Wound Healing Potential of Chlorogenic Acid and Myricetin-3-O-β-Rhamnoside Isolated from Parrotia persica

 , and
, and
Abstract
:1. Introduction
2. Results
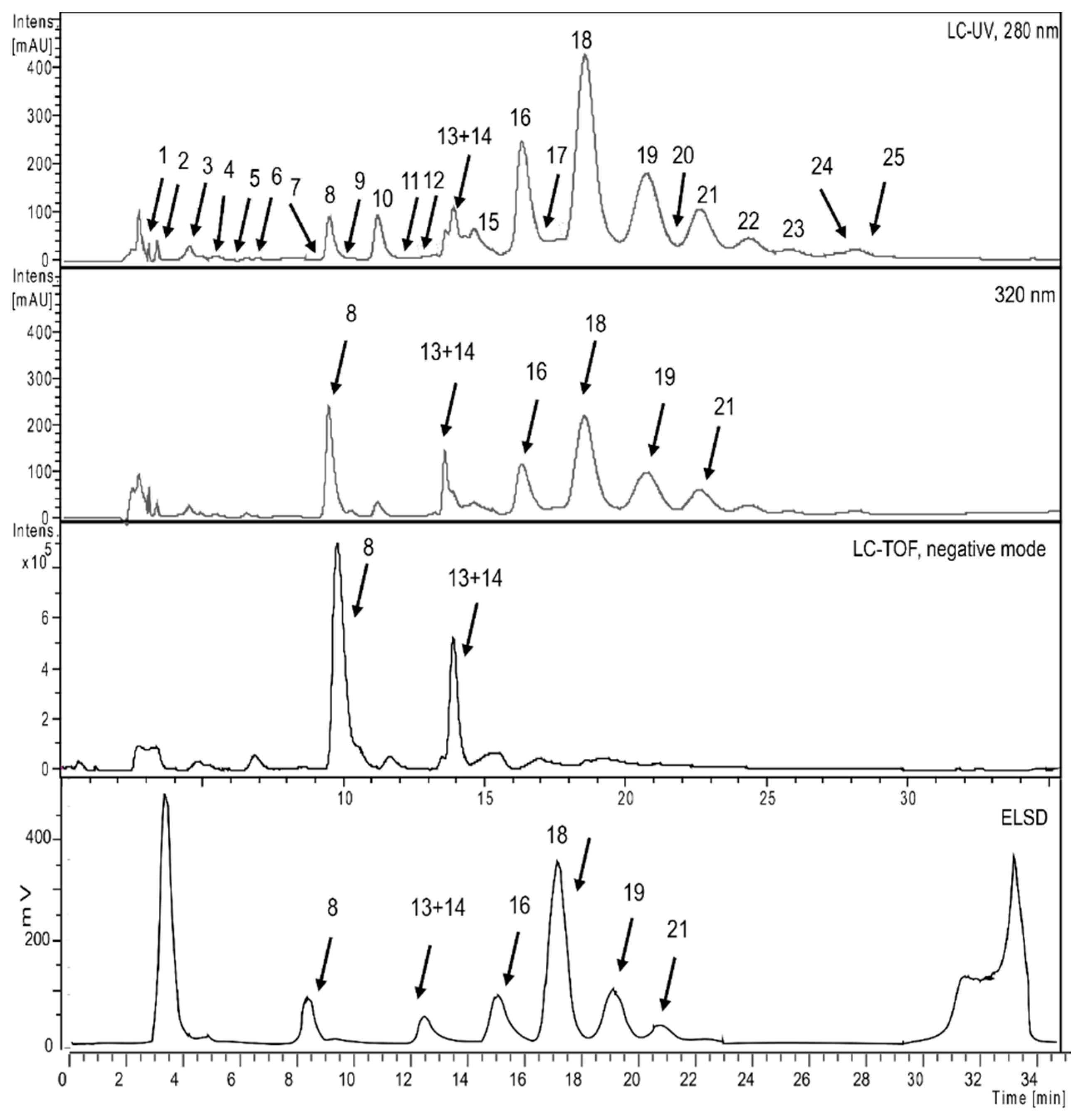
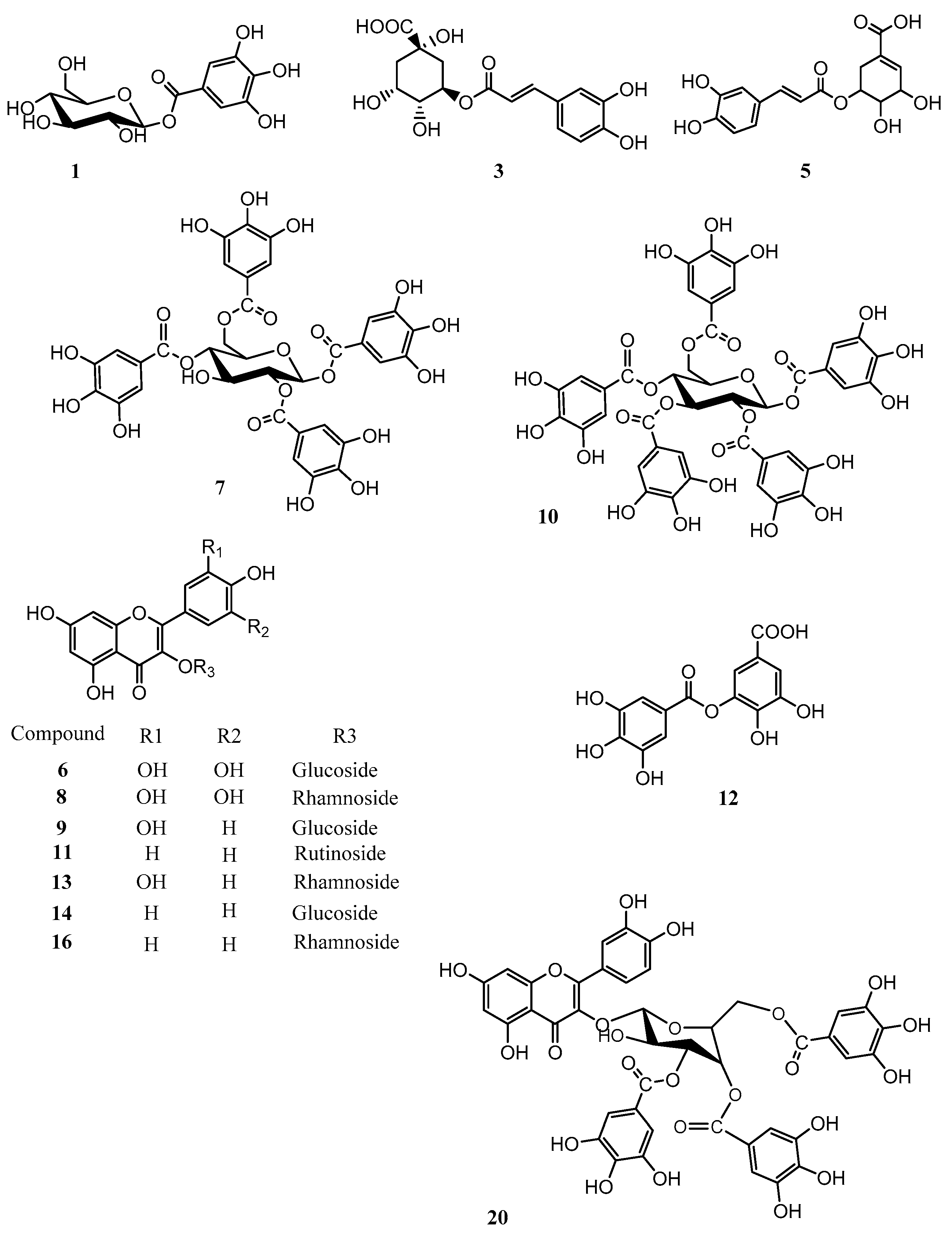
2.1. Phytochemical Profiling of P. persica
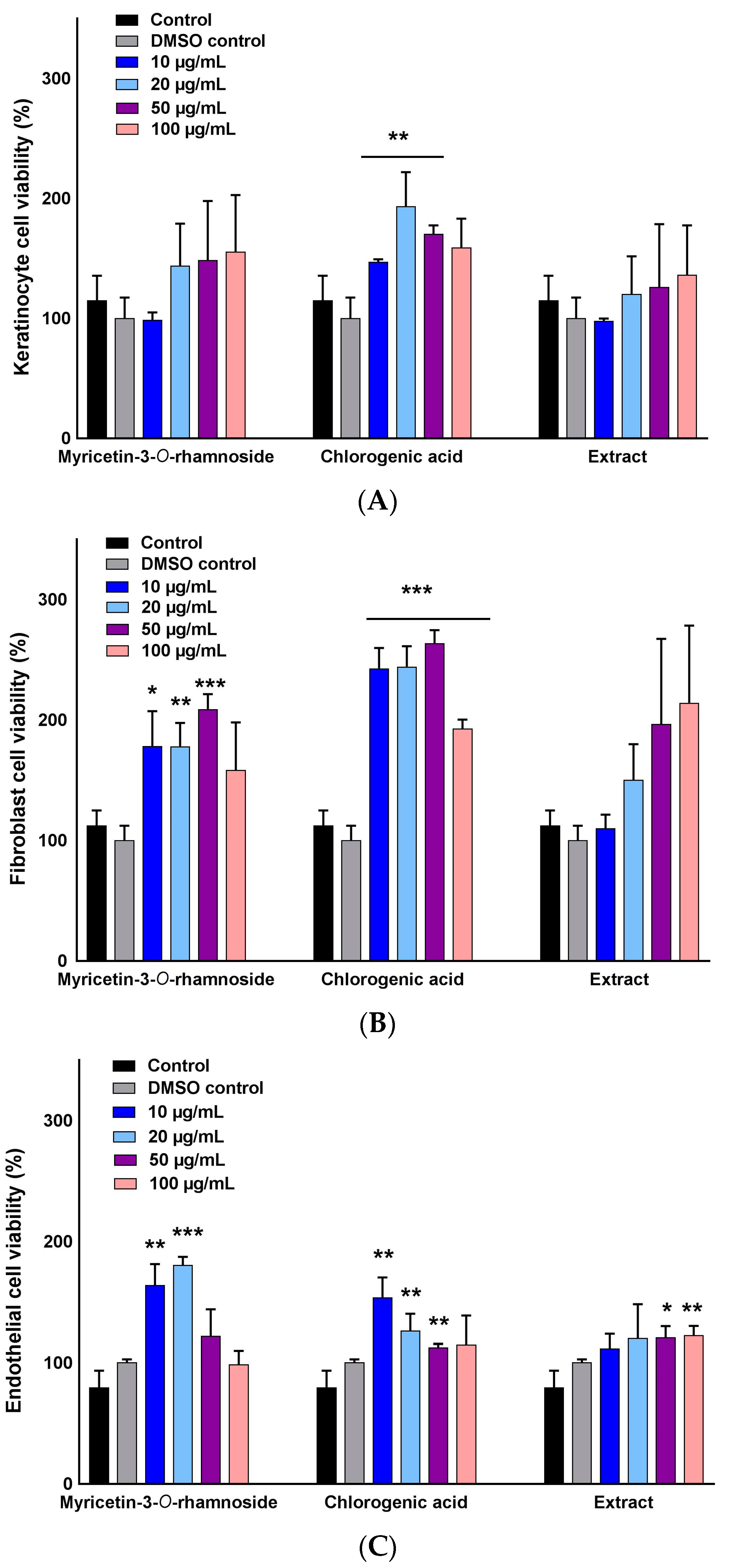
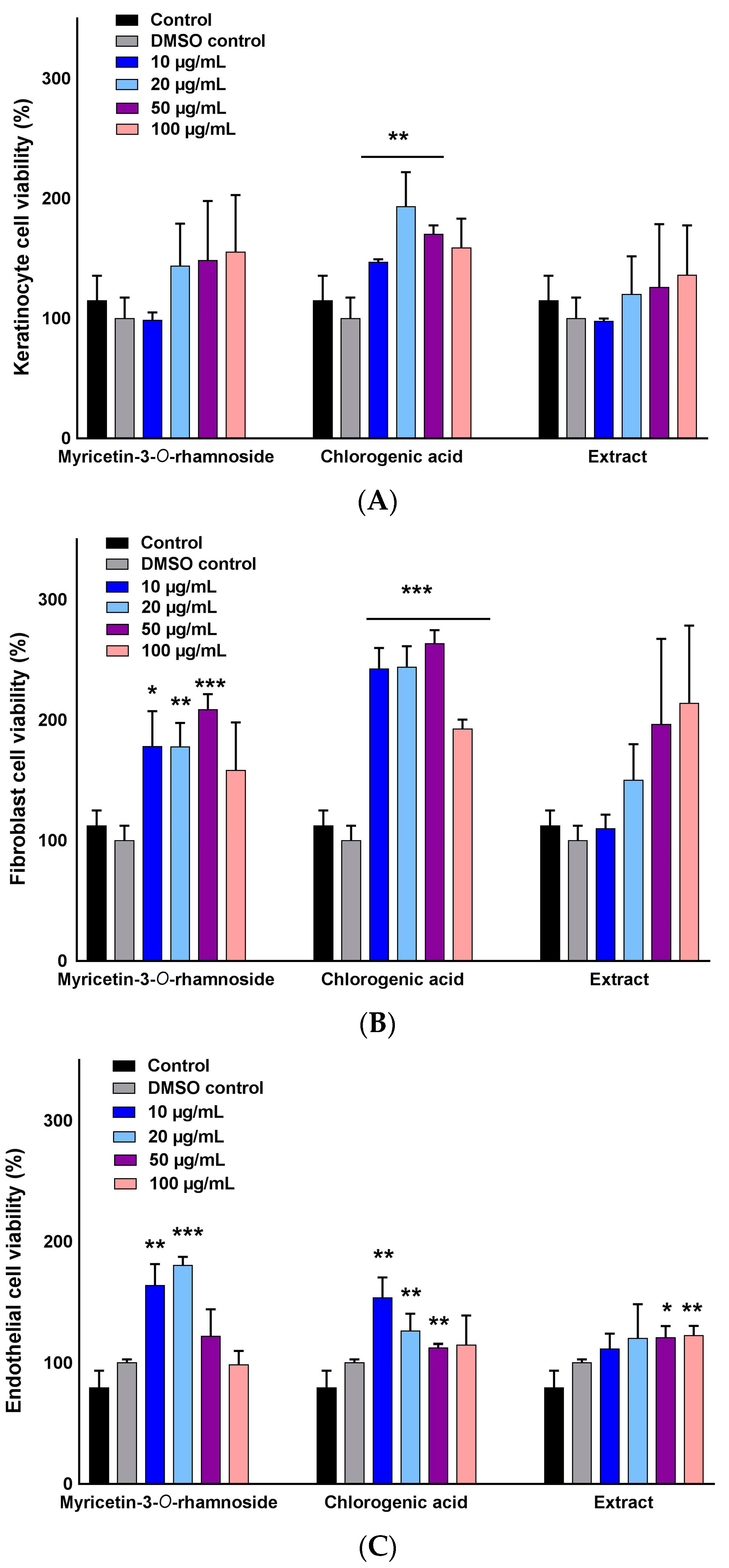
2.2. Selective Stimulation of Cell Growth
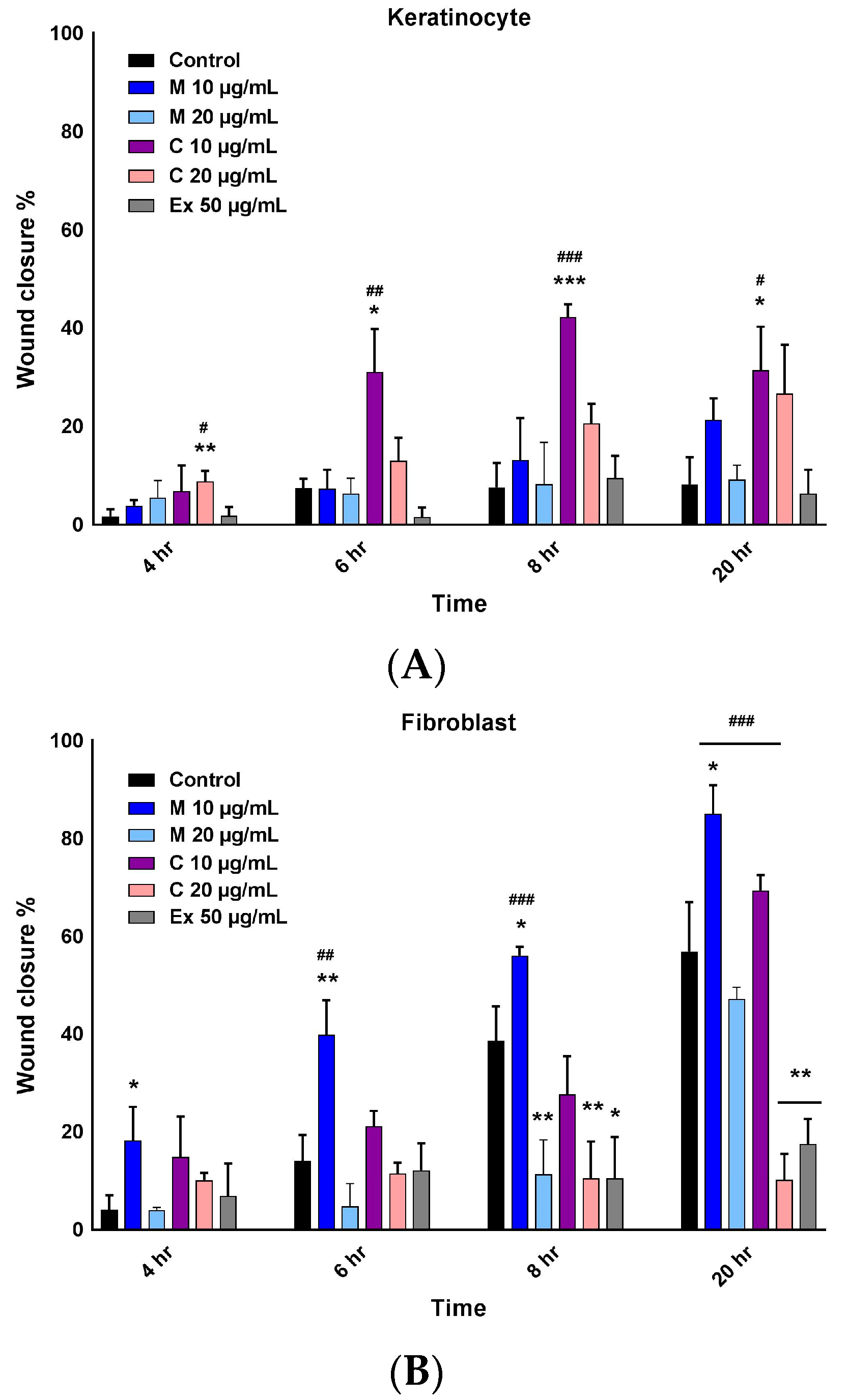
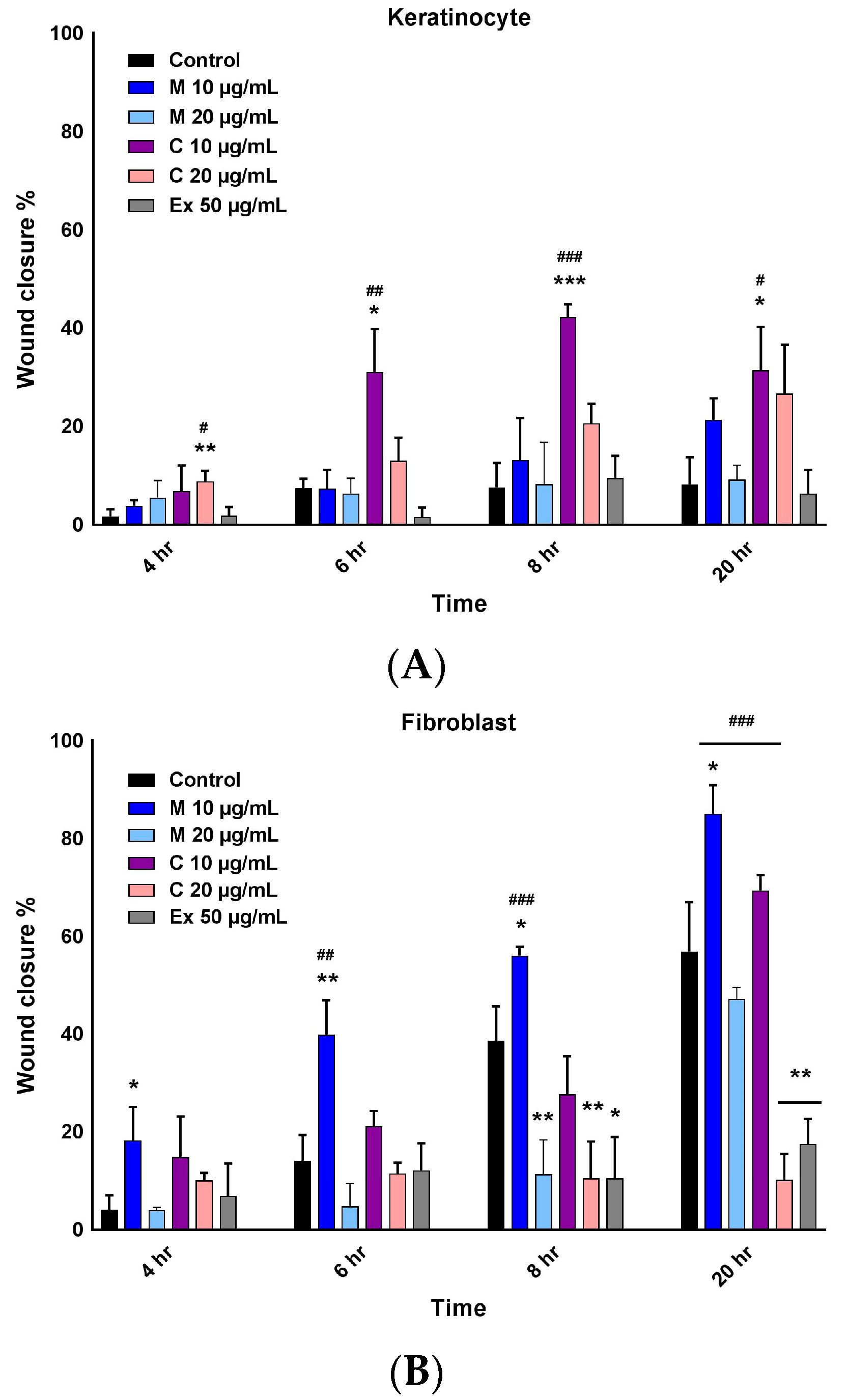
2.3. Wound Healing Activity of Myricetin-3-O-β-Rhamnoside and Chlorogenic Acid
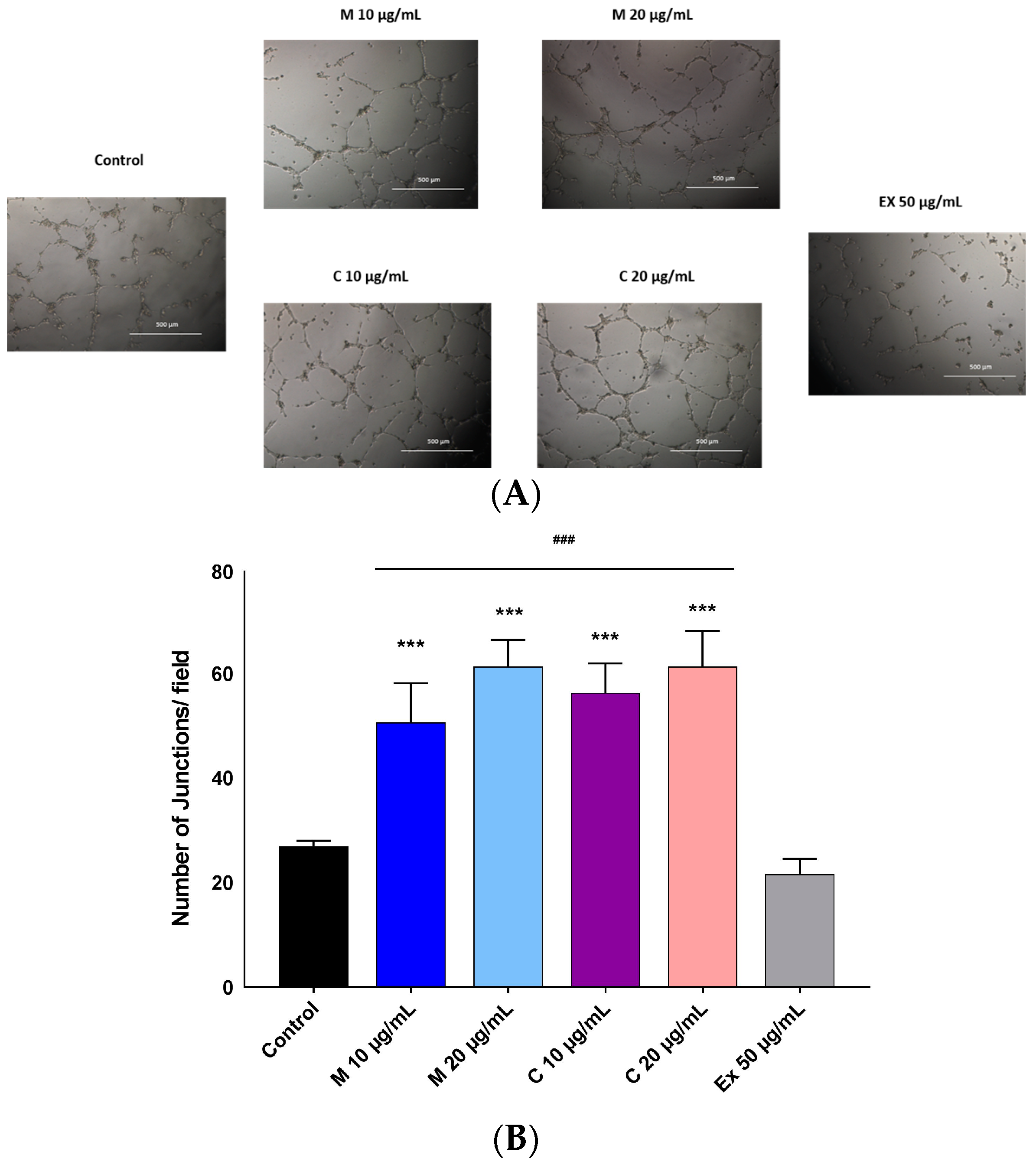
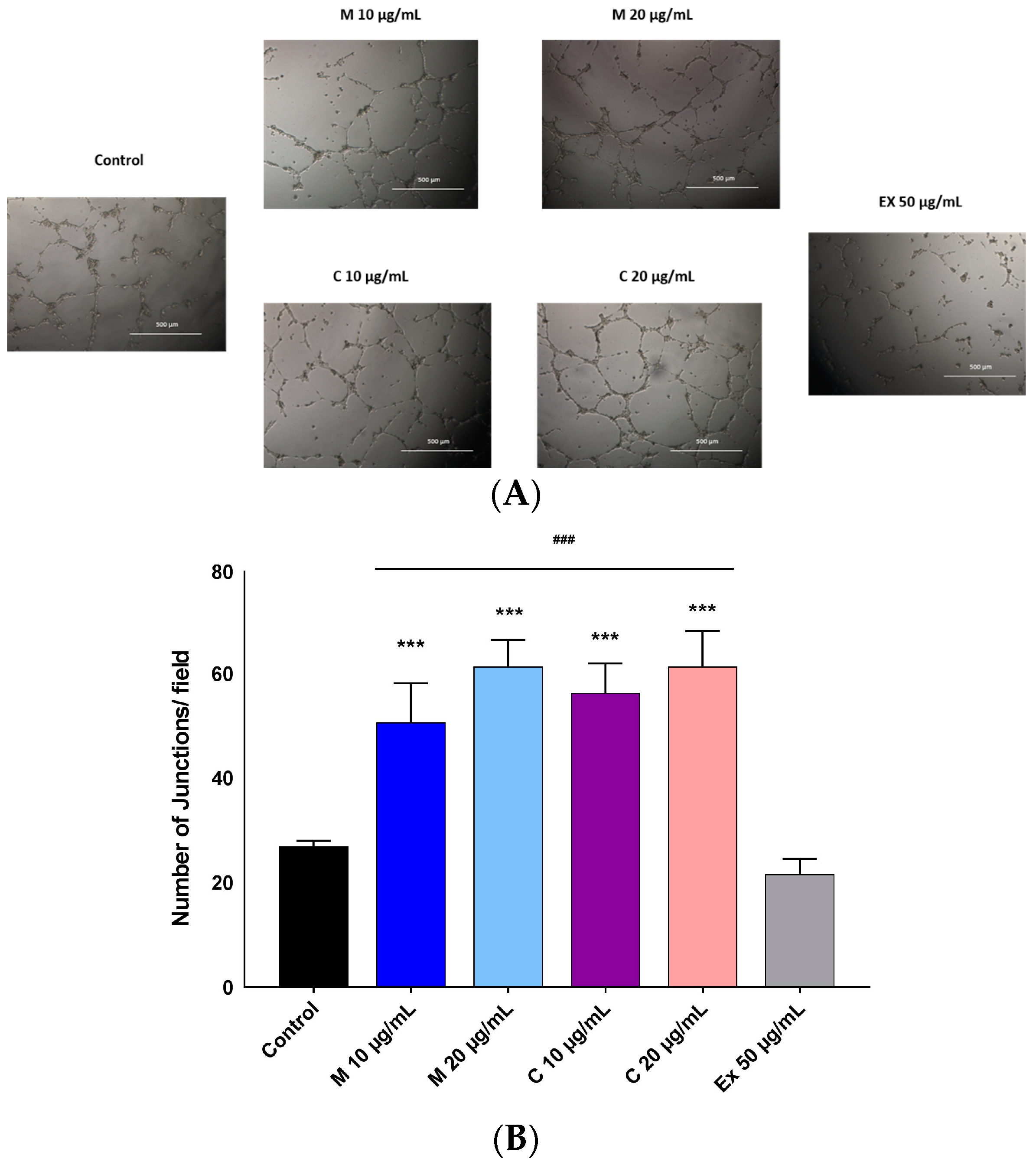
2.4. Pro-Angiogenic Effects of Myricetin-3-O-β-Rhamnoside and Chlorogenic Acid
3. Discussion
4. Materials and Methods
4.1. Chemicals and Solvents
4.2. Cell Culture and Reagents
4.3. Instruments
4.4. Extraction and Isolation
4.5. Analysis of Cell Viability
4.6. In Vitro Scratch Assay
4.7. Capillary Tube Formation
4.8. Statistical Analysis
5. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Gurtner, G.C.; Werner, S.; Barrandon, Y.; Longaker, M.T. Wound repair and regeneration. Nature 2008, 453, 314–321. [Google Scholar] [CrossRef] [PubMed]
- Thakur, R.; Jain, N.; Pathak, R.; Sandhu, S.S. Practices in wound healing studies of plants. Evid. Based Complement. Altern. Med. 2011, 2011. [Google Scholar] [CrossRef] [PubMed]
- Singer, A.J.; Clark, R.A. Cutaneous wound healing. N. Engl. J. Med 1999, 341, 738–746. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.; Wang, Y.; Farhangfar, F.; Zimmer, M.; Zhang, Y. Enhanced keratinocyte proliferation and migration in co-culture with fibroblasts. PLoS ONE 2012, 7, e40951. [Google Scholar] [CrossRef] [PubMed]
- Cheng, F.; Shen, Y.; Mohanasundaram, P.; Lindström, M.; Ivaska, J.; Ny, T.; Eriksson, J.E. Vimentin coordinates fibroblast proliferation and keratinocyte differentiation in wound healing via tgf-β-slug signaling. Proc. Nat. Acad. Sci. USA 2016, 113, E4320–E4327. [Google Scholar] [CrossRef] [PubMed]
- Pastar, I.; Stojadinovic, O.; Yin, N.C.; Ramirez, H.; Nusbaum, A.G.; Sawaya, A.; Patel, S.B.; Khalid, L.; Isseroff, R.R.; Tomic-Canic, M. Epithelialization in wound healing: A comprehensive review. Adv. Wound Care 2014, 3, 445–464. [Google Scholar] [CrossRef] [PubMed]
- Ucuzian, A.A.; Gassman, A.A.; East, A.T.; Greisler, H.P. Molecular mediators of angiogenesis. J. Burn Care Res. Off. Publ. Am. Burn Assoc. 2010, 31, 158. [Google Scholar] [CrossRef] [PubMed]
