Genome-Wide Identification and Comparative Analysis of the 3-Hydroxy-3-methylglutaryl Coenzyme A Reductase (HMGR) Gene Family in Gossypium

Abstract
:1. Introduction
2. Results
2.1. Genome-Wide Identification of HMGR Genes in Gossypium
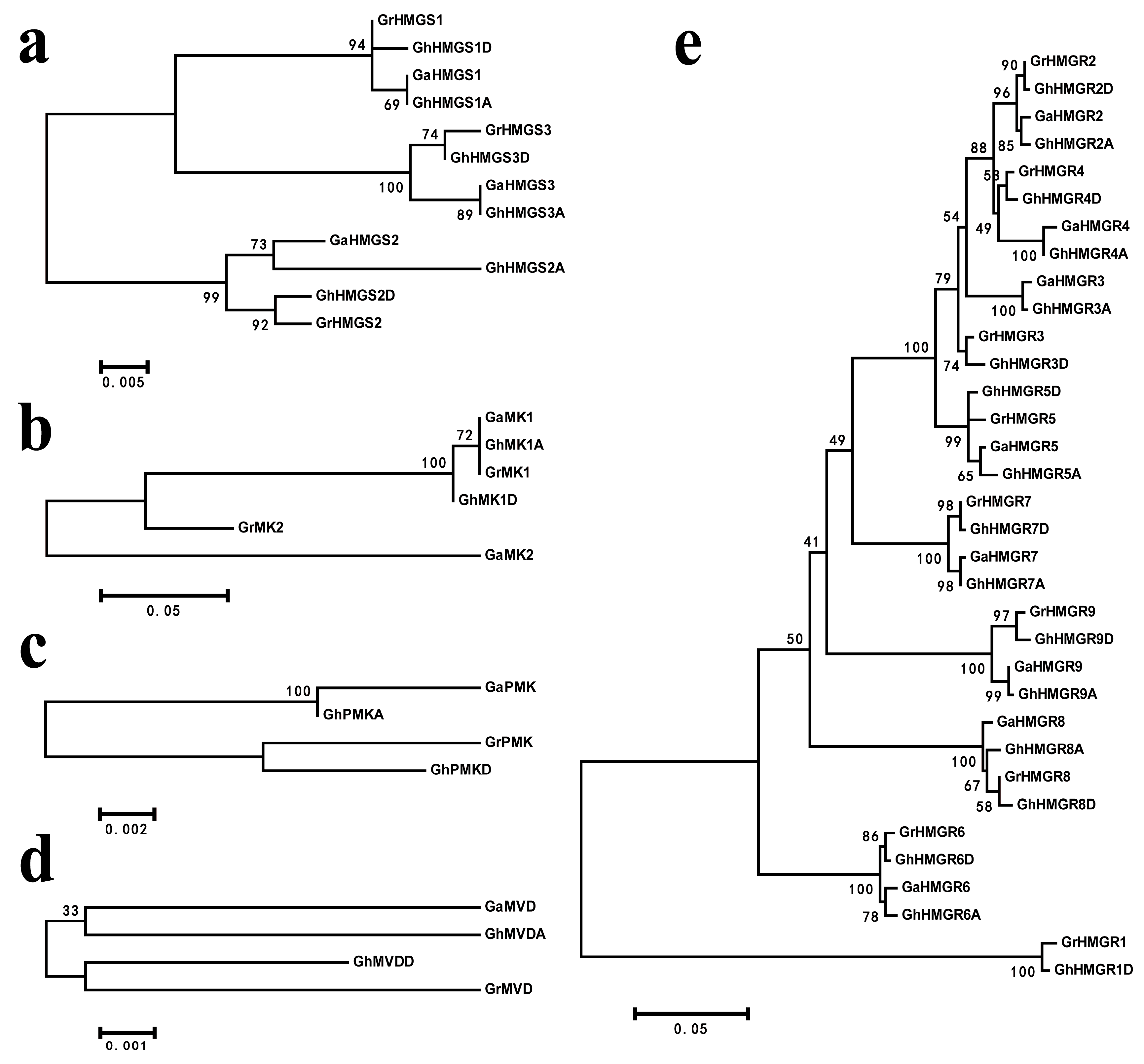
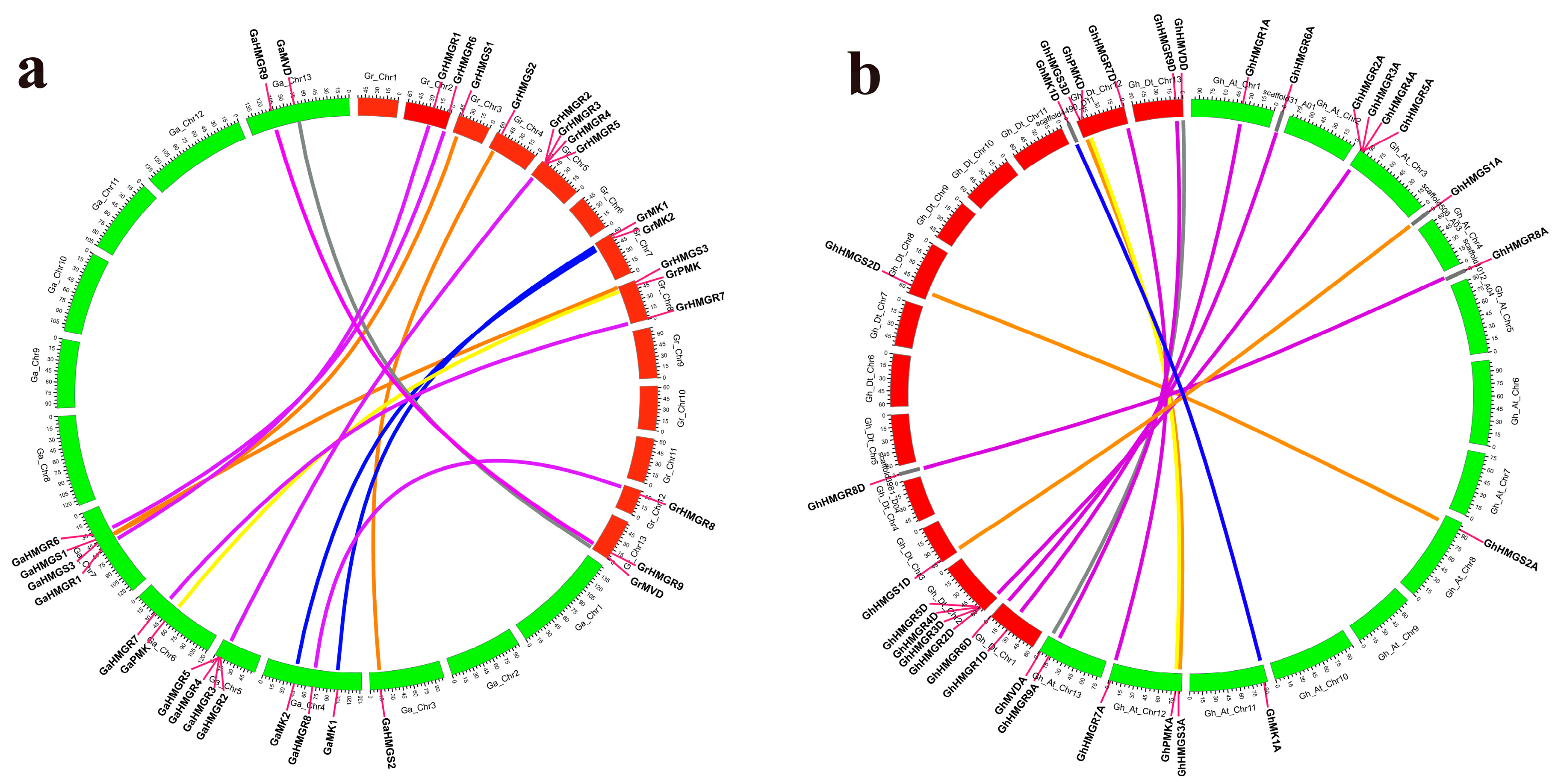
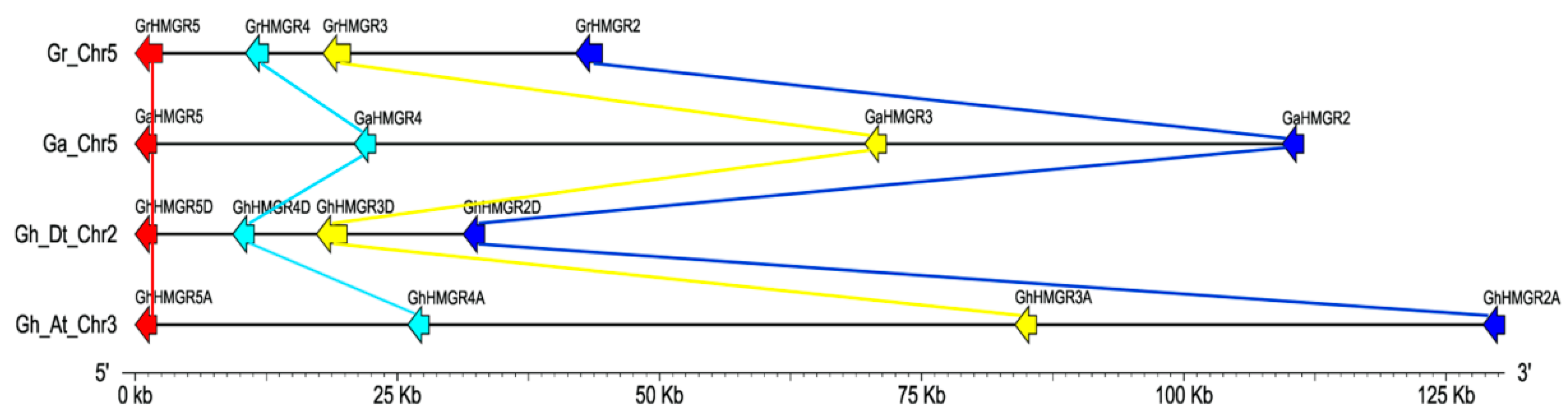
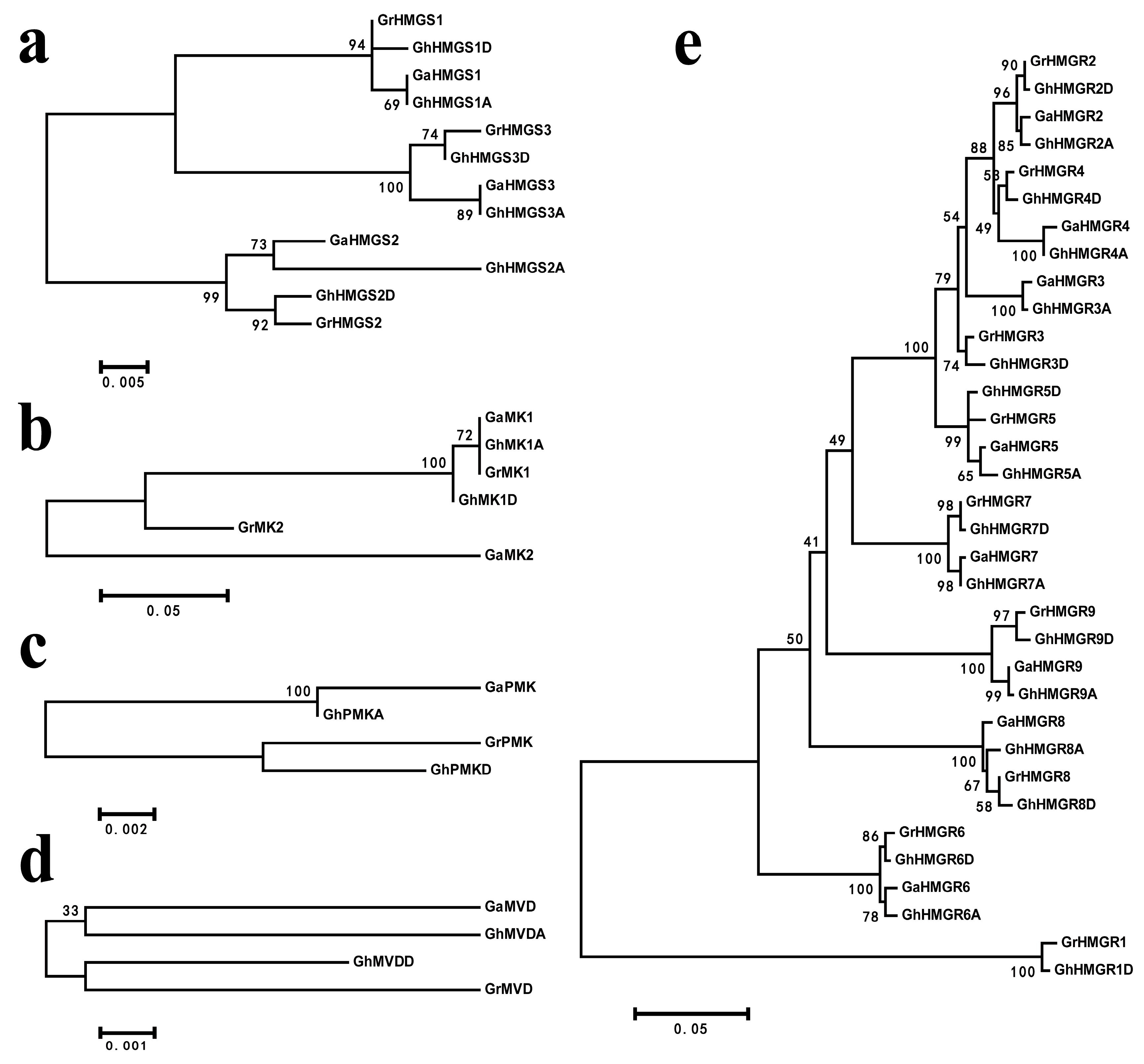
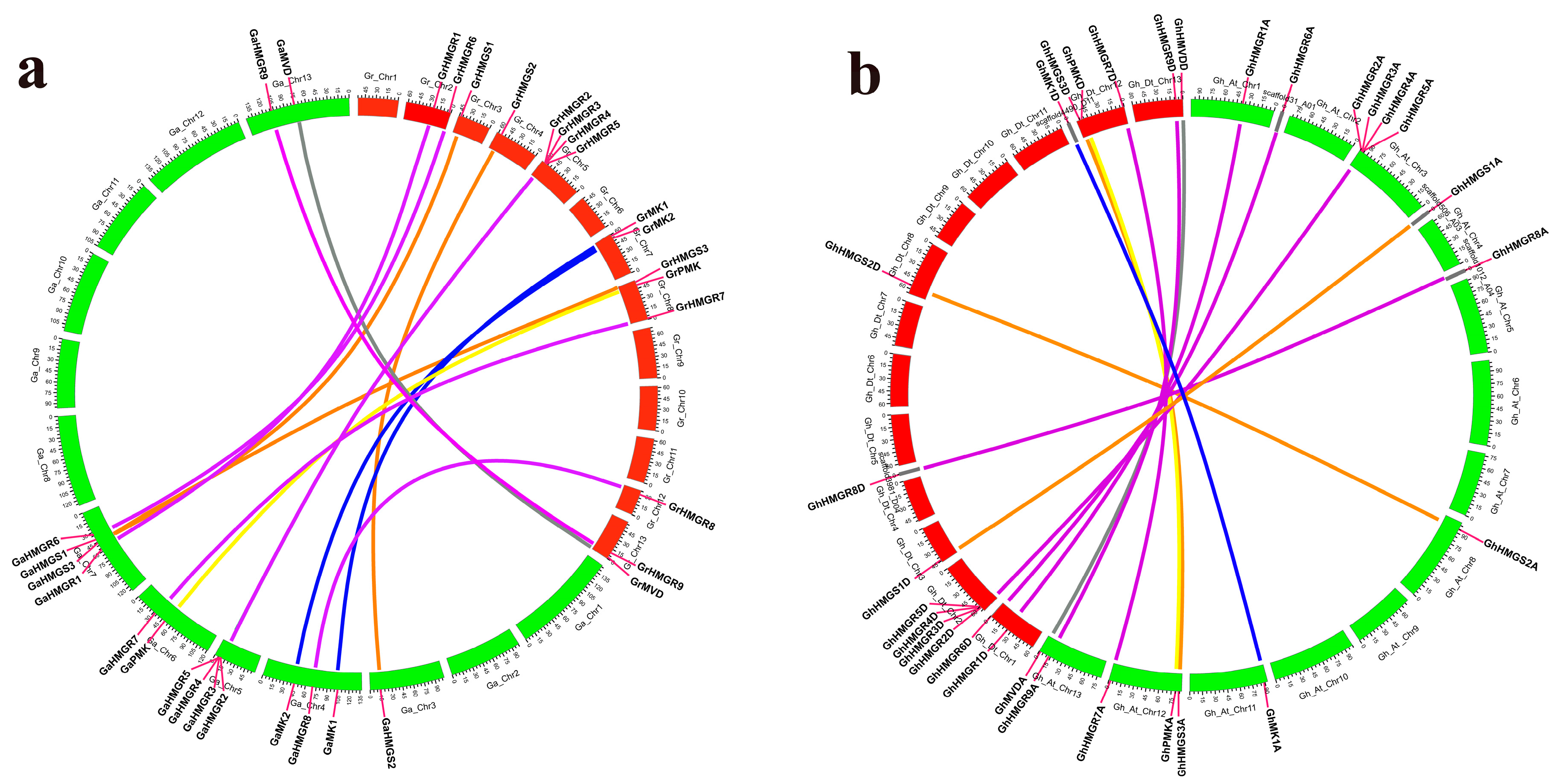
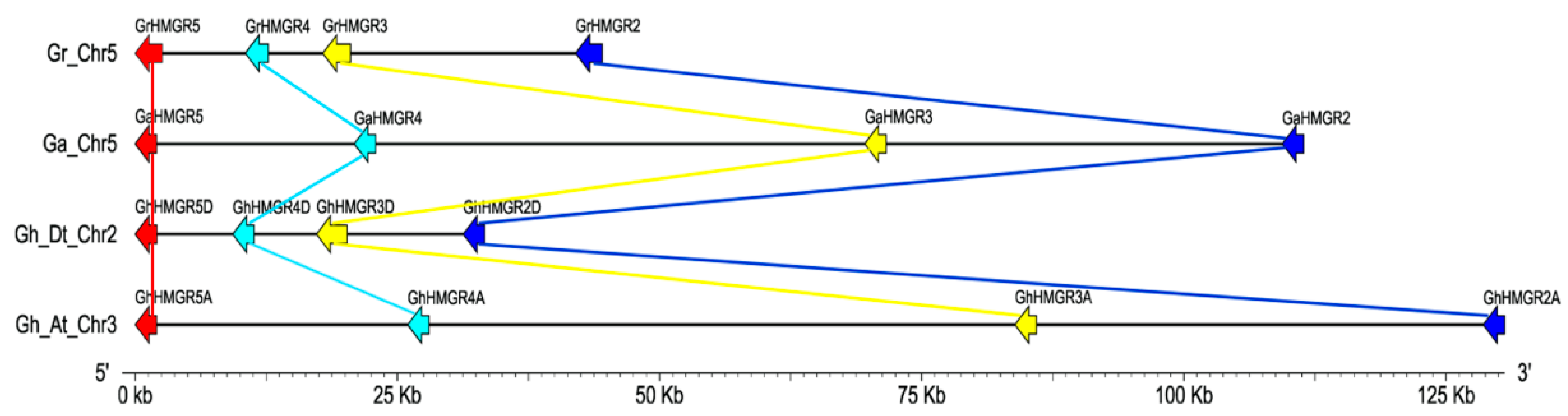
2.2. Chromosomal Distribution and Phylogenetic Analysis of Gossypium HMGR Genes
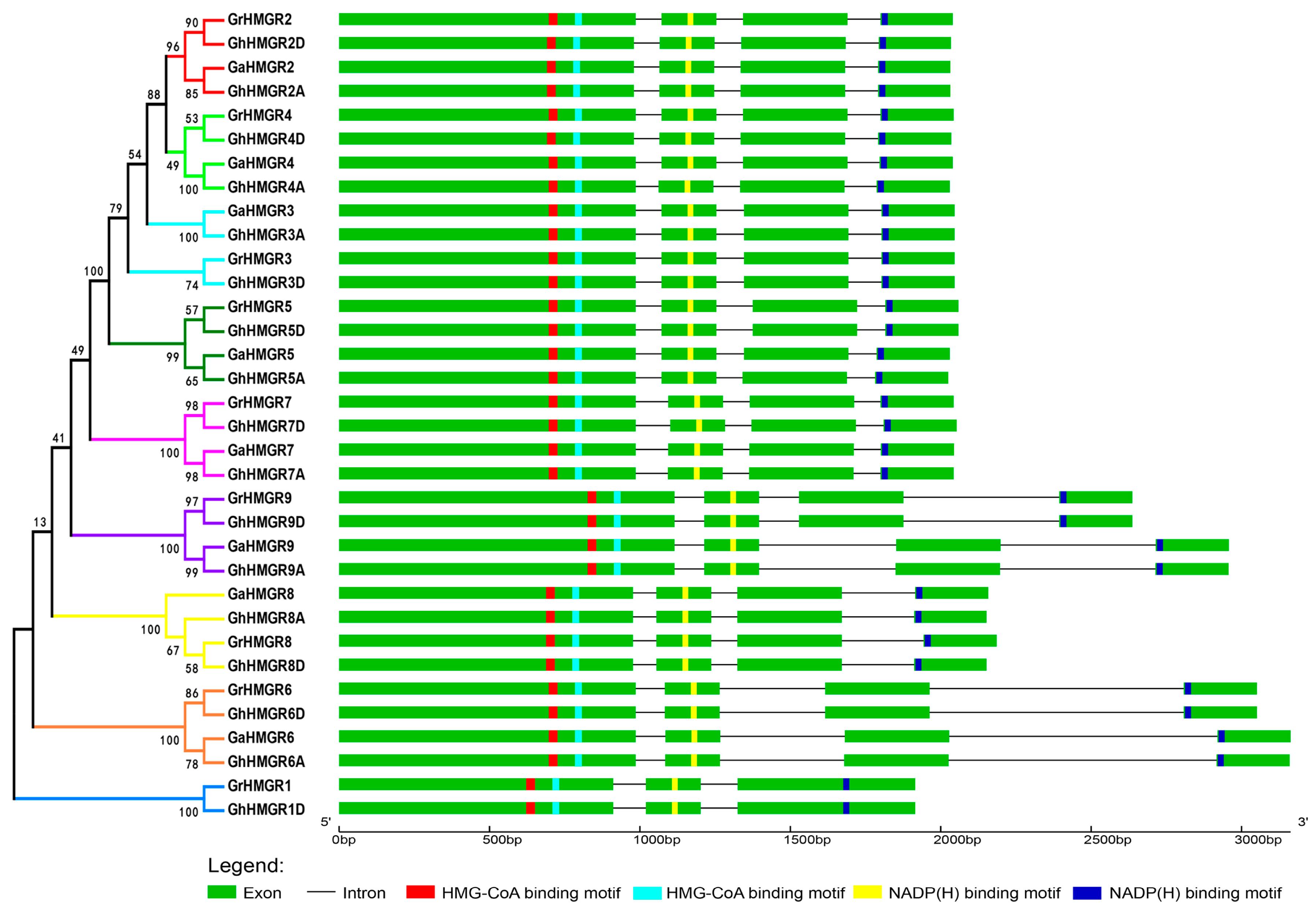
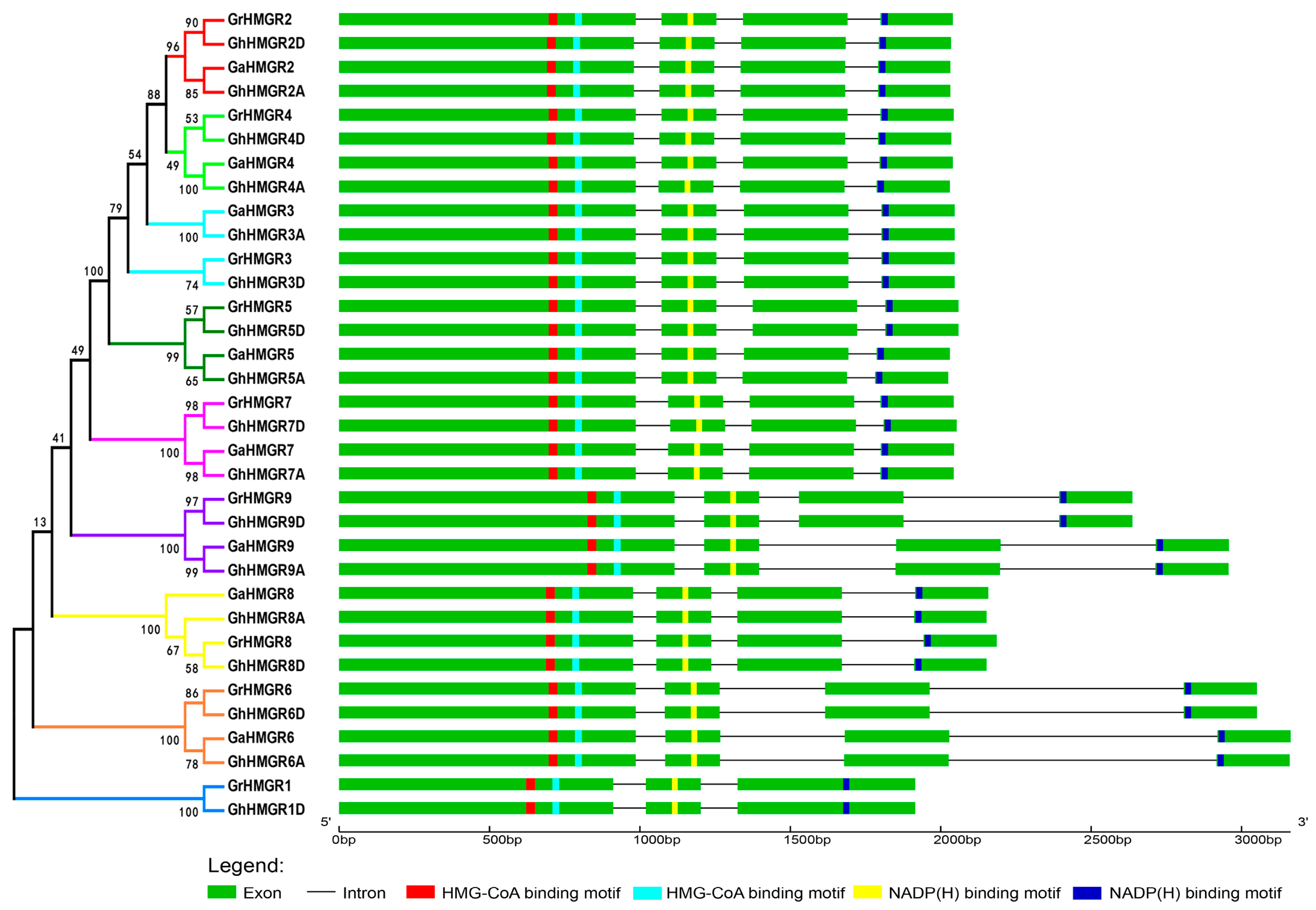
2.3. Gene Structure and Conserved Protein Motifs of Gossypium HMGR Genes
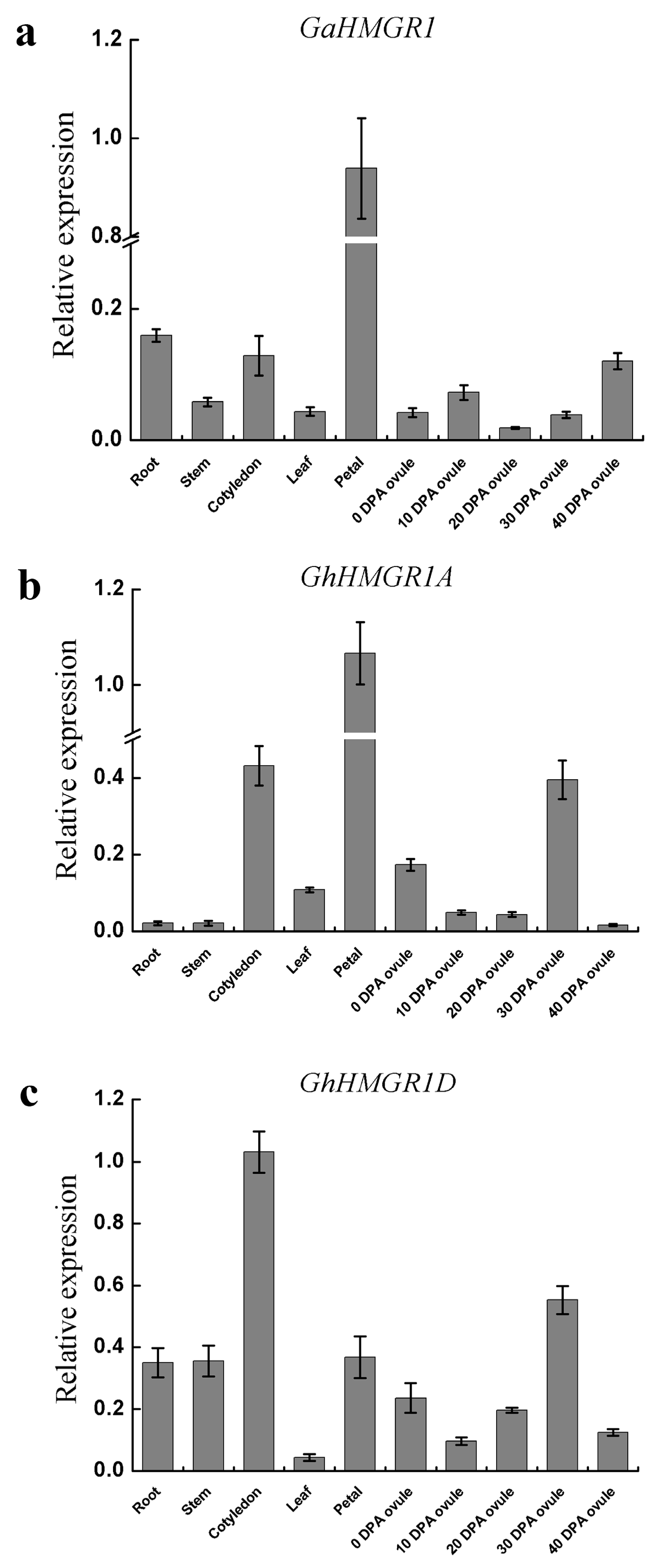
2.4. Identification of HMGR Pseudogenes
3. Discussion
4. Materials and Methods
4.1. Identification of Genes in the MVA Pathway in Gossypium
4.2. Sequence Alignment and Phylogenetic Tree Construction
4.3. Chromosomal Mapping, Protein Motif, and Gene Structure Analysis
4.4. Cotton Plant Growth and Sample Collection
4.5. RNA Isolation and cDNA Synthesis
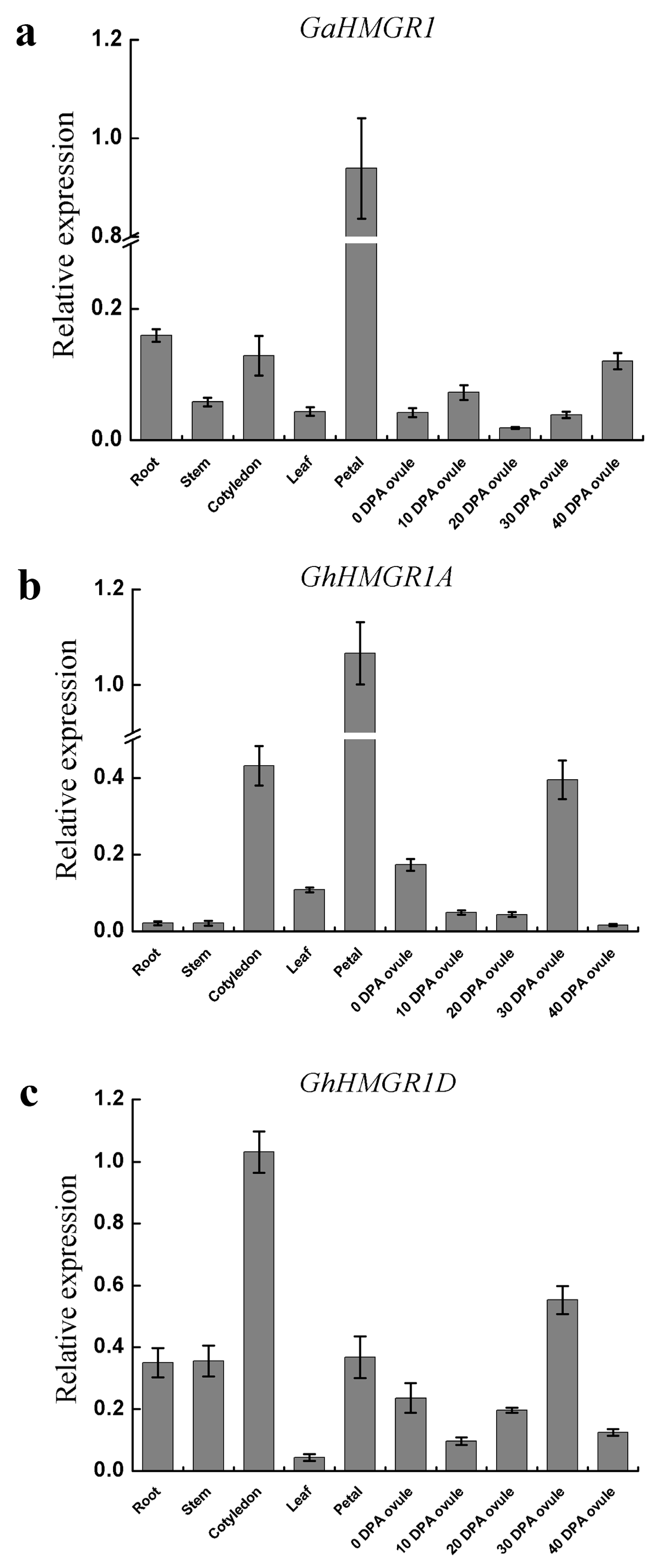
4.6. Reverse Transcription PCR (RT-PCR) and Quantitative Real-Time RT-PCR (qRT-PCR)
5. Conclusions
Supplementary Materials
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Withers, S.T.; Keasling, J.D. Biosynthesis and engineering of isoprenoid small molecules. Appl. Microbiol. Biotechnol. 2007, 73, 980–990. [Google Scholar] [CrossRef] [PubMed]
- Lange, B.M.; Rujan, T.; Martin, W.; Croteau, R. Isoprenoid biosynthesis: The evolution of two ancient and distinct pathways across genomes. Proc. Natl. Acad. Sci. USA 2000, 97, 13172–13177. [Google Scholar] [CrossRef] [PubMed]
- Nicotra, A.B.; Atkin, O.K.; Bonser, S.P.; Davidson, A.M.; Finnegan, E.J.; Mathesius, U.; Poot, P.; Purugganan, M.D.; Richards, C.L.; Valladares, F. Plant phenotypic plasticity in a changing climate. Trends Plant Sci. 2010, 15, 684–692. [Google Scholar] [CrossRef] [PubMed]
- Bouvier, F.; Rahier, A.; Camara, B. Biogenesis, molecular regulation and function of plant isoprenoids. Prog. Lipid Res. 2005, 44, 357–429. [Google Scholar] [CrossRef] [PubMed]
- Bohlmann, J.; Keeling, C.I. Terpenoid biomaterials. Plant J. 2008, 54, 656–669. [Google Scholar] [CrossRef] [PubMed]
- Singh, B.; Sharma, R.A. Plant terpenes: Defense responses, phylogenetic analysis, regulation and clinical applications. 3 Biotech 2015, 5, 129–151. [Google Scholar] [CrossRef] [PubMed]
- Bick, J.A.; Lange, B.M. Metabolic cross talk between cytosolic and plastidial pathways of isoprenoid biosynthesis: Unidirectional transport of intermediates across the chloroplast envelope membrane. Arch. Biochem. Biophys. 2003, 415, 146–154. [Google Scholar] [CrossRef]
