Med. 1998, 24, 1535–1536. [Google Scholar] [CrossRef]
- Doyle, M.P.; Herman, J.G.; Dykstra, R.L. Autocatalytic oxidation of hemoglobin induced by nitrite: Activation and chemical inhibition. Free Radic. Biol. Med. 1985, 1, 145–153. [Google Scholar] [CrossRef]
- Dejam, A.; Hunter, C.J.; Schechter, A.N.; Gladwin, M.T. Emerging role of nitrite in human biology. Blood Cells Mol. Dis. 2004, 32, 423–429. [Google Scholar] [CrossRef] [PubMed]
- Kim-Shapiro, D.B.; Schechter, A.N.; Gladwin, M.T. Unraveling the reactions of nitric oxide, nitrite, and hemoglobin in physiology and therapeutics. Arterioscler. Thromb. Vasc. Biol. 2006, 26, 697–705. [Google Scholar] [CrossRef] [PubMed]
- Gladwin, M.T.; Raat, N.J.; Shiva, S.; Dezfulian, C.; Hogg, N.; Kim-Shapiro, D.B.; Patel, R.P. Nitrite as a vascular endocrine nitric oxide reservoir that contributes to hypoxic signaling, cytoprotection, and vasodilation. Am. J. Physiol. Heart Circ. Physiol. 2006, 291, H2026–H2035. [Google Scholar] [CrossRef] [PubMed]
- Gladwin, M.T.; Grubina, R.; Doyle, M.P. The new chemical biology of nitrite reactions with hemoglobin: R-state catalysis, oxidative denitrosylation, and nitrite reductase/anhydrase. Acc. Chem. Res. 2009, 42, 157–167. [Google Scholar] [CrossRef] [PubMed]
- Crawford, J.H.; Isbell, T.S.; Huang, Z.; Shiva, S.; Chacko, B.K.; Schechter, A.N.; Darley-Usmar, V.M.; Kerby, J.D.; Lang, J.D.; Kraus, D.; et al. Hypoxia, red blood cells, and nitrite regulate NO-dependent hypoxic vasodilation. Blood 2006, 107, 566–574. [Google Scholar] [CrossRef] [PubMed]
- Goetz, B.I.; Wang, P.; Shields, H.W.; Basu, S.; Grubina, R.; Huang, J.; Conradie, J.; Huang, Z.; Jeffers, A.; Jiang, A.; et al. Reply to ‘Nitrite–methemoglobin inadequate for hypoxic vasodilation’. Nat. Chem. Biol. 2009, 5, 367. [Google Scholar] [CrossRef]
- Schwab, D.E.; Stamler, J.S.; Singel, D.J. Nitrite–methemoglobin inadequate for hypoxic vasodilation. Nat. Chem. Biol. 2009, 5, 366. [Google Scholar] [CrossRef] [PubMed]
- Dezfulian, C.; Raat, N.; Shiva, S.; Gladwin, M.T. Role of the anion nitrite in ischemia-reperfusion cytoprotection and therapeutics. Cardiovasc. Res. 2007, 75, 327–338. [Google Scholar] [CrossRef] [PubMed]
- Chen, K.; Piknova, B.; Pittman, R.N.; Schechter, A.N.; Popel, A.S. Nitric oxide from nitrite reduction by hemoglobin in the plasma and erythrocytes. Nitric Oxide 2008, 18, 47–60. [Google Scholar] [CrossRef] [PubMed]
- Weiss, S.J. The role of superoxide in the destruction of erythrocyte targets by human neutrophils. J. Biol. Chem. 1980, 255, 9912–9917. [Google Scholar] [PubMed]
- Grubina, R.; Huang, Z.; Shiva, S.; Joshi, M.S.; Azarov, I.; Basu, S.; Ringwood, L.A.; Jiang, A.; Hogg, N.; Kim-Shapiro, D.B.; et al. Concerted nitric oxide formation and release from the simultaneous reactions of nitrite with deoxy- and oxyhemoglobin. J. Biol. Chem. 2007, 282, 12916–12927. [Google Scholar] [CrossRef] [PubMed]
- Grubina, R.; Basu, S.; Tiso, M.; Kim-Shapiro, D.B.; Gladwin, M.T. Nitrite reductase activity of hemoglobin S (sickle) provides insight into contributions of heme redox potential versus ligand affinity. J. Biol. Chem. 2008, 283, 3628–3638. [Google Scholar] [CrossRef] [PubMed]