- Skiles, M.L.; Hanna, B.; Rucker, L.; Tipton, A.; Brougham-Cook, A.; Jabbarzadeh, E.; Blanchette, J.O. Asc spheroid geometry and culture oxygenation differentially impact induction of preangiogenic behaviors in endothelial cells. Cell Trans. 2015, 24, 2323–2335. [Google Scholar] [CrossRef] [PubMed]
- Harris, G.M.; Rutledge, K.; Cheng, Q.; Blanchette, J.; Jabbarzadeh, E. Strategies to direct angiogenesis within scaffolds for bone tissue engineering. Curr. Pharm. Des. 2013, 19, 3456–3465. [Google Scholar] [CrossRef] [PubMed]
- Werner, S.; Grose, R. Regulation of wound healing by growth factors and cytokines. Physiol. Rev. 2003, 83, 835–870. [Google Scholar] [PubMed]
- Werner, S.; Krieg, T.; Smola, H. Keratinocyte–fibroblast interactions in wound healing. J. Investig. Dermatol 2007, 127, 998–1008. [Google Scholar] [CrossRef] [PubMed]
- Chen, W.-C.; Liou, S.-S.; Tzeng, T.-F.; Lee, S.-L.; Liu, I.-M. Effect of topical application of chlorogenic acid on excision wound healing in rats. Planta Med. 2013, 79, 616–621. [Google Scholar] [CrossRef] [PubMed]
- Bodnar, R.J. Chemokine regulation of angiogenesis during wound healing. Adv Wound Care (New Rochelle) 2015, 4, 641–650. [Google Scholar] [CrossRef] [PubMed]
- Senet, P. [becaplermin gel (regranex gel)]. Ann. Dermatol. Venereol. 2004, 131, 351–358. [Google Scholar] [CrossRef]
- Walker, M.; Parsons, D. The biological fate of silver ions following the use of silver-containing wound care products—A review. Int. Wound J 2014, 11, 496–504. [Google Scholar] [CrossRef] [PubMed]
- Pereira, R.F.; Bartolo, P.J. Traditional therapies for skin wound healing. Adv. Wound Care 2016, 5, 208–229. [Google Scholar] [CrossRef] [PubMed]
- Koeberle, A.; Werz, O. Multi-target approach for natural products in inflammation. Drug Discov. Today 2014, 19, 1871–1882. [Google Scholar] [CrossRef] [PubMed]
- Rodrigues, T.; Reker, D.; Schneider, P.; Schneider, G. Counting on natural products for drug design. Nat. Chem. 2016, 8, 531–541. [Google Scholar] [CrossRef] [PubMed]
- Tsala, D.E.; Amadou, D.; Habtemariam, S. Natural wound healing and bioactive natural products. Phytopharmacology 2013, 4, 532–560. [Google Scholar]
- Tundis, R.; Loizzo, M.; Menichini, F. Natural products as α-amylase and α-glucosidase inhibitors and their hypoglycaemic potential in the treatment of diabetes: An update. Mini Rev. Med. Chem. 2010, 10, 315–331. [Google Scholar] [CrossRef] [PubMed]