- Laule, O.; Furholz, A.; Chang, H.S.; Zhu, T.; Wang, X.; Heifetz, P.B.; Gruissem, W.; Lange, M. Crosstalk between cytosolic and plastidial pathways of isoprenoid biosynthesis in Arabidopsis thaliana. Proc. Natl. Acad. Sci. USA 2003, 100, 6866–6871. [Google Scholar] [CrossRef] [PubMed]
- Rodríguez-Concepción, M. Early Steps in Isoprenoid Biosynthesis: Multilevel Regulation of the Supply of Common Precursors in Plant Cells. Phytochem. Rev. 2006, 5, 1–15. [Google Scholar] [CrossRef]
- Hemmerlin, A.; Harwood, J.L.; Bach, T.J. A raison d’etre for two distinct pathways in the early steps of plant isoprenoid biosynthesis? Prog. Lipid Res. 2012, 51, 95–148. [Google Scholar] [CrossRef] [PubMed]
- Learned, R.M.; Fink, G.R. 3-Hydroxy-3-methylglutaryl-coenzyme A reductase from Arabidopsis thaliana is structurally distinct from the yeast and animal enzymes. Proc. Natl. Acad. Sci. USA 1989, 86, 2779–2783. [Google Scholar] [CrossRef] [PubMed]
- Enjuto, M.; Balcells, L.; Campos, N.; Caelles, C.; Arro, M.; Boronat, A. Arabidopsis thaliana contains two differentially expressed 3-hydroxy-3-methylglutaryl-CoA reductase genes, which encode microsomal forms of the enzyme. Proc. Natl. Acad. Sci. USA 1994, 91, 927–931. [Google Scholar] [CrossRef] [PubMed]
- Ha, S.H.; Lee, S.W.; Kim, Y.M.; Hwang, Y.S. Molecular characterization of Hmg2 gene encoding a 3-hydroxy-methylglutaryl-CoA reductase in rice. Mol. Cells 2001, 11, 295–302. [Google Scholar] [PubMed]
- Aoyagi, K.; Beyou, A.; Moon, K.; Fang, L.; Ulrich, T. Isolation and characterization of cDNAs encoding wheat 3-hydroxy-3-methylglutaryl coenzyme A reductase. Plant Physiol. 1993, 102, 623–628. [Google Scholar] [CrossRef] [PubMed]
- Loguercio, L.L.; Scott, H.C.; Trolinder, N.L.; Wilkins, T.A. Hmg-coA reductase gene family in cotton (Gossypium hirsutum L.): Unique structural features and differential expression of hmg2 potentially associated with synthesis of specific isoprenoids in developing embryos. Plant Cell Physiol. 1999, 40, 750–761. [Google Scholar] [CrossRef] [PubMed]
- Kobayashi, T.; Kato-Emori, S.; Tomita, K.; Ezura, H. Detection of 3-hydroxy-3-methylglutaryl-coenzyme A reductase protein Cm-HMGR during fruit development in melon (Cucumis melo L.). Theor. Appl. Genet. 2002, 104, 779–785. [Google Scholar] [PubMed]
- Devi, K.; Patar, L.; Modi, M.K.; Sen, P. An Insight into Structure, Function, and Expression Analysis of 3-Hydroxy-3-Methylglutaryl-CoA Reductase of Cymbopogon winterianus. Bioinform. Biol. Insights 2017, 11, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Gu, W.; Geng, C.; Xue, W.; Wu, Q.; Chao, J.; Xu, F.; Sun, H.; Jiang, L.; Han, Y.; Zhang, S. Characterization and function of the 3-hydroxy-3-methylglutaryl-CoA reductase gene in Alisma orientale (Sam.) Juz. and its relationship with protostane triterpene production. Plant Physiol. Biochem. 2015, 97, 378–389. [Google Scholar] [CrossRef] [PubMed]
