
Protein Interaction and Na/K-ATPase-Mediated Signal Transduction


Marshall Institute for Interdisciplinary Research, Marshall University, Huntington, WV 25703, USA
*
Author to whom correspondence should be addressed.
Molecules 2017, 22(6), 990; https://doi.org/10.3390/molecules22060990
Submission received: 7 April 2017
/
Revised: 1 June 2017
/
Accepted: 2 June 2017
/
Published: 14 June 2017
(This article belongs to the Special Issue Cardiotonic Steroids)
{kind=link}
{kind=link}
{kind=link}
Abstract
:The Na/K-ATPase (NKA), or Na pump, is a member of the P-type ATPase superfamily. In addition to pumping ions across cell membrane, it is engaged in assembly of multiple protein complexes in the plasma membrane. This assembly allows NKA to perform many non-pumping functions including signal transduction that are important for animal physiology and disease progression. This article will focus on the role of protein interaction in NKA-mediated signal transduction, and its potential utility as target for developing new therapeutics.
1. Introduction
The Na/K-ATPase (NKA) was discovered by Skou 60 years ago as the molecular machine for pumping Na+ and K+ across cell membrane [1]. In the early 1970s, several studies revealed the regulatory effects of ouabain on cell growth and gene expression. At that time, these regulatory effects of ouabain were all ascribed to the pump inhibition and the resulted change in intracellular ion concentration [2,3,4]. About 20 years ago, a series of studies conducted first in neonatal cardiac myocytes and subsequently in renal epithelial cells, showed that ouabain could activate a number of cell growth-related pathways, of which many are independent of changes in intracellular ion concentration. These studies have led to a great effort by many laboratories and subsequent demonstration that the NKA actually has many non-pumping functions [5,6]. In this review, we will first look back at our evolved view of NKA in cell biology. We will then give an in-depth discussion of NKA-mediated signal transduction; its role in animal physiology and disease progression; theoretical consideration and experimental evidence of direct protein interactions as the molecular mechanism; and the possibility of targeting such interactions for developing new therapeutics.
2. Na/K-ATPase and Active Ion Transport
NKA belongs to the P-type ATPase family. Before Skou discovered NKA in 1957, cell biologists had speculated the existence of such transmembrane machinery for over 100 years. One of the most important early studies was conducted by Carl Schmidt who demonstrated the existence of a Na+/K+ concentration gradient across cell membrane [7]. This led to the proposal by Rudolf Heidenhain of a “microscopic steamship” laying within the membrane that is capable of maintaining this gradient [8]. Subsequently, several key discoveries paved the way and convinced cell biologists of a principle responsible for transmembrane movement of ions against their concentration gradients. Most notably were the studies by Ernest Overton, showing that muscle cells had active transport mechanism allowing cells to move Na+ and K+ across cell membrane via the consumption of energy [9,10]. This was confirmed by Heppel and Steinbach in muscle cells using isotopes [11,12,13] and by several groups of U.S. scientists in red blood cells [14,15,16]. Finally, cardiac glycosides were found to be specific inhibitors of such active transport in red blood cells [17], and the requirement of ATP for K+ uptake in these cells further supported and linked the transport system to membrane-bound ATPase sensitive to cardiac glycosides [18,19].
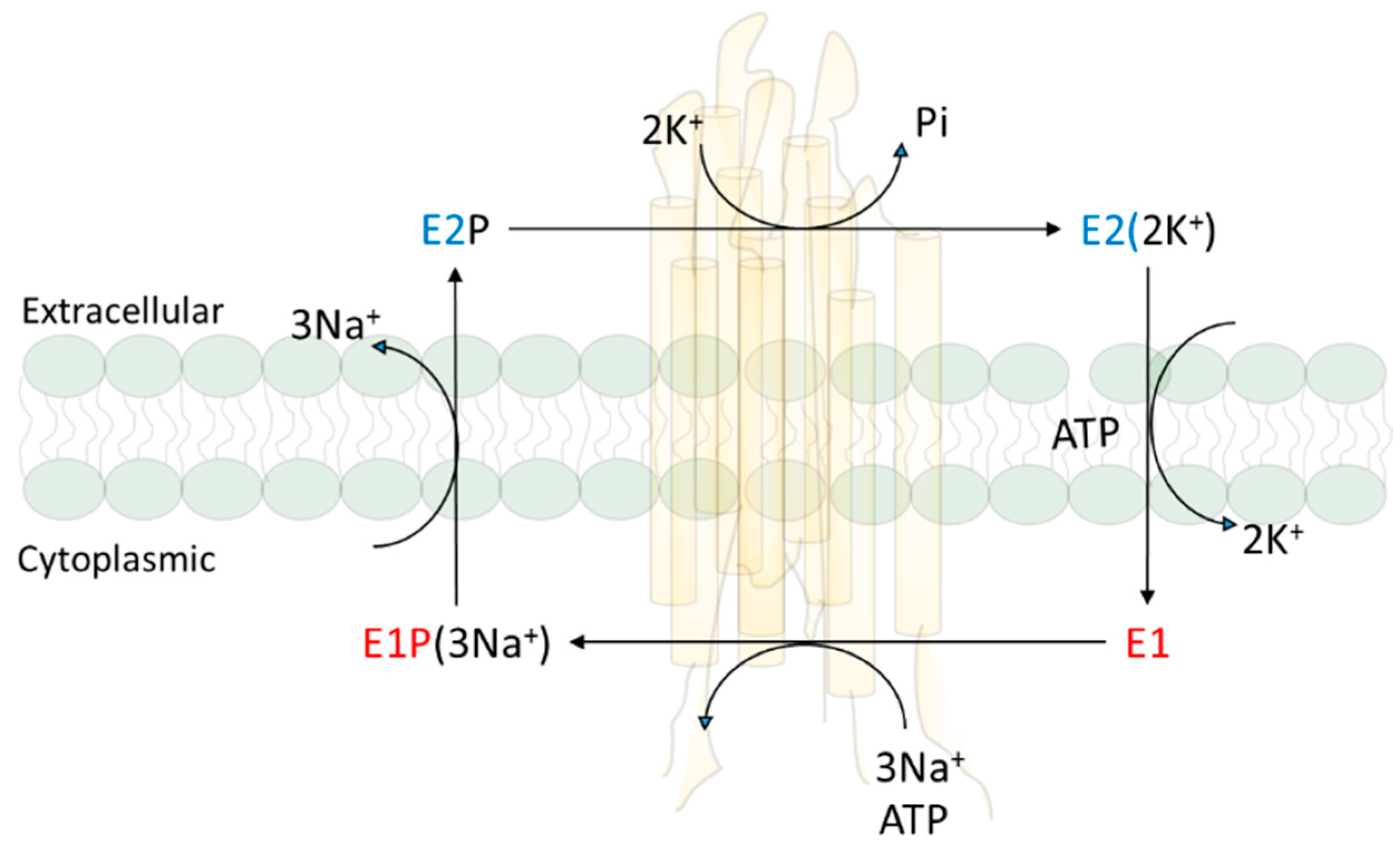
At the time Skou discovered NKA, Robert Post had found that the ATPase is responsible for the active transport of three Na+ and two K+ across the plasma membrane in red blood cells. His subsequent work on the reaction mechanism led to the Albers–Post scheme that is not only true to the NKA, but also applies to other members of P-type ATPase family [20,21,22]. Ion pumping is linked to the cycle of conformational changes. Around the same time, cell biologists and renal physiologists developed a kidney NKA purification protocol, and generated a large number of important mechanistic and cell biological data that refine the structure, reaction mechanism, and cellular regulation of NKA [23,24,25,26]. Importantly, we understand that NKA exists in a dynamic state of conformation equilibrium which was important for its ability to convert ATP hydrolysis to the binding and movement of ions across the plasma membrane as illustrated in Albers–Post reaction mechanism scheme (Figure 1). It also allows the binding of many ligands (chemicals such as cardiotonic steroids that can bind to NKA with high affinity) to the NKA in a conformational state-dependent manner.
NKA, as a large and highly expressed membrane protein complex (most cells contain over one million surface pumps per cell), consists of two noncovalently linked subunits, α and β [27,28]. The α subunit contains ATP and other ligand binding sites, and is considered as the catalytic subunit. The scaffolding function of β subunit is essential for the membrane targeting and full function of the NKA. Four isoforms of NKA have been identified. The existence of different isoforms was first suggested by Michael Marks and Nicholas Seeds in 1987 [29]. They found that ouabain exhibited two distinct inhibition phases of NKA in preparations made from the mouse brain. Subsequently, Sweadner identified at least two isoforms of NKA in membrane preparations from rat brain [30]. The further breakthrough came from the molecular cloning of NKA, first from sheep kidney in 1985 [31], and then the identification of four isoforms from different rat tissues [31,32,33]. Furthermore, studies showed that different isoforms are expressed in a tissue-specific manner [32,33]. The α1 isoform is found in all cells and is prevalent in all epithelial cells. The α2 and α3 isoforms are expressed in skeletal muscle, neuronal tissue, and cardiac myocytes. The α4 isoform is expressed in the testis and regulates sperm motility [27,34]. The sequence identity is about 87% among α1, α2 and α3, while α1 and α4 are 78% identical. Nevertheless, the overall tertiary structure appears to be identical among all isoforms [35].
3D structures of several P-type ATPases including α1 NKA have been resolved [36,37,38,39]. The overall structure of NKA is composed of ten transmembrane helices important for ion binding, occlusion and movement, and three cytosolic domains called the N-domain (nucleotide binding), P-domain (phosphorylation) and A-domain (actuator) that confer ATP hydrolyzing activity. Overall, crystal structures are in agreement with the deduced structures from biochemical studies of the past 60 years. Interestingly, recent resolution of several different CTS-bound NKAs also reveals that although these compounds all inhibit ATPase activity they could actually produce different structure perturbations [40,41].
In short, a lot has been learnt about the structure and function of NKA as an ion pump over the last 60 years. Moreover, we have gradually recognized that the NKA may be engaged in dynamic interaction with other membrane and cytosolic proteins because of the dynamic nature of NKA conformational equilibrium and the large number of NKA in the plasma membrane. Such interactions play at least three different roles in cell biology: (1) dynamic regulation of ionic concentrations including Na+, K+ and consequently Ca2+ by regulating the pumping activity of NKA, the focus of early years of investigation; (2) a key player in cellular signal transduction because of its direct interactions with signaling proteins; and (3) as a signal integrator by organizing specific membrane microdomains and by bridging different affecters and effectors together through its scaffolding function.
3. Na/K-ATPase and Signal Transduction
Cardiotonic Steroids (CTS) include plant-derived digitalis such as digoxin and ouabain, and vertebrate-derived aglycones such as bufalin and marinobufagenin (MBG) [42,43]. Digoxin has been used to manage congestive heart failure for over 200 years, and bufalin/MBG are active components in traditional Chinese medicine “Chan Su”. However, the digitalis-specific inhibition of NKA was not recognized until the discovery of NKA in the 1950s [1,17]. In cardiac myocytes, inhibition of NKA activity by CTS increases intracellular Na+ concentration, which leads to the accumulation of intracellular Ca2+ through functional coupling to the Na+/Ca2+ exchanger (NCX). Consequently, an increase in Ca2+ concentration enhances the contractility of cardiac muscle and causes positive inotropy [42]. In patients with heart/kidney diseases, an increase in endogenous CTS has also been observed [44,45,46].
3.1. Na/K-ATPase: More Than a Pump
In addition to its effect on cardiac contraction, CTS were recognized long time ago to play a role in cell growth regulation. In the 1970s, for instance, ouabain at low nM concentrations was found to regulate gene expression and the mitogen-induced differentiation and proliferation in lymphoblasts [3,47,48]. Taking into the consideration that IC50 of ouabain is around 50–100 nM for human α1 NKA [49,50,51], such low concentration of ouabain on cell growth is unlikely due to the substantial inhibition of transmembrane movement of ions via the NKA, suggesting additional mechanism being responsible for CTS-induced changes in cell growth. Other than cell growth regulation, picomolar concentrations of MBG can stimulate the synthesis of collagen in human dermal fibroblasts [52].
A series of studies from our laboratory published in the late 1990s and early 2000s revealed that CTS could stimulate protein tyrosine phosphorylation and a number of growth-related pathways in cell type- and tissue type-dependent manner [53,54,55,56,57], which has now been largely confirmed by studies from other laboratories around the world [49,58,59,60,61,62,63,64,65,66,67,68,69,70,71,72,73,74,75]. These new findings suggest NKA as an important signal transducer, and the involvement of protein kinase cascades in the cell growth regulation by CTS rather than the inhibition of ATPase activity. At the time, the importance of NKA-mediated signal transduction had not been fully appreciated and was considered as being “moonlighting”. This is in part because of the following two unresolved albeit important issues. First, since NKA has both pumping and signaling functions, it was difficult to study signaling independent of pumping, especially in cardiac myocytes where NKA is tightly coupled to other membrane transporters such as NCX [42,76,77,78,79,80]. Second, the realization that NKA has no tyrosine kinase or phosphatase activity had raised question as to how binding of ouabain to NKA stimulated protein tyrosine phosphorylation, which was required for the regulatory actions of ouabain on cell growth. Although direct protein interaction was speculated at the time, no experimental evidence, especially the involvement of tyrosine kinase/phosphatase, had been reported in the literature. These issues have driven the next ten years of investigations, and led to our current appreciation of the molecular basis of NKA-mediated signal transduction in cells.