- Dzialo, M.; Mierziak, J.; Korzun, U.; Preisner, M.; Szopa, J.; Kulma, A. The potential of plant phenolics in prevention and therapy of skin disorders. Int. J. Mol. Sci. 2016, 17, 160. [Google Scholar] [CrossRef] [PubMed]
- Korkina, L.; De Luca, C.; Pastore, S. Plant polyphenols and human skin: Friends or foes. Ann N. Y. Acad. Sci. 2012, 1259, 77–86. [Google Scholar] [CrossRef] [PubMed]
- Martinotti, S.; Ranzato, E. Propolis: A new frontier for wound healing? Burns Trauma 2015, 3, 9. [Google Scholar] [CrossRef] [PubMed]
- Paszkiewicz, M.; Budzynska, A.; Rozalska, B.; Sadowska, B. The immunomodulatory role of plant polyphenols. Postepy Hig. Med. Dosw. (Online) 2012, 66, 637–646. [Google Scholar] [CrossRef] [PubMed]
- Bui, N.T.; Ho, M.T.; Kim, Y.M.; Lim, Y.; Cho, M. Flavonoids promoting hacat migration: Ii. Molecular mechanism of 4′,6,7-trimethoxyisoflavone via nox2 activation. Phytomedicine 2014, 21, 570–577. [Google Scholar] [CrossRef] [PubMed]
- Samoylenko, A.; Hossain, J.A.; Mennerich, D.; Kellokumpu, S.; Hiltunen, J.K.; Kietzmann, T. Nutritional countermeasures targeting reactive oxygen species in cancer: From mechanisms to biomarkers and clinical evidence. Antioxid. Redox Signal. 2013, 19, 2157–2196. [Google Scholar] [CrossRef] [PubMed]
- Mukherjee, H.; Ojha, D.; Bharitkar, Y.P.; Ghosh, S.; Mondal, S.; Kaity, S.; Dutta, S.; Samanta, A.; Chatterjee, T.K.; Chakrabarti, S. Evaluation of the wound healing activity of shorea robusta, an indian ethnomedicine, and its isolated constituent (s) in topical formulation. J. Ethnopharmacol. 2013, 149, 335–343. [Google Scholar] [CrossRef] [PubMed]
- Karim, A.A.; Azlan, A.; Ismail, A.; Hashim, P.; Abd Gani, S.S.; Zainudin, B.H.; Abdullah, N.A. Phenolic composition, antioxidant, anti-wrinkles and tyrosinase inhibitory activities of cocoa pod extract. BMC Complement. Altern. Med. 2014, 14, 381. [Google Scholar]
- Wu, T.; He, M.; Zang, X.; Zhou, Y.; Qiu, T.; Pan, S.; Xu, X. A structure-activity relationship study of flavonoids as inhibitors of e. Coli by membrane interaction effect. Biochim. Biophys. Acta 2013, 1828, 2751–2756. [Google Scholar] [CrossRef] [PubMed]
- Ebrahimzadeh, M.; Nabavi, S.; Nabavi, S.; Pourmorad, F. Nitric oxide radical scavenging potential of some elburz medicinal plants. Afr. J. Biotechnol. 2010, 9, 5212–5217. [Google Scholar]