- Schaller, H.; Grausem, B.; Benveniste, P.; Chye, M.L.; Tan, C.T.; Song, Y.H.; Chua, N.H. Expression of the Hevea brasiliensis (H.B.K.) Mull. Arg. 3-Hydroxy-3-Methylglutaryl-Coenzyme a Reductase 1 in Tobacco Results in Sterol Overproduction. Plant Physiol. 1995, 109, 761–770. [Google Scholar] [CrossRef] [PubMed]
- Kim, Y.J.; Lee, O.R.; Oh, J.Y.; Jang, M.G.; Yang, D.C. Functional analysis of 3-hydroxy-3-methylglutaryl coenzyme a reductase encoding genes in triterpene saponin-producing ginseng. Plant Physiol. 2014, 165, 373–387. [Google Scholar] [CrossRef] [PubMed]
- Munoz-Bertomeu, J.; Sales, E.; Ros, R.; Arrillaga, I.; Segura, J. Up-regulation of an N-terminal truncated 3-hydroxy-3-methylglutaryl CoA reductase enhances production of essential oils and sterols in transgenic Lavandula latifolia. Plant Biotechnol. J. 2007, 5, 746–758. [Google Scholar] [CrossRef] [PubMed]
- Leivar, P.; Antolin-Llovera, M.; Ferrero, S.; Closa, M.; Arro, M.; Ferrer, A.; Boronat, A.; Campos, N. Multilevel control of Arabidopsis 3-hydroxy-3-methylglutaryl coenzyme A reductase by protein phosphatase 2A. Plant Cell 2011, 23, 1494–1511. [Google Scholar] [CrossRef] [PubMed]
- Lv, D.M.; Zhang, T.T.; Deng, S.; Zhang, Y.H. Functional analysis of the Malus domestica MdHMGR2 gene promoter in transgenic Arabidopsis thaliana. Biol. Plant. 2016, 60, 667–676. [Google Scholar] [CrossRef]
- Lv, D.; Zhang, Y. Isolation and functional analysis of apple MdHMGR1 and MdHMGR4 gene promoters in transgenic Arabidopsis thaliana. Plant Cell Tissue Organ Cult. 2017, 129, 133–143. [Google Scholar] [CrossRef]
- Morshedloo, M.R.; Craker, L.E.; Salami, A.; Nazeri, V.; Sang, H.; Maggi, F. Effect of prolonged water stress on essential oil content, compositions and gene expression patterns of mono- and sesquiterpene synthesis in two oregano (Origanum vulgare L.) subspecies. Plant Physiol. Biochem. 2017, 111, 119–128. [Google Scholar] [CrossRef] [PubMed]
- Wendel, J.F.; Cronn, R.C. Polyploidy and the evolutionary history of cotton. Adv. Agron. 2003, 78, 139–186. [Google Scholar]
- Paterson, A.H.; Wendel, J.F.; Gundlach, H.; Guo, H.; Jenkins, J.; Jin, D.; Llewellyn, D.; Showmaker, K.C.; Shu, S.; Udall, J. Repeated polyploidization of Gossypium genomes and the evolution of spinnable cotton fibres. Nature 2012, 492, 423–427. [Google Scholar] [CrossRef] [PubMed]
- Wang, K.; Wang, Z.; Li, F.; Ye, W.; Wang, J.; Song, G.; Yue, Z.; Cong, L.; Shang, H.; Zhu, S. The draft genome of a diploid cotton Gossypium raimondii. Nat. Genet. 2012, 44, 1098–1103. [Google Scholar] [CrossRef] [PubMed]