- Hopmann, K.H.; Cardey, B.; Gladwin, M.T.; Kim-Shapiro, D.B.; Ghosh, A. Hemoglobin as a nitrite anhydrase: Modeling methemoglobin-mediated N2O3 formation. Chemistry 2011, 17, 6348–6358. [Google Scholar] [CrossRef] [PubMed]
- Minneci, P.C.; Deans, K.J.; Shiva, S.; Zhi, H.; Banks, S.M.; Kern, S.; Natanson, C.; Solomon, S.B.; Gladwin, M.T. Nitrite reductase activity of hemoglobin as a systemic nitric oxide generator mechanism to detoxify plasma hemoglobin produced during hemolysis. Am. J. Physiol. Heart Circ. Physiol. 2008, 295, H743–H754. [Google Scholar] [CrossRef] [PubMed]
- Cooper, C.E.E.; Silaghi-Dumitrescu, R.; Rukengwa, M.; Alayash, A.I.I.; Buehler, P.W.W. Peroxidase activity of hemoglobin towards ascorbate and urate: A synergistic protective strategy against toxicity of Hemoglobin-Based Oxygen Carriers (HBOC). Biochim. Biophys. Acta 2008, 1784, 1415–1420. [Google Scholar] [CrossRef] [PubMed]
- Deac, F.V.; Bolfa, A.M.; Magdas, C.; Sevastre, B.; Turc, S.; Silaghi-Dumitrescu, R. Hemoglobin-based blood substitutes: Which hemoglobin to use? Rom. J. Biochem. 2010, 47, 135–141. [Google Scholar]
- Deac, F.V.; Todea, A.; Bolfa, A.M.; Podea, P.; Petrar, P.; Silaghi-Dumitrescu, R. Ascorbate Binding to Globins. Rom. J. Biochem. 2009, 46, 115–121. [Google Scholar]
- Goldstein, S.; Merenyi, G.; Samuni, A. Kinetics and mechanism of •NO2 reacting with various oxidation states of myoglobin. J. Am. Chem. Soc. 2004, 126, 15694–15701. [Google Scholar] [CrossRef] [PubMed]
- Ratanasopa, K.; Strader, M.B.; Alayash, A.I.; Bulow, L. Dissection of the radical reactions linked to fetal hemoglobin reveals enhanced pseudoperoxidase activity. Front. Physiol. 2015, 6, 39. [Google Scholar] [CrossRef] [PubMed]
- Wade, R.S.; Castro, C.E. Reactions of oxymyoglobin with NO, NO2, and NO2− under argon and in air. Chem. Res. Toxicol. 1996, 9, 1382–1390. [Google Scholar] [CrossRef] [PubMed]
- Bischin, C.; Deac, F.; Silaghi-Dumitrescu, R.; Worrall, J.A.R.A.R.; Rajagopal, B.S.S.; Damian, G.; Cooper, C.E.E. Ascorbate peroxidase activity of cytochrome c. Free Radic. Res. 2011, 45, 439–444. [Google Scholar] [CrossRef] [PubMed]
- Antonini, E.; Brunori, M. Hemoglobin and Myoglobin in Their Reaction with Ligands; Elsevier: Amsterdam, The Netherlands, 1971. [Google Scholar]
- Silaghi-Dumitrescu, R.; Svistunenko, D.A.A.; Cioloboc, D.; Bischin, C.; Scurtu, F.; Cooper, C.E.E. Nitrite binding to globins: Linkage isomerism, EPR silence and reductive chemistry. Nitric Oxide Biol. Chem. 2014, 42, 32–39. [Google Scholar] [CrossRef] [PubMed]
- D’Agnillo, F.; Chang, T.M. Polyhemoglobin-superoxide dismutase-catalase as a blood substitute with antioxidant properties. Nat. Biotechnol. 1998, 16, 667–671. [Google Scholar] [CrossRef] [PubMed]
- Alayash, A.I. Blood substitutes: Why haven’t we been more successful? Trends Biotechnol. 2014, 32, 177–185. [Google Scholar] [CrossRef] [PubMed]