- Sahranavard, S.; Naghibi, F.; Mosaddegh, M.; Esmaeili, S.; Sarkhail, P.; Taghvaei, M.; Ghafari, S. Cytotoxic activities of selected medicinal plants from iran and phytochemical evaluation of the most potent extract. Res. Pharm. Sci. 2009, 4, 133. [Google Scholar] [PubMed]
- Mohammad, A.; Mohana, D.; Raveesha, K.; Azadbakht, M. Antibacterial potential of extracts of leaves of Parrotia persica. Afr. J. Biotechnol. 2007, 6, 2526–2528. [Google Scholar] [CrossRef]
- Deters, A.; Dauer, A.; Schnetz, E.; Fartasch, M.; Hensel, A. High molecular compounds (polysaccharides and proanthocyanidins) from hamamelis virginiana bark: Influence on human skin keratinocyte proliferation and differentiation and influence on irritated skin. Phytochemistry 2001, 58, 949–958. [Google Scholar] [CrossRef]
- Dawid-Pać, R. Medicinal plants used in treatment of inflammatory skin diseases. Adv. Dermatol. Allergol. 2013, 30, 170. [Google Scholar] [CrossRef] [PubMed]
- Duckstein, S.M.; Stintzing, F.C. Investigation on the phenolic constituents in hamamelis virginiana leaves by hplc-dad and lc-ms/ms. Anal. Bioanal. Chem 2011, 401, 677–688. [Google Scholar] [CrossRef] [PubMed]
- Engels, C.; GAnzle, M.G.; Schieber, A. Fractionation of gallotannins from mango (mangifera indica l.) kernels by high-speed counter-current chromatography and determination of their antibacterial activity. J. Agric. Food Chem. 2010, 58, 775–780. [Google Scholar] [CrossRef] [PubMed]
- Duckstein, S.M.; Lorenz, P.; Stintzing, F.C. Conversion of phenolic constituents in aqueous hamamelis virginiana leaf extracts during fermentation. Phytochem. Anal. 2012, 23, 588–597. [Google Scholar] [CrossRef] [PubMed]
- Abu-Reidah, I.M.; Ali-Shtayeh, M.S.; Jamous, R.M.; Arráez-Román, D.; Segura-Carretero, A. Hplc-dad-esi-ms/ms screening of bioactive components from rhus coriaria l.(sumac) fruits. Food Chem. 2015, 166, 179–191. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.-S.; Yang, J.; Kim, M.-J. Alpha glucosidase inhibitory effect, anti-microbial activity and uplc analysis of rhus verniciflua under various extract conditions. J. Med. Plants Res. 2011, 5, 778–783. [Google Scholar]
- Regazzoni, L.; Arlandini, E.; Garzon, D.; Santagati, N.A.; Beretta, G.; Facino, R.M. A rapid profiling of gallotannins and flavonoids of the aqueous extract of rhus coriaria l. By flow injection analysis with high-resolution mass spectrometry assisted with database searching. J. Pharm. Biomed. Anal. 2013, 72, 202–207. [Google Scholar] [CrossRef] [PubMed]