- Li, F.; Fan, G.; Wang, K.; Sun, F.; Yuan, Y.; Song, G.; Li, Q.; Ma, Z.; Lu, C.; Zou, C. Genome sequence of the cultivated cotton Gossypium arboreum. Nat. Genet. 2014, 46, 567–572. [Google Scholar] [CrossRef] [PubMed]
- Li, F.; Fan, G.; Lu, C.; Xiao, G.; Zou, C.; Kohel, R.J.; Ma, Z.; Shang, H.; Ma, X.; Wu, J. Genome sequence of cultivated Upland cotton (Gossypium hirsutum TM-1) provides insights into genome evolution. Nat. Biotechnol. 2015, 33, 524–530. [Google Scholar] [CrossRef] [PubMed]
- Zhang, T.; Hu, Y.; Jiang, W.; Fang, L.; Guan, X.; Chen, J.; Zhang, J.; Saski, C.A.; Scheffler, B.E.; Stelly, D.M. Sequencing of allotetraploid cotton (Gossypium hirsutum L. acc. TM-1) provides a resource for fiber improvement. Nat. Biotechnol. 2015, 33, 531–537. [Google Scholar] [CrossRef] [PubMed]
- Chen, Z.J.; Scheffler, B.E.; Dennis, E.; Triplett, B.A.; Zhang, T.; Guo, W.; Chen, X.; Stelly, D.M.; Rabinowicz, P.D.; Town, C.D. Toward sequencing cotton (Gossypium) genomes. Plant Physiol. 2007, 145, 1303–1310. [Google Scholar] [CrossRef] [PubMed]
- Ashraf, J.; Dongyun, Z.; Qiaolian, W.; Malik, W.; Youping, Z.; Abid, M.A.; Hailiang, C.; Qiuhong, Y.; Guoli, S. Recent Insights in Cotton Functional Genomics: Progress and Future Perspectives. Plant Biotechnol. J. 2017. [Google Scholar] [CrossRef] [PubMed]
- Tian, X.; Ruan, J.; Huang, J.; Fang, X.; Mao, Y.; Wang, L.; Chen, X.; Yang, C. Gossypol: Phytoalexin of cotton. Sci. China Life Sci. 2016, 59, 122–129. [Google Scholar] [CrossRef] [PubMed]
- Cai, Y.; Xie, Y.; Liu, J. Glandless seed and glanded plant research in cotton. A review. Agron. Sustain. Dev. 2010, 30, 181–190. [Google Scholar] [CrossRef]
- Li, W.; Liu, W.; Wei, H.; He, Q.; Chen, J.; Zhang, B.; Zhu, S. Species-specific expansion and molecular evolution of the 3-hydroxy-3-methylglutaryl coenzyme A reductase (HMGR) gene family in plants. PLoS ONE 2014, 9, e94172. [Google Scholar] [CrossRef] [PubMed]
- Yao, H.; Guo, L.; Fu, Y.; Borsuk, L.A.; Wen, T.J.; Skibbe, D.S.; Cui, X.; Scheffler, B.E.; Cao, J.; Emrich, S.J. Evaluation of five ab initio gene prediction programs for the discovery of maize genes. Plant Mol. Biol. 2005, 57, 445–460. [Google Scholar] [CrossRef] [PubMed]
- Campos, N.; Boronat, A. Targeting and topology in the membrane of plant 3-hydroxy-3-methylglutaryl coenzyme A reductase. Plant Cell 1995, 7, 2163–2174. [Google Scholar] [CrossRef] [PubMed]
- Leivar, P.; Gonzalez, V.M.; Castel, S.; Trelease, R.N.; Lopez-Iglesias, C.; Arro, M.; Boronat, A.; Campos, N.; Ferrer, A.; Fernandez-Busquets, X. Subcellular localization of Arabidopsis 3-hydroxy-3-methylglutaryl-coenzyme A reductase. Plant Physiol. 2005, 137, 57–69. [Google Scholar] [CrossRef] [PubMed]