- Toma, V.A.; Farcas, A.D.; Roman, I.; Sevastre, B.; Hathazi, D.; Scurtu, F.; Damian, G.; Silaghi-Dumitrescu, R. Comparative in vivo Effects of Hemoglobin-Based Oxygen Carriers (HBOC) with Varying Prooxidant and Physiological Reactivity. PLoS ONE 2016, 11, e0153909. [Google Scholar] [CrossRef] [PubMed]
- Arkosi, M.; Scurtu, F.; Vulpoi, A.; Silaghi-Dumitrescu, R.; Kurtz, D., Jr. Copolymerization of Recombinant P. gouldii Hemerythrin with Human Serum Albumin for Use in Blood Substitutes. Artif. Cells Blood Substitutes Biotechnol. 2017, 45, 218–223. [Google Scholar] [CrossRef] [PubMed]
- Toma, V.A.A.; Farcas, A.D.D.; Roman, I.; Sevastre, B.; Hathazi, D.; Scurtu, F.; Damian, G.; Silaghi-Dumitrescu, R. In vivo evaluation of hemerythrin-based oxygen carriers: Similarities with hemoglobin-based counterparts. Int. J. Biol. Macromol. 2017, 107, 1422–1427. [Google Scholar] [CrossRef] [PubMed]
- Scurtu, V.F.; Mot, A.C.; Silaghi-Dumitrescu, R. Protein-based blood substitutes: Recent attempts at controlling pro-oxidant reactivity with and beyond hemoglobin. Pharmaceuticals 2013, 6, 867–880. [Google Scholar] [CrossRef] [PubMed]
- Benedec, D.; Popica, I.-E.; Oniga, I.; Hanganu, D.; Duma, M.; Silaghi-Dumitrescu, R.; Bischin, C.; Vlase, L. Comparative HPLC-MS analysis of phenolics from achillea distans and achillea millefolium and their bioactivity. Stud. Univ. Babes-Bolyai Chem. 2015, 60, 257–266. [Google Scholar]
- Mot, A.C.C.; Bischin, C.; Muresan, B.; Parvu, M.; Damian, G.; Vlase, L.; Silaghi-Dumitrescu, R. Antioxidant activity evaluation by physiologically relevant assays based on haemoglobin peroxidase activity and cytochrome c-induced oxidation of liposomes. Nat. Prod. Res. 2016, 30, 1315–1319. [Google Scholar] [CrossRef] [PubMed]
- Găină, L.; Torje, I.; Gal, E.; Lupan, A.; Bischin, C.; Silaghi-Dumitrescu, R.; Damian, G.; Lönnecke, P.; Cristea, C.; Silaghi-Dumitrescu, L. Microwave assisted synthesis, photophysical and redox properties of (phenothiazinyl)vinyl-pyridinium dyes. Dyes Pigment. 2014, 102, 315–325. [Google Scholar] [CrossRef]
- Benedec, D.; Hanganu, D.; Oniga, I.; Tiperciuc, B.; Olah, N.K.; Raita, O.; Bischin, C.; Silaghi-Dumitrescu, R.; Vlase, L. Assessment of rosmarinic acid content in six Lamiaceae species extracts and their antioxidant and antimicrobial potential. Pak. J. Pharm. Sci. 2015, 28, 2297–2303. [Google Scholar] [PubMed]
- Benedec, D.; Hanganu, D.; Oniga, I.; Filip, L.; Bischin, C.; Silaghi-Dumitrescu, R.; Tiperciuc, B.; Vlase, L. Achillea schurii flowers: Chemical, antioxidant, and antimicrobial investigations. Molecules 2016, 21, 1050. [Google Scholar] [CrossRef] [PubMed]
- Mureşan, B.; Cimpoiu, C.; Hosu, A.; Bischin, C.; Gal, E.; Damian, G.; Fischer-Fodor, E.; Silaghi-Dumitrescu, R. Antioxidant content in Romanian traditional distilled alcoholic beverages. Stud. Univ. Babes-Bolyai Chem. 2015, 60, 355–370. [Google Scholar]
- Mocan, A.; Vlase, L.; Arsene, A.L.; Vodnar, D.; Bischin, C.; Silaghi-Dumitrescu, R.; Crişan, G. HPLC/MS analysis of caffeic and chlorogenic acids from three romanian veronica species and their antioxidant and antimicrobial properties. Farmacia 2015, 63, 890–896. [Google Scholar]