- Salminen, J.-P.; Ossipov, V.; Loponen, J.; Haukioja, E.; Pihlaja, K. Characterisation of hydrolysable tannins from leaves of betula pubescens by high-performance liquid chromatography–mass spectrometry. J. Chromatogr. A 1999, 864, 283–291. [Google Scholar] [CrossRef]
- Wojtowicz, A.M.; Oliveira, S.; Carlson, M.W.; Zawadzka, A.; Rousseau, C.F.; Baksh, D. The importance of both fibroblasts and keratinocytes in a bilayered living cellular construct used in wound healing. Wound Repair Regen. 2014, 22, 246–255. [Google Scholar] [CrossRef] [PubMed]
- Budovsky, A.; Yarmolinsky, L.; Ben-Shabat, S. Effect of medicinal plants on wound healing. Wound Repair Regen. 2015, 23, 171–183. [Google Scholar] [CrossRef] [PubMed]
- Dos Santos, M.D.; Almeida, M.C.; Lopes, N.P.; De Souza, G.E.P. Evaluation of the anti-inflammatory, analgesic and antipyretic activities of the natural polyphenol chlorogenic acid. Biol. Pharm. Bull. 2006, 29, 2236–2240. [Google Scholar] [CrossRef] [PubMed]
- Fu, S.; Hui, C.; Li, L.; Cheuk, Y.; Qin, L.; Gao, J.; Chan, K.-M. Total flavones of hippophae rhamnoides promotes early restoration of ultimate stress of healing patellar tendon in a rat model. Med. Eng. Phys. 2005, 27, 313–321. [Google Scholar] [CrossRef] [PubMed]
- Gomathi, K.; Gopinath, D.; Ahmed, M.R.; Jayakumar, R. Quercetin incorporated collagen matrices for dermal wound healing processes in rat. Biomaterials 2003, 24, 2767–2772. [Google Scholar] [CrossRef]
- Chaudhari, M.; Mengi, S. Evaluation of phytoconstituents of terminalia arjuna for wound healing activity in rats. Phytother. Res. 2006, 20, 799–805. [Google Scholar] [CrossRef] [PubMed]
- Tulio Jr, A.Z.; Chang, C.; Edirisinghe, I.; White, K.D.; Jablonski, J.E.; Banaszewski, K.; Kangath, A.; Tadapaneni, R.K.; Burton-Freeman, B.; Jackson, L.S. Berry fruits modulated endothelial cell migration and angiogenesis via phosphoinositide-3 kinase/protein kinase b pathway in vitro in endothelial cells. J. Agric. Food Chem. 2012, 60, 5803–5812. [Google Scholar] [CrossRef] [PubMed]
- Mukherjee, P.K.; Maity, N.; Nema, N.K.; Sarkar, B.K. Bioactive compounds from natural resources against skin aging. Phytomedicine 2011, 19, 64–73. [Google Scholar] [CrossRef] [PubMed]