- Istvan, E.S.; Deisenhofer, J. The structure of the catalytic portion of human HMG-CoA reductase. Biochim. Biophys. Acta 2000, 1529, 9–18. [Google Scholar] [CrossRef]
- Darabi, M.; Izadi-Darbandi, A.; Masoudi-Nejad, A.; Naghavi, M.R.; Nemat-Zadeh, G. Bioinformatics study of the 3-hydroxy-3-methylglotaryl-coenzyme A reductase (HMGR) gene in Gramineae. Mol. Biol. Rep. 2012, 39, 8925–8935. [Google Scholar] [CrossRef] [PubMed]
- Joost, O.; Bianchini, G.; Bell, A.A.; Benedict, C.R.; Magill, C.W. Differential induction of 3-hydroxy-3-methylglutaryl CoA reductase in two cotton species following inoculation with Verticillium. Mol. Plant Microbe Interact. 1995, 8, 880–885. [Google Scholar] [CrossRef] [PubMed]
- Hurles, M. Gene duplication: The genomic trade in spare parts. PLoS Biol. 2004, 2, e206. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Freeling, M. Bias in plant gene content following different sorts of duplication: Tandem, whole-genome, segmental, or by transposition. Annu. Rev. Plant Biol. 2009, 60, 433–453. [Google Scholar] [CrossRef] [PubMed]
- Du, H.; Yang, S.S.; Liang, Z.; Feng, B.R.; Liu, L.; Huang, Y.B.; Tang, Y.X. Genome-wide analysis of the MYB transcription factor superfamily in soybean. BMC Plant Biol. 2012, 12, 106. [Google Scholar] [CrossRef] [PubMed]
- He, H.; Dong, Q.; Shao, Y.; Jiang, H.; Zhu, S.; Cheng, B.; Xiang, Y. Genome-wide survey and characterization of the WRKY gene family in Populus trichocarpa. Plant Cell Rep. 2012, 31, 1199–1217. [Google Scholar] [CrossRef] [PubMed]
- Yin, G.; Xu, H.; Xiao, S.; Qin, Y.; Li, Y.; Yan, Y.; Hu, Y. The large soybean (Glycine max) WRKY TF family expanded by segmental duplication events and subsequent divergent selection among subgroups. BMC Plant Biol. 2013, 13, 148. [Google Scholar] [CrossRef] [PubMed]
- Chai, G.; Wang, Z.; Tang, X.; Yu, L.; Qi, G.; Wang, D.; Yan, X.; Kong, Y.; Zhou, G. R2R3-MYB gene pairs in Populus: Evolution and contribution to secondary wall formation and flowering time. J. Exp. Bot. 2014, 65, 4255–4269. [Google Scholar] [CrossRef] [PubMed]
- Tholl, D.; Lee, S. Terpene Specialized Metabolism in Arabidopsis thaliana. In Arabidopsis Book; The American Society of Plant Biologists: Rockville, MD, USA, 2011; Volume 9, p. e0143. [Google Scholar]
- Zhou, M.; Zhang, C.; Wu, Y.; Tang, Y. Metabolic engineering of gossypol in cotton. Appl. Microbiol. Biotechnol. 2013, 97, 6159–6165. [Google Scholar] [CrossRef] [PubMed]
- Fryxell, P.A. A Redefinition of the Tribe Gossypieae. Bot. Gaz. 1968, 129, 296–308. [Google Scholar] [CrossRef]
- Lynch, M.; Conery, J.S. The evolutionary fate and consequences of duplicate genes. Science 2000, 290, 1151–1155. [Google Scholar] [CrossRef] [PubMed]