3.2. Protein Interaction in Signal Transduction
It is well established that regulated protein interaction is a key to cellular signal transduction. The best studied examples are those of G protein-coupled receptors and receptor tyrosine kinases (RTKs). Extracellular ligand binding to RTKs stabilizes receptor dimerization and causes trans-phosphorylation of the receptor. The phosphorylated tyrosine residues provide binding sites for Src homology 2 (SH2) domain- and phosphotyrosine binding domain-containing proteins, and propagate the downstream signaling [81,82].
Interestingly, early studies identified several NKA-interacting proteins such as ankyrin, adducin and FXYD family of proteins [83,84,85]. However, most of these as well as some of the recent studies have focused on the role of such interactions in the regulation of NKA activity and trafficking. For example, FXYD family proteins are expressed in a tissue-specific manner and appear to act as a third subunit of the enzyme [86]. Although it is not required for functional expression of α/β NKA, FXYD proteins interact, and regulate NKA pumping activity [36,87,88,89]. Interestingly, some interactions between FXYDs and NKA are regulated by membrane receptors. For example, FXYD1, also known as phospholemman (PLM), is a principal phosphorylation substrate of c-AMP dependent protein kinase A and of Ca2+-phospholipid-dependent protein kinase C at Ser68 (PKA), or at Ser63, Ser68 and Thr69 (PKC) [90,91,92]. Unphosphorylated FXYD1 inhibits NKA through the direct protein interaction [93,94,95]. In addition, a number of signaling proteins has also been identified during the studies of hormonal regulation of NKA trafficking in kidney epithelial cells. For example, dopamine stimulates the recruitment of arrestin, spinophilin, GPCR kinase and 14-3-3ε, to the α1 NKA. In another example, the association of 14-3-3ζ to the α1 subunit facilitates the binding of PI3K to the α1 subunit that subsequently leads to the endocytosis of the NKA [96,97]. Conversely, in response to angiotensin II, adaptor protein-1 attaches to the α1 subunit and facilitates the recruitment of the NKA to the plasma membrane [98]. Functionally, Bcl-2 proteins were also reported directly interacting with NKA [99]. The interactions are critical for control of cell survival and apoptosis. The ratio of pro-survival and pro-apoptotic proteins interacting with NKA may determine NKA function.
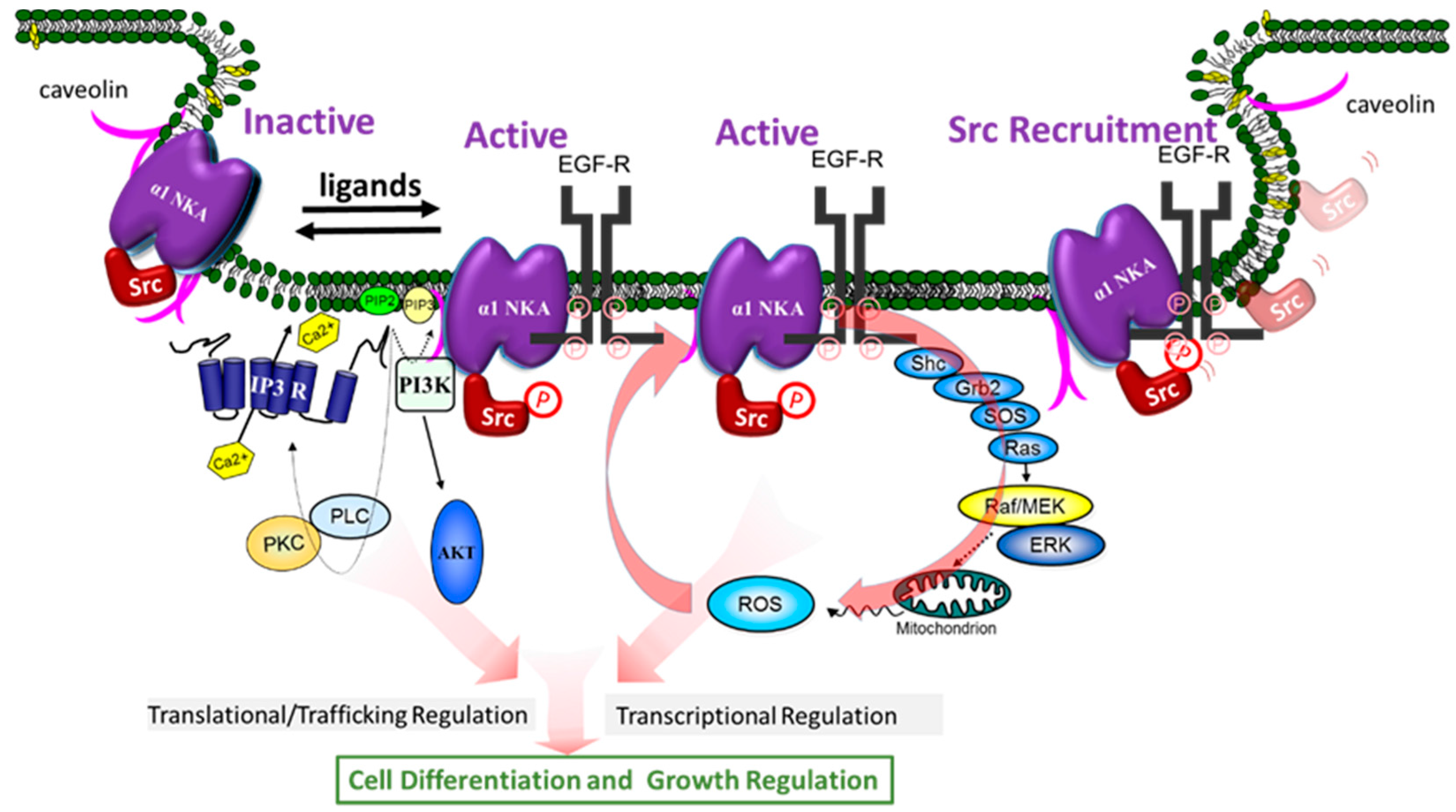
In view of our demonstration that NKA plays a role in signal transduction, the fact that protein interaction is a key to signal transduction has prompted us to ask if NKA is capable of regulating the function of its interacting proteins, and, if so, whether NKA ligands can regulate such interactions, and consequently activate cellular signaling events. The concerted efforts of many over the last ten years have yielded solid evidence that supports this hypothesis. Specifically, these studies have led to the discovery that α1 NKA/Src complex is an important receptor for CTS and other NKA ligands to activate protein/lipid kinase cascades, to generate ROS, and to stimulate Ca2+ oscillation in a cell-specific manner (Figure 2). Moreover, new findings have suggested that α1 NKA may regulate such interactions in a conformation-dependent manner. We further speculate that the receptor NKA can actually adapt both active and inactive conformations, and that NKA ligands may stabilize either active (agonists) or inactive (inverse agonists) conformation to exert their regulatory effects on cells (Figure 2). Finally, recent studies have also revealed NKA as a potential signal integrator important for assembling cellular signalosomes and for effective coupling of affecters and effectors.
3.3. Src Kinase in NKA-Mediated Signal Transduction
The first clue that Src kinase is important for NKA-mediated signal transduction was from the studies by Haas in early 2000 [56]. Src family kinases are membrane-associated non-receptor tyrosine kinases, and they play an essential role in the signal transduction pathways provoked by many extracellular stimuli such as growth factors, and ligands of G protein-coupled receptors [100]. We and others have shown that α1 NKA regulates Src activity through a conformation-dependent interaction. It also plays an important role in Src targeting through a phosphorylation-dependent mechanism. Binding of CTS to this NKA/Src receptor complex leads to the activation of the associated Src, recruitment of additional Src, and the initiation of the signal transduction processes (Figure 2) [101].
3.4. Evidence of NKA/Src Interaction
The following evidence supports the hypothesis that NKA and Src form a functional receptor. First, ouabain and other CTS stimulated protein tyrosine phosphorylation in many different types of cells including cardiac myocytes, smooth muscle and renal epithelial cells to name a few [54,56,59,102,103,104,105,106]. Second, upon ouabain stimulation, Src activation, as evidenced by its translocation from cytosolic fraction to a Triton-insoluble fraction, and an increase in Y418 phosphorylation (pY418) (but not a decrease in Y529 phosphorylation), was one of the earliest events [56]. Moreover, Src inhibitors blocked the ouabain-induced tyrosine phosphorylation and ouabain-activated downstream signal pathways such as ERK. The CTS-induced cell growth effect could also be sufficiently attenuated by Src kinase inhibitors [56,103]. Third, genetic evidence also supports the requirement of Src in ouabain-induced signal transduction because ouabain failed to increase protein tyrosine phosphorylation in SYF cells where Src family kinases are knocked out. On the other hand, rescuing these cells with Src fully restored ouabain-induced signal transduction [107]. Fourth, NKA and Src were co-enriched in caveolar fractions in many cell types [107]. While immunofluorescence imaging analyses confirmed the co-localization of these two proteins, FRET analyses suggested a direct interaction between these two proteins [101]. Further evidence of direct interaction came from co-immunoprecipitation experiments, first reported by Haas et al. [56], then confirmed by many others [59,103,104,105,108,109,110]. However, Kaplan et al. reported the failure of co-immunoprecipitation of NKA with Src in breast cancer cells and questioned whether NKA interacts with Src [111]. Although it remains to be determined, it is important to note that Kaplan lab conducted the immunoprecipitation using a different anti-α1 antibody from other labs. Interestingly, the polyclonal antibody used by Kaplan lab was raised against the fragment of α1 NKA where the putative Src binding site resides, which ironically could provide further support of direct interaction. In addition to immunoprecipitation analyses, GST-fused fragments of intracellular domains of α1 NKA also pulled-down Src from cell lysates, indicating the existence of a direct and specific interaction between these two proteins [101]. Finally, many have demonstrated that the interaction between these two proteins was actually regulated by CTS [56,59,101,103,104,105,108,109,110]. For example, ouabain increased the co-immunoprecipitation of these two proteins, and this increase was sensitive to Src inhibitors [56].
3.5. The Identification of Putative Src Binding Sites and the Discovery of NaKtide as a Specific Inhibitor of NKA-Mediated Signal Transduction
The direct interaction between α1 NKA and Src was further demonstrated by co-precipitation studies using purified dog/pig kidney NKA and fully active but unphosphorylated Src [101]. Functionally, this interaction prevented Src Y418 phosphorylation that is required for full activation of Src kinase activity. Using GST pull-down analyses of different cytosolic domains of α1 NKA and functional domains of Src, two putative Src binding sites have been mapped. One is between second cytosolic domain of α1 subunit of NKA and Src SH2 domain. The other one locates in the N domain of α1 that interacts with Src kinase domain. The latter interaction inhibits Y418 phosphorylation. Again, the polyclonal antibody used in the co-immunoprecipitation from Kaplan Lab was directed to this large fragment of α1 NKA. Importantly, ouabain was shown to release the interaction between purified α1 NKA and Src kinase domain, without affecting the binding of SH2 [101,112]. Further mapping of this interaction has identified the 20 amino acid NaKtide sequence in the N domain of α1 NKA being responsible for the direct interaction between α1 and Src kinase domain [112]. The synthesized NaKtide mimics NKA, capable of interacting with and inhibiting Src. Moreover, when NaKtide is made into pNaKtide by adding TAT (13 amino acids) sequence to the N-terminus, it becomes cell permeable. Functional studies demonstrate that pNaKtide also mimics NKA, and is effective in inhibiting NKA-interacting pool of Src in an ATP concentration-independent manner. Consequently, it blocks ouabain-induced activation of Src, ERK and hypertrophic growth in cardiac myocytes [112]. It also specifically suppresses cell proliferation in cancers, whose Src activity is not efficiently inhibited by α1 NKA [113].