- De Sousa Araújo, T.A.; Alencar, N.L.; de Amorim, E.L.C.; de Albuquerque, U.P. A new approach to study medicinal plants with tannins and flavonoids contents from the local knowledge. J. Ethnopharmacol. 2008, 120, 72–80. [Google Scholar] [CrossRef] [PubMed]
- Shimoyama, A.T.; Santin, J.R.; Machado, I.D.; e Silva, A.M.D.O.; de Melo, I.L.P.; Mancini-Filho, J.; Farsky, S.H. Antiulcerogenic activity of chlorogenic acid in different models of gastric ulcer. N.-S. Arch. Pharmacol. 2013, 386, 5–14. [Google Scholar] [CrossRef] [PubMed]
- Yun, N.; Kang, J.-W.; Lee, S.-M. Protective effects of chlorogenic acid against ischemia/reperfusion injury in rat liver: Molecular evidence of its antioxidant and anti-inflammatory properties. J. Nutr. Biochem. 2012, 23, 1249–1255. [Google Scholar] [CrossRef] [PubMed]
- Dos Santos, J.S.; Monte-Alto-Costa, A. Caffeic acid phenethyl ester improves burn healing in rats through anti-inflammatory and antioxidant effects. J. Burn Care Res. 2013, 34, 682–688. [Google Scholar] [CrossRef] [PubMed]
- Suzuki, A.; Yamamoto, N.; Jokura, H.; Yamamoto, M.; Fujii, A.; Tokimitsu, I.; Saito, I. Chlorogenic acid attenuates hypertension and improves endothelial function in spontaneously hypertensive rats. J. Hypertens. 2006, 24, 1065–1073. [Google Scholar] [CrossRef] [PubMed]
- Wang, B.; Zhong, Y.; Gao, C.; Li, J. Myricetin ameliorates scopolamine-induced memory impairment in mice via inhibiting acetylcholinesterase and down-regulating brain iron. Biochem. Biophysic. Res. Commun. 2017, 490, 336–342. [Google Scholar] [CrossRef] [PubMed]
- Silva, L.; Da Hora, G.; Soares, T.; Bojer, M.; Ingmer, H.; Macedo, A.; Trentin, D. Myricetin protects galleria mellonella against staphylococcus aureus infection and inhibits multiple virulence factors. Sci. Rep. 2017, 7, 2823. [Google Scholar] [CrossRef] [PubMed]
- Huang, J.-H.; Huang, C.-C.; Fang, J.-Y.; Yang, C.; Chan, C.-M.; Wu, N.-L.; Kang, S.-W.; Hung, C.-F. Protective effects of myricetin against ultraviolet-b-induced damage in human keratinocytes. Toxicology. 2010, 24, 21–28. [Google Scholar] [CrossRef] [PubMed]
- Kang, K.A.; Wang, Z.H.; Zhang, R.; Piao, M.J.; Kim, K.C.; Kang, S.S.; Kim, Y.W.; Lee, J.; Park, D.; Hyun, J.W. Myricetin protects cells against oxidative stress-induced apoptosis via regulation of pi3k/akt and mapk signaling pathways. Int. J. Mol. Sci. 2010, 11, 4348–4360. [Google Scholar] [CrossRef] [PubMed]