- Loguercio, L.L.; Wilkins, T.A. Structural analysis of a hmg-coA-reductase pseudogene: Insights into evolutionary processes affecting the hmgr gene family in allotetraploid cotton (Gossypium hirsutum L.). Curr. Genet. 1998, 34, 241–249. [Google Scholar] [CrossRef] [PubMed]
- Andersson, J.O.; Andersson, S.G. Pseudogenes, junk DNA, and the dynamics of Rickettsia genomes. Mol. Biol. Evol. 2001, 18, 829–839. [Google Scholar] [CrossRef] [PubMed]
- Pink, R.C.; Wicks, K.; Caley, D.P.; Punch, E.K.; Jacobs, L.; Francisco Carter, D.R. Pseudogenes: Pseudo-functional or key regulators in health and disease? RNA 2011, 17, 792–798. [Google Scholar] [CrossRef] [PubMed]
- Xiao, J.; Sekhwal, M.K.; Li, P.; Ragupathy, R.; Cloutier, S.; Wang, X.; You, F.M. Pseudogenes and Their Genome-Wide Prediction in Plants. Int. J. Mol. Sci. 2016, 17, 1991. [Google Scholar] [CrossRef] [PubMed]
- Senda, M.; Masuta, C.; Ohnishi, S.; Goto, K.; Kasai, A.; Sano, T.; Hong, J.S.; MacFarlane, S. Patterning of virus-infected Glycine max seed coat is associated with suppression of endogenous silencing of chalcone synthase genes. Plant Cell 2004, 16, 807–818. [Google Scholar] [CrossRef] [PubMed]
- Kasai, A.; Kasai, K.; Yumoto, S.; Senda, M. Structural features of GmIRCHS, candidate of the I gene inhibiting seed coat pigmentation in soybean: Implications for inducing endogenous RNA silencing of chalcone synthase genes. Plant Mol. Biol. 2007, 64, 467–479. [Google Scholar] [CrossRef] [PubMed]
- Tuteja, J.H.; Zabala, G.; Varala, K.; Hudson, M.; Vodkin, L.O. Endogenous, tissue-specific short interfering RNAs silence the chalcone synthase gene family in glycine max seed coats. Plant Cell 2009, 21, 3063–3077. [Google Scholar] [CrossRef] [PubMed]
- Poliseno, L.; Salmena, L.; Zhang, J.; Carver, B.; Haveman, W.J.; Pandolfi, P.P. A coding-independent function of gene and pseudogene mRNAs regulates tumour biology. Nature 2010, 465, 1033–1038. [Google Scholar] [CrossRef] [PubMed]
- Nagegowda, D.A.; Gutensohn, M.; Wilkerson, C.G.; Dudareva, N. Two nearly identical terpene synthases catalyze the formation of nerolidol and linalool in snapdragon flowers. Plant J. 2008, 55, 224–239. [Google Scholar] [CrossRef] [PubMed]
- Yue, Y.; Yu, R.; Fan, Y. Transcriptome profiling provides new insights into the formation of floral scent in Hedychium coronarium. BMC Genom. 2015, 16, 470. [Google Scholar] [CrossRef] [PubMed]
- Nieuwenhuizen, N.J.; Wang, M.Y.; Matich, A.J.; Green, S.A.; Chen, X.; Yauk, Y.K.; Beuning, L.L.; Nagegowda, D.A.; Dudareva, N.; Atkinson, R.G. Two terpene synthases are responsible for the major sesquiterpenes emitted from the flowers of kiwifruit (Actinidia deliciosa). J. Exp. Bot. 2009, 60, 3203–3219. [Google Scholar] [CrossRef] [PubMed]