- Bischin, C.; Tusan, C.; Bartok, A.; Septelean, R.; Damian, G.; Silaghi-Dumitrescu, R. Evaluation of the biochemical effects of silyl-phosphaalkenes on oxidative and nitrosative stress pathways involving metallocenters. Phosphorus, Sulfur Silicon Relat. Elem. 2015, 190, 292–299. [Google Scholar] [CrossRef]
- Bischin, C.; Contra, G.; Tusan, C.; Miclea, P.; Taciuc, V.; Parvu, M.; Silaghi-Dumitrescu, R. Free-radical reactions: The fine line between the anti- and pro-oxidant reactivities. Oxid. Commun. 2008, in press. [Google Scholar]
- Dezsi, S.; Badarau, A.S.; Bischin, C.; Vodnar, D.C.; Silaghi-Dumitrescu, R.; Gheldiu, A.M.; Mocan, A.; Vlase, L. Antimicrobial and antioxidant activities and phenolic profile of Eucalyptus globulus Labill. and Corymbia ficifolia (F. Muell.) K.D. Hill & L.A.S. Johnson leaves. Molecules 2015, 20, 4720–4734. [Google Scholar] [PubMed]
- Mot, A.C.C.; Bischin, C.; Damian, G.; Silaghi-Dumitrescu, R. Antioxidant activity evaluation involving hemoglobin-related free radical reactivity. Methods Mol. Biol. 2015, 1208, 247–255. [Google Scholar] [PubMed]
- Vlase, L.; Benedec, D.; Hanganu, D.; Damian, G.; Csillag, I.; Sevastre, B.; Mot, A.C.C.; Silaghi-Dumitrescu, R.; Tilea, I. Evaluation of antioxidant and antimicrobial activities and phenolic profile for Hyssopus officinalis, Ocimum basilicum and Teucrium chamaedrys. Molecules 2014, 19, 5490–5507. [Google Scholar] [CrossRef] [PubMed]
- Deac, F.; Iacob, B.; Fischer-Fodor, E.; Damian, G.; Silaghi-Dumitrescu, R. Derivatization of haemoglobin with periodate-generated reticulation agents: Evaluation of oxidative reactivity for potential blood substitutes. J. Biochem. 2011, 149, 75–82. [Google Scholar] [CrossRef] [PubMed]
- Hathazi, D.; Mot, A.C.; Vaida, A.; Scurtu, F.; Lupan, I.; Fischer-Fodor, E.; Damian, G.; Kurtz, D.M., Jr.; Silaghi-Dumitrescu, R. Oxidative protection of hemoglobin and hemerythrin by cross-linking with a nonheme iron peroxidase: Potentially improved oxygen carriers for use in blood substitutes. Biomacromolecules 2014, 15, 1920–1927. [Google Scholar] [CrossRef] [PubMed]
- Dunne, J.; Caron, A.; Menu, P.; Alayash, A.I.I.; Buehler, P.W.W.; Wilson, M.T.T.; Silaghi-Dumitrescu, R.; Faivre, B.; Cooper, C.E.E. Ascorbate removes key precursors to oxidative damage by cell-free haemoglobin in vitro and in vivo. Biochem. J. 2006, 399, 513–524. [Google Scholar] [CrossRef] [PubMed]
- Fischer-Fodor, E.; Mot, A.; Deac, F.; Arkosi, M.; Silaghi-Dumitrescu, R. Towards hemerythrin-based blood substitutes: Comparative performance to hemoglobin on human leukocytes and umbilical vein endothelial cells. J. Biosci. 2011, 36, 215–221. [Google Scholar] [CrossRef] [PubMed]
- Zolog, O.; Mot, A.; Deac, F.; Roman, A.; Fischer-Fodor, E.; Silaghi-Dumitrescu, R. A New Polyethyleneglycol-Derivatized Hemoglobin Derivative with Decreased Oxygen Affinity and Limited Toxicity. Protein J. 2011, 30, 27–31. [Google Scholar] [CrossRef] [PubMed]
- Schrödinger Release 2014-1: Schrödinger Suite 2014-1 Protein Preparation Wizard; Epik Version 2.7; Schrödinger, LLC: New York, NY, USA, 2014.