3.6. Identification of CD2 as an Important Src SH2 Ligand
Although the interaction between Src SH2 domain and NKA α1 is not reduced upon ouabain binding, it is important in the recruitment and targeting of Src. In LLC-PK1 cells transfected with α1 CD2, exogenously expressed CD2 competitively bound to Src kinase in cells and prevented Src from being targeted to various effectors. Therefore, as the exogenous Src SH2 ligand, α1 CD2 increased the global Src activity but blocked Src-mediated pathways including ouabain-induced signal transduction [114]. In short, we and others have generated strong evidence of α1 NKA/Src interaction and demonstrated the importance of such interaction in CTS-induced signal transduction. However, it is important to note that Karlish and his colleagues have questioned such interaction based on their work with purified recombinant human α1 NKA expressed in yeast and Src kinase expressed in bacteria [115]. A major difference is noted between the study from Karlish and our investigation. We used non-phosphorylated Src in our studies whereas Karlish used bacteria-expressed Src that is known to be phosphorylated at both Y418 and Y529 sites. Phosphorylation of these sites affects the activity and binding of Src to other proteins [116]. For example, it is known that SH2 plays an important role in directing the interaction between Src and its partners [117]. This is also true for NKA/Src interaction as we reported recently [114]. The heterogeneous nature of bacteria-expressed Src in both Y418 and Y529 phosphorylation makes it difficult to measure the potential interaction involving the SH2 domain, especially if the interaction has low affinity. To this end, we and others have demonstrated that inhibition of Src by PP2 could abolish ouabain-induced increases in Src binding, reaffirming an important role of Src-mediated phosphorylation and SH2-directed interaction.
3.7. The Identification of a Mutant α1 NKA That Pumps but Is Null in Src Interaction
To further verify that NaKtide sequence is engaged in direct interaction with Src, we performed mutagenesis studies [118]. These studies indicated that the N-terminal helical structure of NaKtide is important for the Src binding and inhibition. Moreover, W423, L424, and R427 appear to be in direct contact with Src kinase domain. This conclusion was further supported by mutations (e.g., A420P and A425P) that disrupt the formation of helical structure. Significantly, when A420P or A425P was introduced into full length α1 NKA, we found that the mutated NKA retained full pumping capacity but failed in Src interaction and regulation [118]. Now, we have two mutant α1 NKA that work as a pump but not a signaling receptor.
3.8. NKA/Src Interactions Are Isoform-Specific
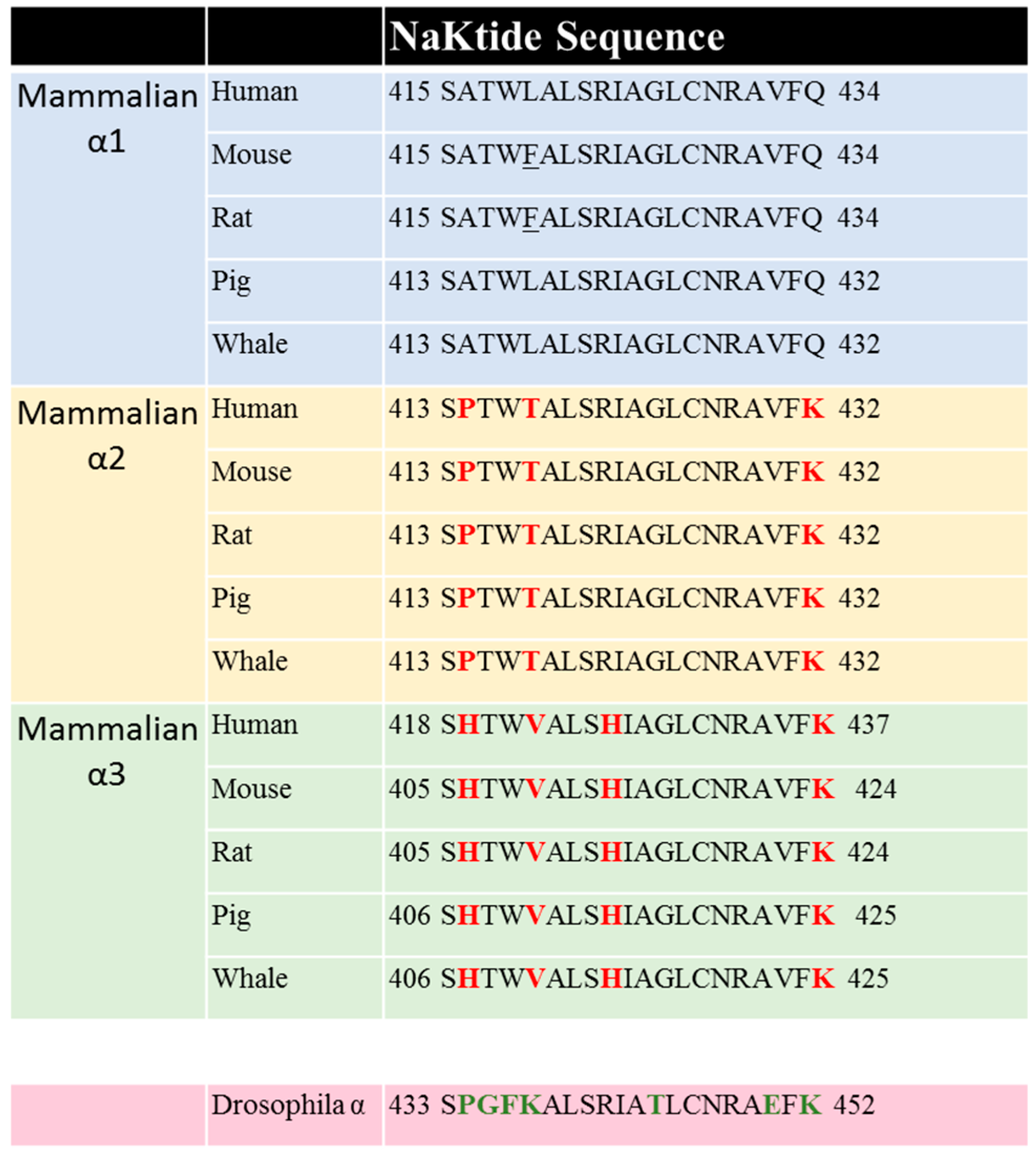
Sequence comparison shows that the NaKtide sequence is highly conserved in mammalian α1 NKA. However, the corresponding NaKtide sequences in α2 and α3 isoforms are different from that in α1. Interestingly, these differences are also conserved in both α2 and α3 sequences (Figure 3). In view of the importance of NaKtide sequence in α1 NKA-mediated Src interaction, we generated α2 and α3-expressing mammalian cells using a knock-down and rescue protocol, and demonstrated that both α2 and α3 NKA isoforms lack Src-interacting capacity [119,120]. As such, they do not carry Src-dependent signal transduction upon ouabain binding. However, α3 isoform differs from α2 because it does signal in a Src-independent manner [119]. These new findings reveal a major functional difference among three NKA isoforms, and re-enforces the importance of Src binding capacity of α1 NKA in the regulation of cell signal transduction. In addition, the observed difference in their ability to conduct signal transduction among three isoforms provide strong evidence that CTS-induced activation of protein kinases is unlikely due to changes in intracellular ATP secondary to the inhibition of the pumping as advocated by some [121,122]. Finally, Lingrel lab and others have generated strong evidence that different NKA isoforms do exert distinct regulation of animal physiology (e.g., muscle contraction). Therefore, our new findings also suggest the importance of the lack of Src binding for α2 and α3 specific cellular singling functions.
3.9. NKA/Src Complex as a Receptor
Mechanistic investigations of last fifteen years have revealed a novel molecular mechanism of NKA-mediated signal transduction. As illustrated in Figure 2, the receptor NKA interacts with several proteins to perform cell-specific signal transductions including Raf/MEK/ERK, PLC/PKC, PI3K/Akt, and Ca2+ signaling and the generation of ROS. One of the most important signaling partners trans-activated by NKA/Src receptor complex is EGF receptor, which is recruited and phosphorylated at several phosphorylation sites other than its major phosphorylation site Y1173 when cells are exposed to CTS [56]. The activated EGF receptor then recruits the adaptor protein Shc, which in turn binds the protein complex Grb2 and SOS. SOS is a guanine nucleotide exchange factor that activates Ras by exchanging GDP for GTP. Activated Ras then stimulates Raf/MEK and p42/44 ERK cascade [56,57]. Activation of this cascade by CTS appears to occur in most of cell types [57,59,104,109,123]. It is of interest to note that activated EGFR is also a critical element in the signal transduction networks of cytokines, H2O2, and pathways utilizing G protein-coupled receptors (51). However, the number of membrane α1 NKA in most of cells is at least over 100 times of G protein-coupled receptors. Thus, it is reasonable to speculate that α1 NKA may regulate the Src-dependent signaling pathways of G protein-coupled receptors. Moreover, in view of a critical role of EGFR in cancer, it would be of great importance to further dissect α1 NKA-mediated regulation of EGFR and its potential role in cancer biology.
Several important features of this newly appreciated signaling mechanism are worthy of further discussion. First, the activation of protein and lipid kinase cascades and the generation of second messengers ensure the formation of a positive feed forward loop that could amplify CTS-provoked signal transduction, and also allow signal diversification, transcriptional and translational regulation of gene expression [52,53,57]. This is best exemplified by the recruitment of additional signaling partners into the receptor complex [57,107,124], and by the ROS-induced signal amplification (Figure 2) [55,125,126]. In accordance, it explains how endogenous CTS could exert profound physiological effects at concentrations well below 1/100th of IC50 [127]. For example, it has been reported that ouabain at 10 to 100 nM was sufficient to stimulate mouse or rat cardiac fibroblasts, resulting in increased collagen production [52,128]. Similarly, such low concentrations of CTS were found to elicit Ca2+ oscillation in both mouse and rat kidney epithelial cells where only the ouabain-resistant α1 NKA is expressed [129]. It is also important to point out that the fetal bovine serum we all use in our cell culture may contain sufficient amount of CTS to promote cell growth [130]. Finally, signal amplification similar to these in rodents has also been observed in human cells [52].
Second, because NKA contains a large number of motifs both intracellularly and extracellularly, it would not be a surprise that NKA could perform much more regulatory function than those outlined in the scheme (Figure 2). Moreover, it is likely that many of these pathways could cross-talk to each other and exert cell-specific regulation depending on the context of available signaling constitutes. This is exemplified by the fact that NKA could regulate PI3K signaling in Src-knock out cells, whereas inhibition of Src also attenuates ouabain-induced PI3K signaling in normal cells [131,132,133,134]. Similarly, Src is also involved in ouabain-induced Ca2+ oscillation by affecting the interaction between IP3 receptor and α1 NKA in renal epithelial cells [129].
Third, this scheme provides a framework to begin addressing the role of NKA-mediated signal transduction in animal physiology. To this end, recent animal studies have demonstrated the importance of this signaling mechanism in wild array of physiological processes including renal salt handling, vascular activity, cardiac growth, and embryonic development to name a few [106,135,136,137,138,139,140,141,142,143]. These new findings call for the need of re-examination of CTS physiology and exploring the potential new pharmacology of exogenous CTS [144]. In the past, most pharmacological studies of CTS were focused on their ability to inhibit NKA. As such, they were used as NKA inhibitors to increase myocardial contractility. Even in this application, clinical studies have demonstrated that the use of lower, but not higher, doses of digoxin is associated with a decrease in mortality in patients with congestive heart failure [145]. Interestingly, recent studies have shown that the activation of NKA signaling, but not inhibition of cellular pump capacity, by CTS is capable of protecting the heart from ischemia/reperfusion injury in rats [146,147,148]. Furthermore, CTS at doses lower than 1/100th of IC50 of NKA activity are effective stimuli of collagen synthesis, suggesting the potential use of these compounds in skin care and wound healing [52,128]. It is equally important to recognize that CTS could also inhibit cell growth in a wide variety of cancer cell lines such as prostate, lung, colon cancer cells and neuroblastoma cells by stimulating several different pathways, including apoptosis and autophage-related processes [61,62,66,70,71,104,149,150,151]. On the other hand, endogenous CTS may play an important role in the pathogenesis of autosomal dominant polycystic kidney disease (ADPKD) by activating Src/EGF receptor/ERK pathways [109,152,153].
Finally, it has been reported that the endocytosis of NKA/Src receptor complex, like many other membrane receptors, is stimulated by its ligands such as CTS [131,154,155]. This occurs via clathrin-coated pits, early and late endosomes, and depends on the activation of Src and PI3K. Although it remains to be further investigated, it is conceivable that CTS-induced endocytosis of receptor NKA/Src could represent a pathway of signal termination. Of course, it might also generate an effective way of communication with intracellular compartments during the signal transduction process [156].
4. Conformation-Dependent Regulation of Src by α1 NKA, a New Hypothesis
The essence of receptor-mediated signal transduction is the intrinsic ability of a receptor to adapt both active and inactive conformational states [157,158]. Several important but seeming un-related studies have led us to test this important concept (hypothesis) in NKA-mediated signal transduction. The first clue was actually from the studies of purified NKA/Src interaction. As reported by Tian et al. [101], purified kidney α1 NKA inhibited Src Y418 phosphorylation, and that addition of ouabain restored Y418 phosphorylation only in the presence of α1 NKA. Interestingly, while vanadate also inhibited ATPase activity of α1 NKA, it showed minimal effect on Y418 phosphorylation at concentrations that produced similar degree of NKA inhibition as ouabain. Most significantly, ouabain was able to further stimulate Y418 phosphorylation in the presence of vanadate that caused complete inhibition of α1 NKA. Because it is known that vanadate facilitates ouabain binding to the α1 NKA, these findings suggest that α1 NKA may interact and regulate Src activity in a conformation-dependent manner. The second line of evidence came from studies of xanthone derivatives. These compounds are potent and specific inhibitors of α1 NKA. However, they show no ouabain-like effect on α1 NKA/Src interaction [159]. This led to studies of Ye et al., testing whether α1 NKA/Src interaction can be modeled on the Albers–Post scheme [134]. By using well-characterized conformation stabilizing chemicals as well as α1 NKA mutant defective in conformation transitions, we find strong evidence that α1 NKA, like G protein-coupled receptors, can adapt both active and inactive conformations to interact and regulate Src. This new framework, taken together with our appreciation of Albers–Post scheme, has led us to deduce that α1 NKA may represent a broad cell signaling mechanism. As such, many ligands of α1 NKA, including CTS and intracellular/extracellular ions, may alter cellular signal transduction through a Src-dependent process (Figure 2).