- Bagdas, D.; Etoz, B.C.; Gul, Z.; Ziyanok, S.; Inan, S.; Turacozen, O.; Gul, N.Y.; Topal, A.; Cinkilic, N.; Tas, S.; et al. In vivo systemic chlorogenic acid therapy under diabetic conditions: Wound healing effects and cytotoxicity/genotoxicity profile. Food Chem. Toxicol. 2015, 81, 54–61. [Google Scholar] [CrossRef] [PubMed]
- Bagdas, D.; Gul, N.Y.; Topal, A.; Tas, S.; Ozyigit, M.O.; Cinkilic, N.; Gul, Z.; Etoz, B.C.; Ziyanok, S.; Inan, S.; et al. Pharmacologic overview of systemic chlorogenic acid therapy on experimental wound healing. Naunyn Schmiedebergs Arch. Pharmacol. 2014, 387, 1101–1116. [Google Scholar] [CrossRef] [PubMed]
- Jin, U.H.; Lee, J.Y.; Kang, S.K.; Kim, J.K.; Park, W.H.; Kim, J.G.; Moon, S.K.; Kim, C.H. A phenolic compound, 5-caffeoylquinic acid (chlorogenic acid), is a new type and strong matrix metalloproteinase-9 inhibitor: Isolation and identification from methanol extract of euonymus alatus. Life Sci 2005, 77, 2760–2769. [Google Scholar] [CrossRef] [PubMed]
- DeCicco-Skinner, K.L.; Henry, G.H.; Cataisson, C.; Tabib, T.; Gwilliam, J.C.; Watson, N.J.; Bullwinkle, E.M.; Falkenburg, L.; O’Neill, R.C.; Morin, A. Endothelial cell tube formation assay for the in vitro study of angiogenesis. J. Vis. Exp. 2014, e51312. [Google Scholar] [CrossRef] [PubMed]
Sample Availability: Samples of the compounds are available from the authors. |

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Peak | tR (min) | Compound | UV-Vis λmax (nm) | m/z [M−H]− | m/z [M−2H]−2 | HPLC-TOF-MS(negative) | Identification Method |
|---|---|---|---|---|---|---|---|
| 1 | 3.0 | Galloyl glucose | 231, 279 | 331.0657 | MS-UV,NMR | ||
| 2 | 3.5 | Astringenin | 405.1172 | MS-UV | |||
| 3 | 4.5 | 3-O-Caffeoylquinic acid | 325 | 353.0861 | 707.1710 | MS-UV,NMR | |
| 4 | 5.0 | Trigalloyl glucose | 275 | 635.0850 | 465.0706 | MS-UV | |
| 5 | 6.2 | 5-O Caffeoylshikimic acid | 288, 320 | 335.0781 | MS-UV,NMR | ||
| 6 | 6.5 | Myricetin-3-O-β-glucoside | 261, 357 | 479.0836 | 316.0206 | MS-UV,NMR | |
| 7 | 8.0 | 1,2,4,6-Tetragalloyl glucose | 275, 220 | 787.0996 | 617.0808 | MS-UV,NMR | |
| 8 | 9.3 | Myricetin-3-O-β-rhamnoside | 260, 352 | 463.0887 | 316.022 | MS-UV,NMR | |
| 9 | 10.4 | Quercetin 3-O-β-glucoside | 266, 356 | 463.0896 | MS-UV,NMR | ||
| 10 | 11.3 | Pentagalloyl glucose | 211, 278 | 939.1110 | 769.0952 | MS-UV,NMR | |
| 11 | 12.0 | Kaempherol-3-O-rutinoside | 593.1525 | 463.0879 | MS-UV,NMR | ||
| 12 | 12.8 | Digallic acid | 321.0258 | MS-UV | |||