- Vranova, E.; Coman, D.; Gruissem, W. Network analysis of the MVA and MEP pathways for isoprenoid synthesis. Annu. Rev. Plant Biol. 2013, 64, 665–700. [Google Scholar] [CrossRef] [PubMed]
- Finn, R.D.; Coggill, P.; Eberhardt, R.Y.; Eddy, S.R.; Mistry, J.; Mitchell, A.L.; Potter, S.C.; Punta, M.; Qureshi, M.; Sangrador-Vegas, A. The Pfam protein families database: Towards a more sustainable future. Nucleic Acids Res. 2016, 44, D279–D285. [Google Scholar] [CrossRef] [PubMed]
- Finn, R.D.; Attwood, T.K.; Babbitt, P.C.; Bateman, A.; Bork, P.; Bridge, A.J.; Chang, H.Y.; Dosztanyi, Z.; El-Gebali, S.; Fraser, M. InterPro in 2017-beyond protein family and domain annotations. Nucleic Acids Res. 2017, 45, D190–D199. [Google Scholar] [CrossRef] [PubMed]
- Larkin, M.A.; Blackshields, G.; Brown, N.P.; Chenna, R.; McGettigan, P.A.; McWilliam, H.; Valentin, F.; Wallace, I.M.; Wilm, A.; Lopez, R. Clustal W and Clustal X version 2.0. Bioinformatics 2007, 23, 2947–2948. [Google Scholar] [CrossRef] [PubMed]
- Tamura, K.; Peterson, D.; Peterson, N.; Stecher, G.; Nei, M.; Kumar, S. MEGA5: Molecular evolutionary genetics analysis using maximum likelihood, evolutionary distance, and maximum parsimony methods. Mol. Biol. Evol. 2011, 28, 2731–2739. [Google Scholar] [CrossRef] [PubMed]
- Liu, W.; Li, W.; He, Q.; Daud, M.K.; Chen, J.; Zhu, S. Characterization of 19 Genes Encoding Membrane-Bound Fatty Acid Desaturases and their Expression Profiles in Gossypium raimondii Under Low Temperature. PLoS ONE 2015, 10, e0123281. [Google Scholar] [CrossRef] [PubMed]
- Krzywinski, M.; Schein, J.; Birol, I.; Connors, J.; Gascoyne, R.; Horsman, D.; Jones, S.J.; Marra, M.A. Circos: An information aesthetic for comparative genomics. Genome Res. 2009, 19, 1639–1645. [Google Scholar] [CrossRef] [PubMed]
- Hu, B.; Jin, J.; Guo, A.Y.; Zhang, H.; Luo, J.; Gao, G. GSDS 2.0: An upgraded gene feature visualization server. Bioinformatics 2015, 31, 1296–1297. [Google Scholar] [CrossRef] [PubMed]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2(-Delta Delta C(T)) Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]
Sample Availability: Samples of the Gossypium species are available from the authors. |


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene Name | Gene Locus | Chromosome | Location | Strand | Protein Length | Mw(kDa) a | pI a |
|---|---|---|---|---|---|---|---|
| GhHMGR1A c | HMGR pseudogene | A01 | 39188679-39190582 | + | _ | _ | _ |
| GhHMGR2A | Gh_A03G1497 | A03 | 95297747-95299778 | − | 582 | 62.33 | 6.24 |
| GhHMGR3A b | Gh_A03G1496_1 | A03 | 95253091-95255136 | − | 585 | 62.68 | 6.14 |
| GhHMGR4A b | Gh_A03G1496_2 | A03 | 95195182-95197211 | − | 585 | 62.67 | 6.00 |