4.1. NKA/Src/ROS Loop and Disease Progression
ROS participates in various cellular activities [160,161,162,163,164,165]. Our early studies demonstrate that ouabain stimulates ROS generation in a Ras-dependent way via NKA/ Src signaling [55,166]. On the other hand, modification of α1 NKA by ROS has been well documented [125,167,168,169,170,171,172], and such modification by ROS can directly alter the conformation states of α1 NKA [173]. Our new appreciation of α1 NKA/Src signaling mechanism has led to several studies of whether ROS can work similar to CTS on NKA/Src complex. These studies have led to the following observations. First, an increase in H2O2 generation is sufficient to cause the activation of Src and ERK, and stimulated α1 NKA endocytosis in LLC-PK1 cells. Disruption of NKA/Src interaction by either pNaKtide or by the expression of Src-interaction null mutants (A420P) abolishes H2O2-induced Src/ERK activation [126]. On the other hand, ouabain stimulates the generation of ROS, which results in direct carbonylation of Pro 222 and Thr 224 in the α1 subunit of NKA [125]. Moreover, inhibition of the carbonylation by anti-oxidants attenuates ouabain-induced activation of protein kinase cascades. Thus, it is proposed that NKA/Src and ROS form a signal amplification loop allowing not only CTS but also ROS to generate signals from the NKA.
In view of the well-established role of ROS stress in the progression of many chronic diseases, we and others have recently explored whether the newly appreciated NKA/Src/ROS loop is essential for un-regulated ROS signaling. These studies have demonstrated that this signaling loop is indeed activated and plays an important role in the development of atherosclerosis, renal inflammation-induced tissue damage, and metabolic syndrome as well as uremic cardiomyopathy [136,139,174].
4.2. NKA/Src Interaction as a Drug Target
The rationale for targeting NKA-mediated signal transduction to develop new therapeutics has been discussed [6,43,144]. Recent in vitro and in vivo studies have demonstrated the feasibility of targeting NKA/Src interaction, and effectiveness of an inhibitor, pNaKtide, of this interaction as a potential therapeutics of cardio-renal diseases and metabolic syndrome. As discussed above, pNaKtide is composed of NaKtide sequence (20 amino acid peptide) from human α1 NKA and a TAT leader (13 amino acids peptide). The TAT leader is the so-called cell penetrating peptide that helps large molecules to across cell membrane. pNaKtide not only readily passes cell membrane, it resides, similar to α1 NKA, in the plasma membrane, making it highly specific as an inhibitor of NKA/Src complex. Moreover, it is potent, as 0.1 to 1 μM is sufficient to completely block CTS- or ROS-induced signal transduction in cell cultures [112,126]. It also has a good safety profile as no cellular toxicity was observed up to 20 μM in three different cell lines. Remarkably, it is readily taken up in vivo by the heart, kidney, liver and fat tissues, and shows a plasma membrane distribution as well [140,175,176]. Significantly, pNaKtide was effective in blocking ROS amplification and α1 NKA-mediated signal transduction in animals fed with high fat diet, and consequently attenuated metabolic syndrome [140]. Moreover, recent studies have further demonstrated its effectiveness as an inhibitor of ROS amplification and NKA/Src signaling in animal model of chronic kidney failure, Western diet-induced liver damage and atherosclerosis [175,176]. For example, in animal models of uremic cardiomyopathy induced by 5/6 nephrectomy, it not only prevented cardiac hypertrophy and fibrosis, but also improved cardiac function and hematocrit. Remarkably, it was also capable of reversing cardiac lesions in a dose-dependent manner [140,175].
5. Conclusions and Perspectives
Studies from many laboratories of the past 20 years have documented that NKA has an ion-pumping independent receptor function that confers a ligand-like effect of CTS on protein/lipid kinases, intracellular Ca2+ oscillation and ROS generation. Direct protein interactions between NKA and its partners are responsible for this newly appreciated signaling mechanism. Meanwhile, our appreciation of this signaling mechanism has also evolved from “moonlighting” to an essential pathway in animal physiology and disease progression. It is important to recognize that the aforementioned investigations only mark the beginning of a fascinating field. In addition to the continued effort of many in defining the molecular mechanism of NKA-mediated signal transduction, isoform-specificity and identifying cell/tissue-specific signalosomes, the following two areas of research may further advance our understanding of NKA. First, efforts have been and will continue to be made to generate new animal models including transgenic animals with specific defect in NKA signaling and tool drugs targeting NKA/Src or NKA/other partner interactions. These new animal models and tool drugs will help advancing our understanding of NKA-mediated signal transduction in animal physiology and disease progression, provide further validation of NKA-mediated protein interaction as a druggable target, and generate lead candidates for the development of clinically useful drugs.
Second, NKA, as discussed, interacts with many proteins. Unlike other membrane receptors, it is highly expressed in most of cells. As such, it may also work as an important scaffold. As an example, it is of interesting to look at the interaction of α1 NKA with caveolin-1, and a Src-dependent interplay among the α1 NKA, caveolin-1 and cholesterol. Both caveolin-1 and cholesterol are important structural components of caveolae that are flask-shaped vesicular invaginations on the plasma membrane [154,177,178,179]. Caveolae are known to play an important role in cellular signal transduction. The α1 subunit of NKA contains a highly conserved caveolin binding motif at the N-terminus, and brings caveolin-1 to its regulatory kinase Src [107,179]. Reduction in the expression of α1 NKA stimulates Src, resulting in an increase in caveolin-1 Y14 phosphorylation. This leads to the reduction of membrane caveolin-1 and cholesterol, and consequently a decrease in the number of caveolae [179,180]. On the other hand, reduction of membrane cholesterol can activate Src in a α1 NKA-dependent manner, leading to an increase in the endocytosis of α1 NKA. Thus, this Src-dependent interplay may establish a highly efficient feed-forward mechanism that detects the change in cellular cholesterol and/or α1 NKA, and then alters the structure and function of the plasma membrane [107,123,178,179,181,182,183,184,185]. In accordance, studies have shown that α1 NKA/caveolin-1 interaction is not only essential for CTS-induced activation of protein/lipid kinase cascades, but also for α1 NKA to interact with other signaling proteins. One of these is IP3 receptor (Figure 2). This latter interaction may also depend on other scaffolding proteins such as ankyrin. Nevertheless, the interaction between caveolar α1 NKA and ER IP3R allows the formation of an efficient Ca2+ signaling machine by tethering affecters (e.g., membrane receptors), signal transducer (e.g., Src and phospholipase C) and effectors (e.g., Ca2+ channels) together. Consequently, α1 NKA is necessary not only for CTS but also for purinergic stimulation of Ca2+ oscillation [107,124,186]. Clearly, much remains to be learned about the potential role of α1 NKA as scaffold and its interplay with other receptors in animal physiology and disease progression, which could open up new opportunities for the discovery of other NKA-specific drug targets.
Acknowledgments
This work was supported by NIH grants HL109015.
Author Contributions
Xiaoyu Cui wrote and edited the manuscript, Zijian Xie wrote and edited the manuscript.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Skou, J.C. The influence of some cations on an adenosine triphosphatase from peripheral nerves. Biochim. Biophys. Acta 1957, 23, 394–401. [Google Scholar] [CrossRef]
- Quastel, M.R.; Kaplan, J.G. Inhibition by ouabain of human lymphocyte transformation induced by phytohaemagglutinin in vitro. Nature 1968, 219, 198–200. [Google Scholar] [CrossRef] [PubMed]
- Kaplan, J.G. Membrane cation transport and the control of proliferation of mammalian cells. Annu. Rev. Physiol. 1978, 40, 19–41. [Google Scholar] [CrossRef] [PubMed]
- Shiratori, O. Growth inhibitory effect of cardiac glycosides and aglycones on neoplastic cells: In vitro and in vivo studies. GANN Jpn. J. Cancer Res. 1967, 58, 521–528. [Google Scholar]
- Xie, Z.; Cai, T. Na+-K+–ATPase-Mediated signal transduction: From protein interaction to cellular function. Mol. Interv. 2003, 3, 157–168. [Google Scholar] [CrossRef] [PubMed]
- Aperia, A. New roles for an old enzyme: Na,K-ATPase emerges as an interesting drug target. J. Intern. Med. 2007, 261, 44–52. [Google Scholar] [CrossRef] [PubMed]
- Zaleski, S. Carl schmidt. Chem. Ber. 1894, 27, 963–978. [Google Scholar] [CrossRef]
- Heidenhain, R. Neue versuche über die aufsaugung im dünndarm. Pflügers Arch. Eur. J. Physiol. 1894, 56, 579–631. [Google Scholar] [CrossRef]
- Overton, E. Ueber die allgemeinen osmotischen eigenschaften der zelle, ihre vermutlichen ursachen und ihre bedeutung für die physiologie. Vierteljahrsschr. Nat. Ges. Zürich. 1899, 88–135. [Google Scholar]
- Overton, E. Beiträge zur allgemeinen muskel und nervenphysilogie. II. Ueber die unentbehrlichkeit von natrium- (oder lithium) ionen für dencontractionsactdesmuskels. Pflügers Arch. Ges. Physiol. 1902, 346–386. [Google Scholar] [CrossRef]
- Heppel, L.A. The diffusion of radioactive sodium into the muscles of potassium-deprivedrats. Am. J. Physiol. 1940, 128, 449–454. [Google Scholar]
- Steinbach, H.B. Sodium and potassium in frog muscle. J. Biol. Chem. 1940, 133, 695–701. [Google Scholar]
- Dean, R.B. Theories of electrolyte equilibrium in muscle. Biol. Symp. 1941, 3, 331–348. [Google Scholar]
- Solomon, R.Z.; Hald, P.M.; Peters, J.P. The state of the inorganic components of human red blood cells. J. Biol. Chem. 1940, 132, 732–738. [Google Scholar]
- Danowski, T.S. The transfer of potassium across the human blood cell membrane. J. Biol. Chem. 1941, 139, 693–705. [Google Scholar]