| 13 | 13.5 | Quercetin-3-O-β-rhamnoside | 258, 352 | 447.0935 | 300.0237 | MS-UV,NMR | |
| 14 | 13.5 | Kaempferol-3-O-β-glucoside | 447.0935 | MS-UV,NMR | |||
| 15 | 14.7 | Hexagalloyl glucose | 214, 278 | 1091.1195 | 939.1100, 769.0865, 637.0700 | MS-UV | |
| 16 | 16.3 | Heptagalloyl glucose | 214, 275 | 1243.1278 | 621.0412 | 1091.1181, 939,1102, 769.0859 | MS-UV |
| 17 | 17.1 | Kaempferol-3-O-β-rhamnoside | 431.0960 | MS-UV,NMR | |||
| 18 | 18.4 | Octagalloyl glucose | 214, 276 | 1395.1373 | 697.0706 | 1243.1265, 1091.1175, 939.1093, 767.1083 | MS-UV |
| 19 | 20.4 | Nonagalloyl glucose | 214, 276 | 1547.1480 | 773.0742 | 1395.1372, 1243.1268, 1091.1167, 919.1177 | MS-UV |
| 20 | 21.0 | Quercetin-3-(3,4,6 trigalloyl glucose) | 919.1236 | 767.1121, 615.1005, 463.0889 | MS-UV,NMR | ||
| 21 | 22.1 | Decagalloyl glucose | 214, 276 | 1699.1581 | 849.0767 | 1547.1470, 1395.134, 1243.1254, 1091.1146, 939.1083 | MS-UV |
| 22 | 23.7 | Undecagalloyl glucose | 214, 273, 305 | 925.0841 | 849.0778, 773.0715, 697.0648, 621.0590, 545.0549, 469.0500 | MS-UV | |
| 23 | 25.2 | Dodecagalloyl glucose | 214, 273, 305 | 1001.5901 | 925.0841, 849.0781, 773.0721, 697.654, 621.0580, 545.0556, 469.0498 | MS-UV | |
| 24 | 26.8 | Tridecagalloyl glucose | 214, 273, 305 | 1077.5907 | 1001.5907, 925.0847, 849.0779, 773.0731, 697.0649, 621.0598, 545.0554, 469.0506 | MS-UV | |
| 25 | 27.8 | Tetradecagalloyl glucose | 214, 273, 305 | 1153.6028 | 1077.5960, 1001.5898, 925.0832, 849.0781, 773.0710, 697.0658, 621.0593, 545.0556, 469.0505 | MS-UV |
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Moghadam, S.E.; Ebrahimi, S.N.; Salehi, P.; Moridi Farimani, M.; Hamburger, M.; Jabbarzadeh, E. Wound Healing Potential of Chlorogenic Acid and Myricetin-3-O-β-Rhamnoside Isolated from Parrotia persica. Molecules 2017, 22, 1501. https://doi.org/10.3390/molecules22091501
Moghadam SE, Ebrahimi SN, Salehi P, Moridi Farimani M, Hamburger M, Jabbarzadeh E. Wound Healing Potential of Chlorogenic Acid and Myricetin-3-O-β-Rhamnoside Isolated from Parrotia persica. Molecules. 2017; 22(9):1501. https://doi.org/10.3390/molecules22091501
Chicago/Turabian StyleMoghadam, Sara E., Samad N. Ebrahimi, Peyman Salehi, Mahdi Moridi Farimani, Matthias Hamburger, and Ehsan Jabbarzadeh. 2017. "Wound Healing Potential of Chlorogenic Acid and Myricetin-3-O-β-Rhamnoside Isolated from Parrotia persica" Molecules 22, no. 9: 1501. https://doi.org/10.3390/molecules22091501