- Small-Molecule Drug Discovery Suite 2014-1: QSite, Version 6.2; Schrödinger, LLC: New York, NY, USA, 2014.
- Unno, M.; Chen, H.; Kusama, S.; Shaik, S.; Ikeda-Saito, M. Structural characterization of the fleeting ferric peroxo species in myoglobin: Experiment and theory. J. Am. Chem. Soc. 2007, 129, 13394–13395. [Google Scholar] [CrossRef] [PubMed]
- Schrödinger Release 2014-1: Maestro, Version 9.7; Schrödinger, LLC: New York, NY, USA, 2014.
- Zhao, Y.; Truhlar, D.G. A new local density functional for main-group thermochemistry, transition metal bonding, thermochemical kinetics, and noncovalent interactions. J. Chem. Phys. 2006, 125, 194101–194118. [Google Scholar] [CrossRef] [PubMed]
Sample Availability: Samples of the hemoglobins employed in the present study are available from the authors. |

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Model | Oscillator Strength | Excitation Energy (nm) |
|---|---|---|
| Heme–peroxynitrate | 0.374 | 379 |
| 0.401 | 376 | |
| Heme-oxy | 0.200 | 379 |
| 0.153 | 368 | |
| 0.359 | 365 | |
| 0.249 | 355 | |
| Ferryl | 0.113 | 458 |
| 0.174 | 424 | |
| 0.094 | 417 |
| K1 | n1 | K2 | n2 | |
|---|---|---|---|---|
| Hb | 112 M−1 s−1 | 2 | 11.51 s−1 | 1 |
| polyHb | 3.4 M−1 s−1 | 0.5 | 6.3 M−1 s−1 | 0.4 |
| copolyHbBSA | 73 s−1 | 1.1 | 7.9 M−1 s−1 | 0.4 |
| copolyHbRbr | 68.8 s−1 | 0.7 | 9.3 M−1 s−1 | 0.5 |
| copolyHbBSA DSS | 75.8 s−1 | 1.1 | 8.3 M−1 s−1 | 0.4 |
| Type of Hemoglobin | kobs1(1/s) | Stdev1 | kobs2(1/s) | Stdev2 |
|---|---|---|---|---|
| Bovine Hb | 51.53 | 0.91 | 5.75 | 0.21 |
| Goat Hb | 58.30 | 0.85 | 10.87 | 0.35 |
| Cabaline Hb | 43.93 | 1.03 | 11.36 | 0.50 |
| Ovine Hb | 27.02 | 0.65 | 8.48 | 0.23 |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hathazi, D.; Scurtu, F.; Bischin, C.; Mot, A.; Attia, A.A.A.; Kongsted, J.; Silaghi-Dumitrescu, R. The Reaction of Oxy Hemoglobin with Nitrite: Mechanism, Antioxidant-Modulated Effect, and Implications for Blood Substitute Evaluation. Molecules 2018, 23, 350. https://doi.org/10.3390/molecules23020350
Hathazi D, Scurtu F, Bischin C, Mot A, Attia AAA, Kongsted J, Silaghi-Dumitrescu R. The Reaction of Oxy Hemoglobin with Nitrite: Mechanism, Antioxidant-Modulated Effect, and Implications for Blood Substitute Evaluation. Molecules. 2018; 23(2):350. https://doi.org/10.3390/molecules23020350
Chicago/Turabian StyleHathazi, Denisa, Florina Scurtu, Cristina Bischin, Augustin Mot, Amr A. A. Attia, Jacob Kongsted, and Radu Silaghi-Dumitrescu. 2018. "The Reaction of Oxy Hemoglobin with Nitrite: Mechanism, Antioxidant-Modulated Effect, and Implications for Blood Substitute Evaluation" Molecules 23, no. 2: 350. https://doi.org/10.3390/molecules23020350