- Harris, J.E. The influence of the metabolism of human erythrocytes on their potassiumcontent. J. Biol. Chem. 1941, 141, 579–595. [Google Scholar]
- Schatzmann, H.J. Cardiac glycosides as inhibitors of active potassium and sodium transport by erythrocyte membrane. Helv. Physiol. Pharmacol. Acta 1953, 11, 346–354. [Google Scholar] [PubMed]
- Engelhardt, W.A.; Ljubimowa, M.N. Myosine and adenosinetriphosphatase. Nature 1939, 144, 668–669. [Google Scholar] [CrossRef]
- Gardos, G. Accumulation of potassium ions by human blood corpuscles. Acta Physiol. Hung. 1954, 6, 191–199. [Google Scholar] [PubMed]
- Clarke, R.J.; Fan, X. Pumping ions. Clin. Exp. Pharmacol. Physiol. 2011, 38, 726–733. [Google Scholar] [CrossRef] [PubMed]
- Siegel, G.J.; Albers, R.W. Sodium-Potassium-Activated adenosine triphosphatase of electrophorus electric organ. IV. Modification of responses to sodium and potassium by arsenite plus 2,3-dimercaptopropanol. J. Biol. Chem. 1967, 242, 4972–4979. [Google Scholar] [PubMed]
- Post, R.L.; Kume, S.; Tobin, T.; Orcutt, B.; Sen, A.K. Flexibility of an active center in sodium-plus-potassium adenosine triphosphatase. J. Gen. Physiol. 1969, 54, 306–326. [Google Scholar] [CrossRef] [PubMed]
- Jorgensen, P.L. Isolation and characterization of the components of the sodium pump. Q. Rev. Biophys. 1974, 7, 239–274. [Google Scholar] [CrossRef] [PubMed]
- Jorgensen, P.L. Purification and characterization of (Na+ + K+)-ATPase. VI. Differential tryptic modification of catalytic functions of the purified enzyme in presence of NaCL and KCL. Biochim. Biophys. Acta 1977, 466, 97–108. [Google Scholar] [CrossRef]
- Jorgensen, P.L. Purification and characterization of (Na+, K+)-ATPase. V. Conformational changes in the enzyme transitions between the Na-form and the K-form studied with tryptic digestion as a tool. Biochim. Biophys. Acta 1975, 401, 399–415. [Google Scholar] [CrossRef]
- Jorgensen, P.L.; Klodos, I. Purification and characterization of (Na+ + K+)-ATPase. VII. Tryptic degradation of the Na-form of the enzyme protein resulting in selective modification of dephosphorylation reactions of the (Na+ + K+)-ATPase. Biochim. Biophys. Acta 1978, 507, 8–16. [Google Scholar] [CrossRef]
- Blanco, G.; Mercer, R.W. Isozymes of the Na-K-ATPase: Heterogeneity in structure, diversity in function. Am. J. Physiol. 1998, 275, F633–F650. [Google Scholar] [PubMed]
- Sweadner, K.J. Isozymes of the Na+/K+-ATPase. Biochim. Biophys. Acta 1989, 988, 185–220. [Google Scholar] [CrossRef]
- Marks, M.J.; Seeds, N.W. A heterogeneous ouabain-ATPase interaction in mouse brain. Life Sci. 1978, 23, 2735–2744. [Google Scholar] [CrossRef]
- Sweadner, K.J. Two molecular forms of (Na+ + K+)-stimulated ATPase in brain. Separation, and difference in affinity for strophanthidin. J. Biol. Chem. 1979, 254, 6060–6067. [Google Scholar] [PubMed]
- Shull, G.E.; Schwartz, A.; Lingrel, J.B. Amino-Acid sequence of the catalytic subunit of the (Na+ + K+)ATPase deduced from a complementary DNA. Nature 1985, 316, 691–695. [Google Scholar] [CrossRef] [PubMed]
- Shull, G.E.; Greeb, J.; Lingrel, J.B. Molecular cloning of three distinct forms of the Na+,K+-ATPase α-subunit from rat brain. Biochemistry 1986, 25, 8125–8132. [Google Scholar] [CrossRef] [PubMed]
- Shamraj, O.I.; Lingrel, J.B. A putative fourth Na+,K(+)-ATPase α-subunit gene is expressed in testis. Proc. Natl. Acad. Sci. USA 1994, 91, 12952–12956. [Google Scholar] [CrossRef] [PubMed]
- Sanchez, G.; Nguyen, A.N.; Timmerberg, B.; Tash, J.S.; Blanco, G. The Na,K-ATPase α4 isoform from humans has distinct enzymatic properties and is important for sperm motility. Mol. Hum. Reprod. 2006, 12, 565–576. [Google Scholar] [CrossRef] [PubMed]
- Jorgensen, P.L.; Andersen, J.P. Structural basis for E1-E2 conformational transitions in Na,K-pump and Ca-pump proteins. J. Membr. Biol. 1988, 103, 95–120. [Google Scholar] [CrossRef] [PubMed]
- Morth, J.P.; Pedersen, B.P.; Toustrup-Jensen, M.S.; Sorensen, T.L.; Petersen, J.; Andersen, J.P.; Vilsen, B.; Nissen, P. Crystal structure of the sodium-potassium pump. Nature 2007, 450, 1043–1049. [Google Scholar] [CrossRef] [PubMed]
- Nyblom, M.; Poulsen, H.; Gourdon, P.; Reinhard, L.; Andersson, M.; Lindahl, E.; Fedosova, N.; Nissen, P. Crystal structure of Na+, K(+)-ATPase in the Na(+)-bound state. Science 2013, 342, 123–127. [Google Scholar] [CrossRef] [PubMed]
- Morth, J.P.; Pedersen, B.P.; Buch-Pedersen, M.J.; Andersen, J.P.; Vilsen, B.; Palmgren, M.G.; Nissen, P. A structural overview of the plasma membrane Na+, K+-ATPase and H+-ATPase ion pumps. Nat. Rev. Mol. Cell Biol. 2011, 12, 60–70. [Google Scholar] [CrossRef] [PubMed]
- Ogawa, H.; Cornelius, F.; Hirata, A.; Toyoshima, C. Sequential substitution of K(+) bound to Na(+),K(+)-ATPase visualized by X-ray crystallography. Nat. Commun. 2015, 6, 8004. [Google Scholar] [CrossRef] [PubMed]
- Laursen, M.; Gregersen, J.L.; Yatime, L.; Nissen, P.; Fedosova, N.U. Structures and characterization of digoxin- and bufalin-bound Na+, K+-ATPase compared with the ouabain-bound complex. Proc. Natl. Acad. Sci. USA 2015, 112, 1755–1760. [Google Scholar] [CrossRef] [PubMed]
- Laursen, M.; Yatime, L.; Nissen, P.; Fedosova, N.U. Crystal structure of the high-affinity Na+ K+-ATPase-ouabain complex with Mg2+ bound in the cation binding site. Proc. Natl. Acad. Sci. USA 2013, 110, 10958–10963. [Google Scholar] [CrossRef] [PubMed]
- Akera, T.; Brody, T.M. Inotropic action of digitalis and ion transport. Life Sci. 1976, 18, 135–144. [Google Scholar] [CrossRef]
- Schoner, W.; Scheiner-Bobis, G. Endogenous and exogenous cardiac glycosides and their mechanisms of action. Am. J. Cardiovasc. Drugs 2007, 7, 173–189. [Google Scholar] [CrossRef] [PubMed]
- Pierdomenico, S.D.; Bucci, A.; Manunta, P.; Rivera, R.; Ferrandi, M.; Hamlyn, J.M.; Lapenna, D.; Cuccurullo, F.; Mezzetti, A. Endogenous ouabain and hemodynamic and left ventricular geometric patterns in essential hypertension. Am. J. Hypertens. 2001, 14, 44–50. [Google Scholar] [CrossRef]
- Wang, J.G.; Staessen, J.A.; Messaggio, E.; Nawrot, T.; Fagard, R.; Hamlyn, J.M.; Bianchi, G.; Manunta, P. Salt, endogenous ouabain and blood pressure interactions in the general population. J. Hypertens. 2003, 21, 1475–1481. [Google Scholar] [CrossRef] [PubMed]
- Manunta, P.; Stella, P.; Rivera, R.; Ciurlino, D.; Cusi, D.; Ferrandi, M.; Hamlyn, J.M.; Bianchi, G. Left ventricular mass, stroke volume, and ouabain-like factor in essential hypertension. Hypertension 1999, 34, 450–456. [Google Scholar] [CrossRef] [PubMed]
- Cuff, J.M.; Lichtman, A. The early effects of ouabain on potassium metabolism and rate of proliferation of mouse lymphoblasts. J. Cell Physiol. 1975, 85, 209–215. [Google Scholar] [CrossRef] [PubMed]
- Pollack, L.R.; Tate, E.H.; Cook, J.S. Na+,K+-ATPase in hela cells after prolonged growth in low K+ or ouabain. J. Cell Physiol. 1981, 106, 85–97. [Google Scholar] [CrossRef] [PubMed]
- Touza, N.A.; Pocas, E.S.; Quintas, L.E.; Cunha-Filho, G.; Santos, M.L.; Noel, F. Inhibitory effect of combinations of digoxin and endogenous cardiotonic steroids on Na+/K+-ATPase activity in human kidney membrane preparation. Life Sci. 2011, 88, 39–42. [Google Scholar] [CrossRef] [PubMed]
- Lifshitz, Y.; Petrovich, E.; Haviv, H.; Goldshleger, R.; Tal, D.M.; Garty, H.; Karlish, S.J. Purification of the human α2 isoform of Na,K-ATPase expressed in pichia pastoris. Stabilization by lipids and FXYD1. Biochemistry 2007, 46, 14937–14950. [Google Scholar] [CrossRef] [PubMed]
- Blanco, G. The Na/K-ATPase and its isozymes: What we have learned using the baculovirus expression system. Front. Biosci. 2005, 10, 2397–2411. [Google Scholar] [CrossRef] [PubMed]
- El-Okdi, N.; Smaili, S.; Raju, V.; Shidyak, A.; Gupta, S.; Fedorova, L.; Elkareh, J.; Periyasamy, S.; Shapiro, A.P.; Kahaleh, M.B.; et al. Effects of cardiotonic steroids on dermal collagen synthesis and wound healing. J. Appl. Physiol. 2008, 105, 30–36. [Google Scholar] [CrossRef] [PubMed]
- Huang, L.; Li, H.; Xie, Z. Ouabain-Induced hypertrophy in cultured cardiac myocytes is accompanied by changes in expression of several late response genes. J. Mol. Cell Cardiol. 1997, 29, 429–437. [Google Scholar] [CrossRef] [PubMed]
- Kometiani, P.; Li, J.; Gnudi, L.; Kahn, B.B.; Askari, A.; Xie, Z. Multiple signal transduction pathways link Na+/K+-ATPase to growth-related genes in cardiac myocytes. The roles of ras and mitogen-activated protein kinases. J. Biol. Chem. 1998, 273, 15249–15256. [Google Scholar] [CrossRef] [PubMed]
- Xie, Z.; Kometiani, P.; Liu, J.; Li, J.; Shapiro, J.I.; Askari, A. Intracellular reactive oxygen species mediate the linkage of Na+/K+-ATPase to hypertrophy and its marker genes in cardiac myocytes. J. Biol. Chem. 1999, 274, 19323–19328. [Google Scholar] [CrossRef] [PubMed]
- Haas, M.; Askari, A.; Xie, Z. Involvement of Src and epidermal growth factor receptor in the signal-transducing function of Na+/K+-ATPase. J. Biol. Chem. 2000, 275, 27832–27837. [Google Scholar] [CrossRef] [PubMed]
- Haas, M.; Wang, H.; Tian, J.; Xie, Z. Src-Mediated inter-receptor cross-talk between the Na+/K+-ATPase and the epidermal growth factor receptor relays the signal from ouabain to mitogen-activated protein kinases. J. Biol. Chem. 2002, 277, 18694–18702. [Google Scholar] [CrossRef] [PubMed]
- Dmitrieva, R.I.; Doris, P.A. Ouabain is a potent promoter of growth and activator of ERK1/2 in ouabain-resistant rat renal epithelial cells. J. Biol. Chem. 2003, 278, 28160–28166. [Google Scholar] [CrossRef] [PubMed]
- Ferrandi, M.; Molinari, I.; Barassi, P.; Minotti, E.; Bianchi, G.; Ferrari, P. Organ hypertrophic signaling within caveolae membrane subdomains triggered by ouabain and antagonized by PST 2238. J. Biol. Chem. 2004, 279, 33306–33314. [Google Scholar] [CrossRef] [PubMed]
- Trevisi, L.; Visentin, B.; Cusinato, F.; Pighin, I.; Luciani, S. Antiapoptotic effect of ouabain on human umbilical vein endothelial cells. Biochem. Biophys. Res. Commun. 2004, 321, 716–721. [Google Scholar] [CrossRef] [PubMed]
- Lopez-Lazaro, M.; Pastor, N.; Azrak, S.S.; Ayuso, M.J.; Austin, C.A.; Cortes, F. Digitoxin inhibits the growth of cancer cell lines at concentrations commonly found in cardiac patients. J. Nat. Prod. 2005, 68, 1642–1645. [Google Scholar] [CrossRef] [PubMed]
- Bielawski, K.; Winnicka, K.; Bielawska, A. Inhibition of DNA topoisomerases I and II, and growth inhibition of breast cancer MCF-7 cells by ouabain, digoxin and proscillaridin A. Biol. Pharm. Bull. 2006, 29, 1493–1497. [Google Scholar] [CrossRef] [PubMed]