| GhHMGR5A | Gh_A03G1495 | A03 | 95169197-95171221 | − | 585 | 62.71 | 6.25 |
| GhHMGR6A | Gh_A01G2017 | scaffold31_A01 | 38993-42152 | + | 585 | 62.82 | 6.20 |
| GhHMGR7A | Gh_A12G0103 | A12 | 1425978-1428020 | − | 585 | 62.62 | 6.24 |
| GhHMGR8A b | Gh_A04G1424 | scaffold1012_A04 | 184892-187043 | − | 581 | 62.26 | 6.69 |
| GhHMGR9A | Gh_A13G0557 | A13 | 13047920-13050876 | − | 628 | 67.60 | 6.26 |
| GhHMGR1D | Gh_D01G1158 | D01 | 25640487-25642401 | + | 560 | 60.40 | 6.53 |
| GhHMGR2D | Gh_D02G1965 | D02 | 63580366-63582399 | − | 582 | 62.33 | 6.43 |
| GhHMGR3D | Gh_D02G1964 | D02 | 63566380-63569291 | − | 585 | 62.54 | 6.00 |
| GhHMGR4D | Gh_D02G1963 | D02 | 63558389-63560423 | − | 583 | 62.40 | 6.17 |
| GhHMGR5D | Gh_D02G1962 | D02 | 63549091-63551149 | − | 585 | 62.78 | 6.25 |
| GhHMGR6D | Gh_D01G0134 | D01 | 984873-987923 | + | 585 | 62.79 | 5.83 |
| GhHMGR7D | Gh_D12G0115 | D12 | 1451036-1453088 | − | 585 | 62.44 | 6.43 |
| GhHMGR8D b | Gh_D04G2012 | scaffold3981_D04 | 28507-30658 | + | 581 | 62.26 | 6.49 |
| GhHMGR9D | Gh_D13G0573 | D13 | 7846186-7848822 | + | 628 | 67.46 | 6.50 |
| Species | Type | Genomic Group | HMGR Pseudogene |
|---|---|---|---|
| G. raimondii | wild species | D5 | Non-existence |
| G. herbaceum race. africanum | wild species | A1 | Existence |
| G. herbaceum cv. Jinta | cultivar | A1 | Existence |
| G. arboreum cv. Shixiya1 | cultivar | A2 | Existence |
| G. darwinii | wild species | (AD)5 | Existence |
| G. mustelinum | wild species | (AD)4 | Existence |
| G. hirsutum race. latifolium | semi-domesticated species | (AD)1 | Existence |
| G. hirsutum cv. TM-1 | cultivar | (AD)1 | Existence |
| G. hirsutum cv. CCRI41 | cultivar | (AD)1 | Existence |
| G. barbadense cv. Xinhai21 | cultivar | (AD)2 | Existence |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liu, W.; Zhang, Z.; Li, W.; Zhu, W.; Ren, Z.; Wang, Z.; Li, L.; Jia, L.; Zhu, S.; Ma, Z. Genome-Wide Identification and Comparative Analysis of the 3-Hydroxy-3-methylglutaryl Coenzyme A Reductase (HMGR) Gene Family in Gossypium. Molecules 2018, 23, 193. https://doi.org/10.3390/molecules23020193
Liu W, Zhang Z, Li W, Zhu W, Ren Z, Wang Z, Li L, Jia L, Zhu S, Ma Z. Genome-Wide Identification and Comparative Analysis of the 3-Hydroxy-3-methylglutaryl Coenzyme A Reductase (HMGR) Gene Family in Gossypium. Molecules. 2018; 23(2):193. https://doi.org/10.3390/molecules23020193
Chicago/Turabian StyleLiu, Wei, Zhiqiang Zhang, Wei Li, Wei Zhu, Zhongying Ren, Zhenyu Wang, Lingli Li, Lin Jia, Shuijin Zhu, and Zongbin Ma. 2018. "Genome-Wide Identification and Comparative Analysis of the 3-Hydroxy-3-methylglutaryl Coenzyme A Reductase (HMGR) Gene Family in Gossypium" Molecules 23, no. 2: 193. https://doi.org/10.3390/molecules23020193