- Golden, W.C.; Martin, L.J. Low-Dose ouabain protects against excitotoxic apoptosis and up-regulates nuclear Bcl-2 in vivo. Neuroscience 2006, 137, 133–144. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Zelenin, S.; Aperia, A.; Aizman, O. Low doses of ouabain protect from serum deprivation-triggered apoptosis and stimulate kidney cell proliferation via activation of NF-κB. J. Am. Soc. Nephrol. 2006, 17, 1848–1857. [Google Scholar] [CrossRef] [PubMed]
- Ramirez-Ortega, M.; Maldonado-Lagunas, V.; Melendez-Zajgla, J.; Carrillo-Hernandez, J.F.; Pastelin-Hernandez, G.; Picazo-Picazo, O.; Ceballos-Reyes, G. Proliferation and apoptosis of hela cells induced by in vitro stimulation with digitalis. Eur. J. Pharmacol. 2006, 534, 71–76. [Google Scholar] [CrossRef] [PubMed]
- Kulikov, A.; Eva, A.; Kirch, U.; Boldyrev, A.; Scheiner-Bobis, G. Ouabain activates signaling pathways associated with cell death in human neuroblastoma. Biochim. Biophys. Acta 2007, 1768, 1691–1702. [Google Scholar] [CrossRef] [PubMed]
- Li, M.; Wang, Q.; Guan, L. Effects of ouabain on proliferation, intracellular free calcium and c-myc mRNA expression in vascular smooth muscle cells. J. Comp. Physiol. B 2007, 177, 589–595. [Google Scholar] [CrossRef] [PubMed]
- Mijatovic, T.; Roland, I.; van Quaquebeke, E.; Nilsson, B.; Mathieu, A.; van Vynckt, F.; Darro, F.; Blanco, G.; Facchini, V.; Kiss, R. The α subunit of the sodium pump could represent a novel target to combat non-small cell lung cancers. J. Pathol. 2007, 212, 170–179. [Google Scholar] [CrossRef] [PubMed]
- Mijatovic, T.; de Neve, N.; Gailly, P.; Mathieu, V.; Haibe-Kains, B.; Bontempi, G.; Lapeira, J.; Decaestecker, C.; Facchini, V.; Kiss, R. Nucleolus and c-myc: Potential targets of cardenolide-mediated antitumor activity. Mol. Cancer Ther. 2008, 7, 1285–1296. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.; Zheng, M.; Li, Z.; Li, R.; Jia, L.; Xiong, X.; Southall, N.; Wang, S.; Xia, M.; Austin, C.P.; et al. Cardiac glycosides inhibit p53 synthesis by a mechanism relieved by Src or MAPK inhibition. Cancer Res. 2009, 69, 6556–6564. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Zhan, Y.; Xu, R.; Shao, R.; Jiang, J.; Wang, Z. Src mediates extracellular signal-regulated kinase 1/2 activation and autophagic cell death induced by cardiac glycosides in human non-small cell lung cancer cell lines. Mol. Carcinog. 2015, 54, E26–E34. [Google Scholar] [CrossRef] [PubMed]
- Patel, S. Plant-Derived cardiac glycosides: Role in heart ailments and cancer management. Biomed. Pharmacother. 2016, 84, 1036–1041. [Google Scholar] [CrossRef] [PubMed]
- Cornelius, F.; Kanai, R.; Toyoshima, C. A structural view on the functional importance of the sugar moiety and steroid hydroxyls of cardiotonic steroids in binding to Na,K-ATPase. J. Biol. Chem. 2013, 288, 6602–6616. [Google Scholar] [CrossRef] [PubMed]
- Zeino, M.; Brenk, R.; Gruber, L.; Zehl, M.; Urban, E.; Kopp, B.; Efferth, T. Cytotoxicity of cardiotonic steroids in sensitive and multidrug-resistant leukemia cells and the link with Na(+)/K(+)-ATPase. J. Steroid Biochem. Mol. Biol. 2015, 150, 97–111. [Google Scholar] [CrossRef] [PubMed]
- Estape, E.S.; Torres-Negron, I.; Gonzalez, L.; Martinez-Maldonado, M. A new animal model to study endogenous cardiotonic steroids and the progression of cardiovascular events in salt-sensitive hypertension. Int. Arch. Trans. Med. 2015, 1, 002. [Google Scholar] [CrossRef]
- Blaustein, M.P.; Juhaszova, M.; Golovina, V.A. The cellular mechanism of action of cardiotonic steroids: A new hypothesis. Clin. Exp. Hypertens. 1998, 20, 691–703. [Google Scholar] [CrossRef] [PubMed]
- Langer, G.A. Effects of digitalis on myocardial ionic exchange. Circulation 1972, 46, 180–187. [Google Scholar] [CrossRef] [PubMed]
- Moore, E.D.; Etter, E.F.; Philipson, K.D.; Carrington, W.A.; Fogarty, K.E.; Lifshitz, L.M.; Fay, F.S. Coupling of the Na+/Ca2+ exchanger, Na+/K+ pump and sarcoplasmic reticulum in smooth muscle. Nature 1993, 365, 657–660. [Google Scholar] [CrossRef] [PubMed]
- Juhaszova, M.; Blaustein, M.P. Distinct distribution of different Na+ pump α subunit isoforms in plasmalemma. Physiological implications. Ann. N. Y. Acad. Sci. 1997, 834, 524–536. [Google Scholar] [CrossRef] [PubMed]
- Song, H.; Lee, M.Y.; Kinsey, S.P.; Weber, D.J.; Blaustein, M.P. An N-terminal sequence targets and tethers Na+ pump α2 subunits to specialized plasma membrane microdomains. J. Biol. Chem. 2006, 281, 12929–12940. [Google Scholar] [CrossRef] [PubMed]
- Pawson, T. Protein modules and signalling networks. Nature 1995, 373, 573–580. [Google Scholar] [CrossRef] [PubMed]
- Ren, S.; Yang, G.; He, Y.; Wang, Y.; Li, Y.; Chen, Z. The conservation pattern of short linear motifs is highly correlated with the function of interacting protein domains. BMC Genom. 2008, 9, 452. [Google Scholar] [CrossRef] [PubMed]
- Nelson, W.J.; Veshnock, P.J. Ankyrin binding to (Na+ + K+)ATPase and implications for the organization of membrane domains in polarized cells. Nature 1987, 328, 533–536. [Google Scholar] [CrossRef] [PubMed]
- Ferrandi, M.; Salardi, S.; Tripodi, G.; Barassi, P.; Rivera, R.; Manunta, P.; Goldshleger, R.; Ferrari, P.; Bianchi, G.; Karlish, S.J. Evidence for an interaction between adducin and Na(+)-K(+)-ATPase: Relation to genetic hypertension. Am. J. Physiol. 1999, 277, H1338–H1349. [Google Scholar] [PubMed]
- Tripodi, G.; Valtorta, F.; Torielli, L.; Chieregatti, E.; Salardi, S.; Trusolino, L.; Menegon, A.; Ferrari, P.; Marchisio, P.C.; Bianchi, G. Hypertension-Associated point mutations in the adducin α and β subunits affect actin cytoskeleton and ion transport. J. Clin. Investig. 1996, 97, 2815–2822. [Google Scholar] [CrossRef] [PubMed]
- Sweadner, K.J.; Rael, E. The FXYD gene family of small ion transport regulators or channels: cDNA sequence, protein signature sequence, and expression. Genomics 2000, 68, 41–56. [Google Scholar] [CrossRef] [PubMed]
- Geering, K. FXYD proteins: New regulators of Na-K-ATPase. Am. J. Physiol. Renal. Physiol. 2006, 290, F241–F250. [Google Scholar] [CrossRef] [PubMed]
- Ogawa, H.; Shinoda, T.; Cornelius, F.; Toyoshima, C. Crystal structure of the sodium-potassium pump (Na+, K+-ATPase) with bound potassium and ouabain. Proc. Natl. Acad. Sci. USA 2009, 106, 13742–13747. [Google Scholar] [CrossRef] [PubMed]
- Yatime, L.; Laursen, M.; Morth, J.P.; Esmann, M.; Nissen, P.; Fedosova, N.U. Structural insights into the high affinity binding of cardiotonic steroids to the Na+, K+-ATPase. J. Struct. Biol. 2011, 174, 296–306. [Google Scholar] [CrossRef] [PubMed]
- Palmer, C.J.; Scott, B.T.; Jones, L.R. Purification and complete sequence determination of the major plasma membrane substrate for cAMP-dependent protein kinase and protein kinase C in myocardium. J. Biol. Chem. 1991, 266, 11126–11130. [Google Scholar] [PubMed]
- Walaas, S.I.; Czernik, A.J.; Olstad, O.K.; Sletten, K.; Walaas, O. Protein kinase C and cyclic AMP-dependent protein kinase phosphorylate phospholemman, an insulin and adrenaline-regulated membrane phosphoprotein, at specific sites in the carboxy terminal domain. Biochem. J. 1994, 304, 635–640. [Google Scholar] [CrossRef] [PubMed]
- Lansbery, K.L.; Burcea, L.C.; Mendenhall, M.L.; Mercer, R.W. Cytoplasmic targeting signals mediate delivery of phospholemman to the plasma membrane. Am. J. Physiol. Cell Physiol. 2006, 290, C1275–C1286. [Google Scholar] [CrossRef] [PubMed]
- Bibert, S.; Roy, S.; Schaer, D.; Horisberger, J.D.; Geering, K. Phosphorylation of phospholemman (FXYD1) by protein kinases A and C modulates distinct Na,K-ATPase isozymes. J. Biol. Chem. 2008, 283, 476–486. [Google Scholar] [CrossRef] [PubMed]
- Cheung, J.Y.; Rothblum, L.I.; Moorman, J.R.; Tucker, A.L.; Song, J.; Ahlers, B.A.; Carl, L.L.; Wang, J.; Zhang, X.Q. Regulation of cardiac Na+/Ca2+ exchanger by phospholemman. Ann. N. Y. Acad. Sci. 2007, 1099, 119–134. [Google Scholar] [CrossRef] [PubMed]
- Despa, S.; Bossuyt, J.; Han, F.; Ginsburg, K.S.; Jia, L.G.; Kutchai, H.; Tucker, A.L.; Bers, D.M. Phospholemman-Phosphorylation mediates the β-adrenergic effects on Na/K pump function in cardiac myocytes. Circ. Res. 2005, 97, 252–259. [Google Scholar] [CrossRef] [PubMed]
- Efendiev, R.; Chen, Z.; Krmar, R.T.; Uhles, S.; Katz, A.I.; Pedemonte, C.H.; Bertorello, A.M. The 14-3-3 protein translates the Na+,K+-ATPase α1-subunit phosphorylation signal into binding and activation of phosphoinositide 3-kinase during endocytosis. J. Biol. Chem. 2005, 280, 16272–16277. [Google Scholar] [CrossRef] [PubMed]
- Yudowski, G.A.; Efendiev, R.; Pedemonte, C.H.; Katz, A.I.; Berggren, P.O.; Bertorello, A.M. Phosphoinositide-3 kinase binds to a proline-rich motif in the Na+, K+-ATPase α subunit and regulates its trafficking. Proc. Natl. Acad. Sci. USA 2000, 97, 6556–6561. [Google Scholar] [CrossRef] [PubMed]
- Efendiev, R.; Budu, C.E.; Bertorello, A.M.; Pedemonte, C.H. G-Protein-Coupled receptor-mediated traffic of Na,K-ATPase to the plasma membrane requires the binding of adaptor protein 1 to a Tyr-255-based sequence in the α-subunit. J. Biol. Chem. 2008, 283, 17561–17567. [Google Scholar] [CrossRef] [PubMed]
- Lauf, P.K.; Alqahtani, T.; Flues, K.; Meller, J.; Adragna, N.C. Interaction between Na-K-ATPase and Bcl-2 proteins BclXL and BAK. Am. J. Physiol. Cell Physiol. 2015, 308, C51–C60. [Google Scholar] [CrossRef] [PubMed]
- Thomas, S.M.; Brugge, J.S. Cellular functions regulated by Src family kinases. Annu. Rev. Cell Dev. Biol. 1997, 13, 513–609. [Google Scholar] [CrossRef] [PubMed]
- Tian, J.; Cai, T.; Yuan, Z.; Wang, H.; Liu, L.; Haas, M.; Maksimova, E.; Huang, X.Y.; Xie, Z.J. Binding of Src to Na+/K+-ATPase forms a functional signaling complex. Mol. Biol. Cell 2006, 17, 317–326. [Google Scholar] [CrossRef] [PubMed]
- Contreras, R.G.; Shoshani, L.; Flores-Maldonado, C.; Lazaro, A.; Cereijido, M. Relationship between Na(+),K(+)-ATPase and cell attachment. J. Cell Sci. 1999, 112, 4223–4232. [Google Scholar] [PubMed]
- Aydemir-Koksoy, A.; Abramowitz, J.; Allen, J.C. Ouabain-Induced signaling and vascular smooth muscle cell proliferation. J. Biol. Chem. 2001, 276, 46605–46611. [Google Scholar] [CrossRef] [PubMed]
- Kometiani, P.; Liu, L.; Askari, A. Digitalis-Induced signaling by Na+/K+-ATPase in human breast cancer cells. Mol. Pharmacol. 2005, 67, 929–936. [Google Scholar] [CrossRef] [PubMed]
- Kotova, O.; Al-Khalili, L.; Talia, S.; Hooke, C.; Fedorova, O.V.; Bagrov, A.Y.; Chibalin, A.V. Cardiotonic steroids stimulate glycogen synthesis in human skeletal muscle cells via a Src- and ERK1/2-dependent mechanism. J. Biol. Chem. 2006, 281, 20085–20094. [Google Scholar] [CrossRef] [PubMed]
- Kennedy, D.J.; Vetteth, S.; Periyasamy, S.M.; Kanj, M.; Fedorova, L.; Khouri, S.; Kahaleh, M.B.; Xie, Z.; Malhotra, D.; Kolodkin, N.I.; et al. Central role for the cardiotonic steroid marinobufagenin in the pathogenesis of experimental uremic cardiomyopathy. Hypertension 2006, 47, 488–495. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Haas, M.; Liang, M.; Cai, T.; Tian, J.; Li, S.; Xie, Z. Ouabain assembles signaling cascades through the caveolar Na+/K+-ATPase. J. Biol. Chem. 2004, 279, 17250–17259. [Google Scholar] [CrossRef] [PubMed]
- Ferrandi, M.; Molinari, I.; Torielli, L.; Padoani, G.; Salardi, S.; Rastaldi, M.P.; Ferrari, P.; Bianchi, G. Adducin- and ouabain-related gene variants predict the antihypertensive activity of rostafuroxin, part 1: Experimental studies. Sci. Trans. Med. 2010, 2, 59ra86. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, A.-N.T.; Wallace, D.P.; Blanco, G. Ouabain binds with high affinity to the Na,K-ATPase in human polycystic kidney cells. J. Am. Soc. Nephrol. 2007, 18, 46–57. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, A.N.; Jansson, K.; Sanchez, G.; Sharma, M.; Reif, G.A.; Wallace, D.P.; Blanco, G. Ouabain activates the Na-K-ATPase signalosome to induce autosomal dominant polycystic kidney disease cell proliferation. AJP Renal Physiol. 2011, 301, F897–F906. [Google Scholar] [CrossRef] [PubMed]
- Clifford, R.J.; Kaplan, J.H. Human breast tumor cells are more resistant to cardiac glycoside toxicity than non-tumorigenic breast cells. PLoS ONE 2013, 8, e84306. [Google Scholar] [CrossRef] [PubMed]
- Li, Z.; Cai, T.; Tian, J.; Xie, J.X.; Zhao, X.; Liu, L.; Shapiro, J.I.; Xie, Z. Naktide, a Na/K-ATPase-derived peptide Src inhibitor, antagonizes ouabain-activated signal transduction in cultured cells. J. Biol. Chem. 2009, 284, 21066–21076. [Google Scholar] [CrossRef] [PubMed]
- Li, Z.; Zhang, Z.; Xie, J.X.; Li, X.; Tian, J.; Cai, T.; Cui, H.; Ding, H.; Shapiro, J.I.; Xie, Z. Na/k-ATPase mimetic pnaktide peptide inhibits the growth of human cancer cells. J. Biol. Chem. 2011, 286, 32394–32403. [Google Scholar] [CrossRef] [PubMed]
- Banerjee, M.; Duan, Q.; Xie, Z. SH2 ligand-like effects of second cytosolic domain of Na/K-ATPase α1 subunit on Src kinase. PLoS ONE 2015, 10, e0142119. [Google Scholar] [CrossRef] [PubMed]
- Yosef, E.; Katz, A.; Peleg, Y.; Mehlman, T.; Karlish, S.J. Do Src kinase and caveolin interact directly with Na,K-ATPase? J. Biol. Chem. 2016, 291, 11736–11750. [Google Scholar] [CrossRef] [PubMed]
- Brott, B.K.; Decker, S.; O’Brien, M.C.; Jove, R. Molecular features of the viral and cellular Src kinases involved in interactions with the gtpase-activating protein. Mol. Cell Biol. 1991, 11, 5059–5067. [Google Scholar] [CrossRef] [PubMed]
- Shvartsman, D.E.; Donaldson, J.C.; Diaz, B.; Gutman, O.; Martin, G.S.; Henis, Y.I. Src kinase activity and SH2 domain regulate the dynamics of Src association with lipid and protein targets. J. Cell Biol. 2007, 178, 675–686. [Google Scholar] [CrossRef] [PubMed]
- Lai, F.; Madan, N.; Ye, Q.; Duan, Q.; Li, Z.; Wang, S.; Si, S.; Xie, Z. Identification of a mutant α1 Na/K-ATPase that pumps but is defective in signal transduction. J. Biol. Chem. 2013, 288, 13295–13304. [Google Scholar] [CrossRef] [PubMed]
- Madan, N.; Xu, Y.; Duan, Q.; Banerjee, M.; Larre, I.; Pierre, S.V.; Xie, Z. Src-Independent ERK signaling through the rat α3 isoform of Na/K-ATPase. Am. J. Physiol. Cell Physiol. 2017, 312, C222–C232. [Google Scholar] [CrossRef] [PubMed]
- Xie, J.; Ye, Q.; Cui, X.; Madan, N.; Yi, Q.; Pierre, S.V.; Xie, Z. Expression of rat Na-K-ATPase α2 enables ion pumping but not ouabain-induced signaling in α1-deficient porcine renal epithelial cells. Am. J. Physiol. 2015, 309, 373–382. [Google Scholar] [CrossRef] [PubMed]
- Gable, M.E.; Abdallah, S.L.; Najjar, S.M.; Liu, L.; Askari, A. Digitalis-Induced cell signaling by the sodium pump: On the relation of Src to Na(+)/K(+)-ATPase. Biochem. Biophys. Res. Commun. 2014, 446, 1151–1154. [Google Scholar] [CrossRef] [PubMed]
- Weigand, K.M.; Swarts, H.G.; Fedosova, N.U.; Russel, F.G.; Koenderink, J.B. Na,K-ATPase activity modulates Src activation: A role for ATP/ADP ratio. Biochim. Biophys. Acta 2012, 1818, 1269–1273. [Google Scholar] [CrossRef] [PubMed]
- Liu, L.; Mohammadi, K.; Aynafshar, B.; Wang, H.; Li, D.; Liu, J.; Ivanov, A.V.; Xie, Z.; Askari, A. Role of caveolae in signal-transducing function of cardiac Na+/K+-ATPase. Am. J. Physiol. Cell Physiol. 2003, 284, C1550–C1560. [Google Scholar] [CrossRef] [PubMed]
- Yuan, Z.; Cai, T.; Tian, J.; Ivanov, A.V.; Giovannucci, D.R.; Xie, Z. Na/K-ATPase tethers phospholipase C and IP3 receptor into a calcium-regulatory complex. Mol. Biol. Cell 2005, 16, 4034–4045. [Google Scholar] [CrossRef] [PubMed]
- Yan, Y.; Shapiro, A.P.; Haller, S.; Katragadda, V.; Liu, L.; Tian, J.; Basrur, V.; Malhotra, D.; Xie, Z.J.; Abraham, N.G.; et al. Involvement of reactive oxygen species in a feed-forward mechanism of Na/K-ATPase-mediated signaling transduction. J. Biol. Chem. 2013, 288, 34249–34258. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Ye, Q.; Liu, C.; Xie, J.X.; Yan, Y.; Lai, F.; Duan, Q.; Li, X.; Tian, J.; Xie, Z. Involvement of Na/K-ATPase in hydrogen peroxide-induced activation of the Src/ERK pathway in LLC-PK1 cells. Free Radic. Biol. Med. 2014, 71, 415–426. [Google Scholar] [CrossRef] [PubMed]
- Lingrel, J.B. The physiological significance of the cardiotonic steroid/ouabain-binding site of the Na,K-ATPase. Annu. Rev. Physiol. 2010, 72, 395–412. [Google Scholar] [CrossRef] [PubMed]
- Quintas, L.E.; Pierre, S.V.; Liu, L.; Bai, Y.; Liu, X.; Xie, Z.J. Alterations of Na+/K+-ATPase function in caveolin-1 knockout cardiac fibroblasts. J. Mol. Cell Cardiol. 2010, 49, 525–531. [Google Scholar] [CrossRef] [PubMed]
- Fontana, J.M.; Burlaka, I.; Khodus, G.; Brismar, H.; Aperia, A. Calcium oscillations triggered by cardiotonic steroids. FEBS J. 2013, 280, 5450–5455. [Google Scholar] [CrossRef] [PubMed]
- Dvela, M.; Rosen, H.; Ben-Ami, H.C.; Lichtstein, D. Endogenous ouabain regulates cell viability. Am. J. Physiol. Cell Physiol. 2012, 302, C442–C452. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Kesiry, R.; Periyasamy, S.M.; Malhotra, D.; Xie, Z.; Shapiro, J.I. Ouabain induces endocytosis of plasmalemmal Na/K-ATPase in LLC-PK1 cells by a clathrin-dependent mechanism. Kidney Int. 2004, 66, 227–241. [Google Scholar] [CrossRef] [PubMed]
- Liu, L.; Zhao, X.; Pierre, S.V.; Askari, A. Association of PI3K-Akt signaling pathway with digitalis-induced hypertrophy of cardiac myocytes. Am. J. Physiol. Cell Physiol. 2007, 293, C1489–C1497. [Google Scholar] [CrossRef] [PubMed]
- Wu, J.; Akkuratov, E.E.; Bai, Y.; Gaskill, C.M.; Askari, A.; Liu, L. Cell signaling associated with Na(+)/K(+)-ATPase: Activation of phosphatidylinositide 3-kinase IA/Akt by ouabain is independent of Src. Biochemistry 2013, 52, 9059–9067. [Google Scholar] [CrossRef] [PubMed]
- Ye, Q.; Lai, F.; Banerjee, M.; Duan, Q.; Li, Z.; Si, S.; Xie, Z. Expression of mutant α1 Na/K-ATPase defective in conformational transition attenuates Src-mediated signal transduction. J. Biol. Chem. 2013, 288, 5803–5814. [Google Scholar] [CrossRef] [PubMed]
- Elkareh, J.; Kennedy, D.J.; Yashaswi, B.; Vetteth, S.; Shidyak, A.; Kim, E.G.; Smaili, S.; Periyasamy, S.M.; Hariri, I.M.; Fedorova, L.; et al. Marinobufagenin stimulates fibroblast collagen production and causes fibrosis in experimental uremic cardiomyopathy. Hypertension 2007, 49, 215–224. [Google Scholar] [CrossRef] [PubMed]
- Kennedy, D.J.; Chen, Y.; Huang, W.; Viterna, J.; Liu, J.; Westfall, K.; Tian, J.; Bartlett, D.J.; Tang, W.H.; Xie, Z.; et al. CD36 and Na/K-ATPase-α1 form a proinflammatory signaling loop in kidney. Hypertension 2013, 61, 216–224. [Google Scholar] [CrossRef] [PubMed]
- Tian, J.; Shidyak, A.; Periyasamy, S.M.; Haller, S.; Taleb, M.; El-Okdi, N.; Elkareh, J.; Gupta, S.; Gohara, S.; Fedorova, O.V.; et al. Spironolactone attenuates experimental uremic cardiomyopathy by antagonizing marinobufagenin. Hypertension 2009, 54, 1313–1320. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Yan, Y.; Liu, L.; Xie, Z.; Malhotra, D.; Joe, B.; Shapiro, J.I. Impairment of Na/K-ATPase signaling in renal proximal tubule contributes to Dahl salt-sensitive hypertension. J. Biol. Chem. 2011, 286, 22806–22813. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.; Kennedy, D.J.; Ramakrishnan, D.P.; Yang, M.; Huang, W.; Li, Z.; Xie, Z.; Chadwick, A.C.; Sahoo, D.; Silverstein, R.L. Oxidized LDL-bound CD36 recruits an Na+/K+-ATPase-Lyn complex in macrophages that promotes atherosclerosis. Sci. Signal. 2015, 8, ra91. [Google Scholar] [CrossRef] [PubMed]
- Sodhi, K.; Maxwell, K.; Yan, Y.; Liu, J.; Chaudhry, M.A.; Getty, M.; Xie, Z.; Abraham, N.G.; Shapiro, J.I. Pnaktide inhibits Na/K-ATPase reactive oxygen species amplification and attenuates adipogenesis. Sci. Adv. 2015, 1, e1500781. [Google Scholar] [CrossRef] [PubMed]
- Dvela-Levitt, M.; Cohen-Ben Ami, H.; Rosen, H.; Ornoy, A.; Hochner-Celnikier, D.; Granat, M.; Lichtstein, D. Reduction in maternal circulating ouabain impairs offspring growth and kidney development. J. Am. Soc. Nephrol. 2015, 26, 1103–1114. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Khodus, G.R.; Kruusmagi, M.; Kamali-Zare, P.; Liu, X.L.; Eklof, A.C.; Zelenin, S.; Brismar, H.; Aperia, A. Ouabain protects against adverse developmental programming of the kidney. Nat. Commun. 2010, 1, 42. [Google Scholar] [CrossRef] [PubMed]
- Song, H.; Karashima, E.; Hamlyn, J.M.; Blaustein, M.P. Ouabain-Digoxin antagonism in rat arteries and neurones. J. Physiol. 2014, 592, 941–969. [Google Scholar] [CrossRef] [PubMed]
- Xie, Z.; Xie, J. The Na/K-ATPase-mediated signal transduction as a target for new drug development. Front. Biosci. 2005, 10, 3100–3109. [Google Scholar] [CrossRef] [PubMed]
- Ahmed, A.; Pitt, B.; Rahimtoola, S.H.; Waagstein, F.; White, M.; Love, T.E.; Braunwald, E. Effects of digoxin at low serum concentrations on mortality and hospitalization in heart failure: A propensity-matched study of the dig trial. Int. J. Cardiol. 2008, 123, 138–146. [Google Scholar] [CrossRef] [PubMed]
- Pasdois, P.; Quinlan, C.L.; Rissa, A.; Tariosse, L.; Vinassa, B.; Costa, A.D.; Pierre, S.V.; Dos Santos, P.; Garlid, K.D. Ouabain protects rat hearts against ischemia-reperfusion injury via pathway involving Src kinase, mitoKATP, and ROS. Am. J. Physiol. Heart Circ. Physiol. 2007, 292, H1470–H1478. [Google Scholar] [CrossRef] [PubMed]
- D’Urso, G.; Frascarelli, S.; Zucchi, R.; Biver, T.; Montali, U. Cardioprotection by ouabain and digoxin in perfused rat hearts. J. Cardiovasc. Pharmacol. 2008, 52, 333–337. [Google Scholar] [CrossRef] [PubMed]
- Pierre, S.V.; Yang, C.; Yuan, Z.; Seminerio, J.; Mouas, C.; Garlid, K.D.; Dos-Santos, P.; Xie, Z. Ouabain triggers preconditioning through activation of the Na+,K+-ATPase signaling cascade in rat hearts. Cardiovasc. Res. 2007, 73, 488–496. [Google Scholar] [CrossRef] [PubMed]
- McConkey, D.J.; Lin, Y.; Nutt, L.K.; Ozel, H.Z.; Newman, R.A. Cardiac glycosides stimulate Ca2+ increases and apoptosis in androgen-independent, metastatic human prostate adenocarcinoma cells. Cancer Res. 2000, 60, 3807–3812. [Google Scholar] [PubMed]
- Wang, Y.; Qiu, Q.; Shen, J.J.; Li, D.D.; Jiang, X.J.; Si, S.Y.; Shao, R.G.; Wang, Z. Cardiac glycosides induce autophagy in human non-small cell lung cancer cells through regulation of dual signaling pathways. Int. J. Biochem. Cell Biol. 2012, 44, 1813–1824. [Google Scholar] [CrossRef] [PubMed]
- Prassas, I.; Karagiannis, G.S.; Batruch, I.; Dimitromanolakis, A.; Datti, A.; Diamandis, E.P. Digitoxin-Induced cytotoxicity in cancer cells is mediated through distinct kinase and interferon signaling networks. Mol. Cancer Ther. 2011, 10, 2083–2093. [Google Scholar] [CrossRef] [PubMed]
- Blanco, G.; Wallace, D.P. Novel role of ouabain as a cystogenic factor in autosomal dominant polycystic kidney disease. Am. J. Physiol. Renal Physiol. 2013, 305, F797–F812. [Google Scholar] [CrossRef] [PubMed]
- Jansson, K.; Magenheimer, B.S.; Maser, R.L.; Calvet, J.P.; Blanco, G. Overexpression of the polycystin-1 C-tail enhances sensitivity of M-1 cells to ouabain. J. Membr. Biol. 2013, 246, 581–590. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Liang, M.; Liu, L.; Malhotra, D.; Xie, Z.; Shapiro, J.I. Ouabain-Induced endocytosis of the plasmalemmal Na/K-ATPase in LLC-PK1 cells requires caveolin-1. Kidney Int. 2005, 67, 1844–1854. [Google Scholar] [CrossRef] [PubMed]
- Gupta, S.; Yan, Y.; Malhotra, D.; Liu, J.; Xie, Z.; Najjar, S.M.; Shapiro, J.I. Ouabain and insulin induce sodium pump endocytosis in renal epithelium. Hypertension 2012, 59, 665–672. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Xie, Z.J. The sodium pump and cardiotonic steroids-induced signal transduction protein kinases and calcium-signaling microdomain in regulation of transporter trafficking. Biochim. Biophys. Acta 2010, 1802, 1237–1245. [Google Scholar] [CrossRef] [PubMed]
- Digby, G.J.; Lober, R.M.; Sethi, P.R.; Lambert, N.A. Some G protein heterotrimers physically dissociate in living cells. Proc. Natl. Acad. Sci. USA 2006, 103, 17789–17794. [Google Scholar] [CrossRef] [PubMed]
- Rubenstein, L.A.; Zauhar, R.J.; Lanzara, R.G. Molecular dynamics of a biophysical model for β2-adrenergic and G protein-coupled receptor activation. J. Mol. Graph. Model. 2006, 25, 396–409. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Z.; Li, Z.; Tian, J.; Jiang, W.; Wang, Y.; Zhang, X.; Li, Z.; You, Q.; Shapiro, J.I.; Si, S.; et al. Identification of hydroxyxanthones as Na/K-ATPase ligands. Mol. Pharmacol. 2010, 77, 961–967. [Google Scholar] [CrossRef] [PubMed]
- Rhee, S.G. H2O2, a necessary evil for cell signaling. Science 2006, 312, 1882–1883. [Google Scholar] [CrossRef] [PubMed]
- Sun, G.; Kemble, D.J. To C or not to C: Direct and indirect redox regulation of Src protein tyrosine kinase. Cell Cycle 2009, 8, 2353–2355. [Google Scholar] [CrossRef] [PubMed]
- MacKay, C.E.; Knock, G.A. Control of vascular smooth muscle function by Src-family kinases and reactive oxygen species in health and disease. J. Physiol. 2015, 593, 3815–3828. [Google Scholar] [CrossRef] [PubMed]
- Sedeek, M.; Nasrallah, R.; Touyz, R.M.; Hebert, R.L. Nadph oxidases, reactive oxygen species, and the kidney: Friend and foe. J. Am. Soc. Nephrol. 2013, 24, 1512–1518. [Google Scholar] [CrossRef] [PubMed]
- Madamanchi, N.R.; Runge, M.S. Redox signaling in cardiovascular health and disease. Free Radic. Biol. Med. 2013, 61, 473–501. [Google Scholar] [CrossRef] [PubMed]
- Corcoran, A.; Cotter, T.G. Redox regulation of protein kinases. FEBS J. 2013, 280, 1944–1965. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Tian, J.; Haas, M.; Shapiro, J.I.; Askari, A.; Xie, Z. Ouabain interaction with cardiac Na+/K+-ATPase initiates signal cascades independent of changes in intracellular Na+ and Ca2+ concentrations. J. Biol. Chem. 2000, 275, 27838–27844. [Google Scholar] [PubMed]
- Xie, Z.J.; Wang, Y.H.; Askari, A.; Huang, W.H.; Klaunig, J.E.; Askari, A. Studies on the specificity of the effects of oxygen metabolites on cardiac sodium pump. J. Mol. Cell. Cardiol. 1990, 22, 911–920. [Google Scholar] [CrossRef]
- Huang, W.H.; Wang, Y.; Askari, A. (Na+ + K+)-ATPase: Inactivation and degradation induced by oxygen radicals. Int. J. Biochem. 1992, 24, 621–626. [Google Scholar] [PubMed]
- Yan, Y.; Shapiro, A.P.; Mopidevi, B.R.; Chaudhry, M.A.; Maxwell, K.; Haller, S.T.; Drummond, C.A.; Kennedy, D.J.; Tian, J.; Malhotra, D.; et al. Protein carbonylation of an amino acid residue of the Na/K-ATPase α1 subunit determines Na/K-ATPase signaling and sodium transport in renal proximal tubular cells. J. Am. Heart Assoc. 2016, 5, e003675. [Google Scholar] [CrossRef] [PubMed]
- Liu, C.C.; Garcia, A.; Mahmmoud, Y.A.; Hamilton, E.J.; Galougahi, K.K.; Fry, N.A.; Figtree, G.A.; Cornelius, F.; Clarke, R.J.; Rasmussen, H.H. Susceptibility of β1 Na+-K+ pump subunit to glutathionylation and oxidative inhibition depends on conformational state of pump. J. Biol. Chem. 2012, 287, 12353–12364. [Google Scholar] [CrossRef] [PubMed]
- Bundgaard, H.; Liu, C.C.; Garcia, A.; Hamilton, E.J.; Huang, Y.; Chia, K.K.; Hunyor, S.N.; Figtree, G.A.; Rasmussen, H.H. β(3) Adrenergic stimulation of the cardiac Na+-K+ pump by reversal of an inhibitory oxidative modification. Circulation 2010, 122, 2699–2708. [Google Scholar] [CrossRef] [PubMed]
- White, C.N.; Liu, C.C.; Garcia, A.; Hamilton, E.J.; Chia, K.K.; Figtree, G.A.; Rasmussen, H.H. Activation of camp-dependent signaling induces oxidative modification of the cardiac Na+-K+ pump and inhibits its activity. J. Biol. Chem. 2010, 285, 13712–13720. [Google Scholar] [CrossRef] [PubMed]
- Goldshleger, R.; Karlish, S.J. Fe-Catalyzed cleavage of the α subunit of Na/K-ATPase: Evidence for conformation-sensitive interactions between cytoplasmic domains. Proc. Natl. Acad. Sci. USA 1997, 94, 9596–9601. [Google Scholar] [CrossRef] [PubMed]
- Xie, J.X.; Li, X.; Xie, Z. Regulation of renal function and structure by the signaling Na/K-ATPase. IUBMB Life 2013, 65, 991–998. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Tian, J.; Chaudhry, M.; Maxwell, K.; Yan, Y.; Wang, X.; Shah, P.T.; Khawaja, A.A.; Martin, R.; Robinette, T.J.; et al. Attenuation of Na/K-ATPase mediated oxidant amplification with pnaktide ameliorates experimental uremic cardiomyopathy. Sci. Rep. 2016, 6, 34592. [Google Scholar] [CrossRef] [PubMed]
- Sodhi, K.; Srikanthan, K.; Goguet-Rubio, P.; Nichols, A.; Mallick, A.; Nawab, A.; Martin, R.; Shah, P.T.; Chaudhry, M. pNaKtide Attenuates steatohepatitis and atherosclerosis by blocking Na/K-ATPase/ROS amplification in C57Bl6 and ApoE knockout mice fed a western diet. Sci. Rep. 2017, 7, 193. [Google Scholar] [CrossRef] [PubMed]
- Garver, W.S.; Hossain, G.S.; Winscott, M.M.; Heidenreich, R.A. The Npc1 mutation causes an altered expression of caveolin-1, annexin II and protein kinases and phosphorylation of caveolin-1 and annexin II in murine livers. Biochim. Biophys. Acta 1999, 1453, 193–206. [Google Scholar] [CrossRef]
- Lockwich, T.P.; Liu, X.; Singh, B.B.; Jadlowiec, J.; Weiland, S.; Ambudkar, I.S. Assembly of Trp1 in a signaling complex associated with caveolin-scaffolding lipid raft domains. J. Biol. Chem. 2000, 275, 11934–11942. [Google Scholar] [CrossRef] [PubMed]
- Cai, T.; Wang, H.; Chen, Y.; Liu, L.; Gunning, W.T.; Quintas, L.E.; Xie, Z.J. Regulation of caveolin-1 membrane trafficking by the Na/K-ATPase. J. Cell Biol. 2008, 182, 1153–1169. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.; Cai, T.; Wang, H.; Li, Z.; Loreaux, E.; Lingrel, J.B.; Xie, Z. Regulation of intracellular cholesterol distribution by Na/K-ATPase. J. Biol. Chem. 2009, 284, 14881–14890. [Google Scholar] [CrossRef] [PubMed]
- Liang, M.; Tian, J.; Liu, L.; Pierre, S.; Liu, J.; Shapiro, J.; Xie, Z.J. Identification of a pool of non-pumping Na/K-ATPase. J. Biol. Chem. 2007, 282, 10585–10593. [Google Scholar] [CrossRef] [PubMed]
- Liang, M.; Cai, T.; Tian, J.; Qu, W.; Xie, Z.J. Functional characterization of Src-interacting Na/K-ATPase using RNA interference assay. J. Biol. Chem. 2006, 281, 19709–19719. [Google Scholar] [CrossRef] [PubMed]
- Fujimoto, T.; Nakade, S.; Miyawaki, A.; Mikoshiba, K.; Ogawa, K. Localization of inositol 1,4,5-trisphosphate receptor-like protein in plasmalemmal caveolae. J. Cell Biol. 1992, 119, 1507–1513. [Google Scholar] [CrossRef] [PubMed]
- Lisanti, M.P.; Scherer, P.E.; Vidugiriene, J.; Tang, Z.; Hermanowski-Vosatka, A.; Tu, Y.H.; Cook, R.F.; Sargiacomo, M. Characterization of caveolin-rich membrane domains isolated from an endothelial-rich source: Implications for human disease. J. Cell Biol. 1994, 126, 111–126. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.; Li, X.; Ye, Q.; Tian, J.; Jing, R.; Xie, Z. Regulation of α1 Na/K-ATPase expression by cholesterol. J. Biol. Chem. 2011, 286, 15517–15524. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.; Cai, T.; Yang, C.; Turner, D.A.; Giovannucci, D.R.; Xie, Z. Regulation of inositol 1,4,5-trisphosphate receptor-mediated calcium release by the Na/K-ATPase in cultured renal epithelial cells. J. Biol. Chem. 2008, 283, 1128–1136. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Schematic presentation of Albers–Post reaction mechanism.
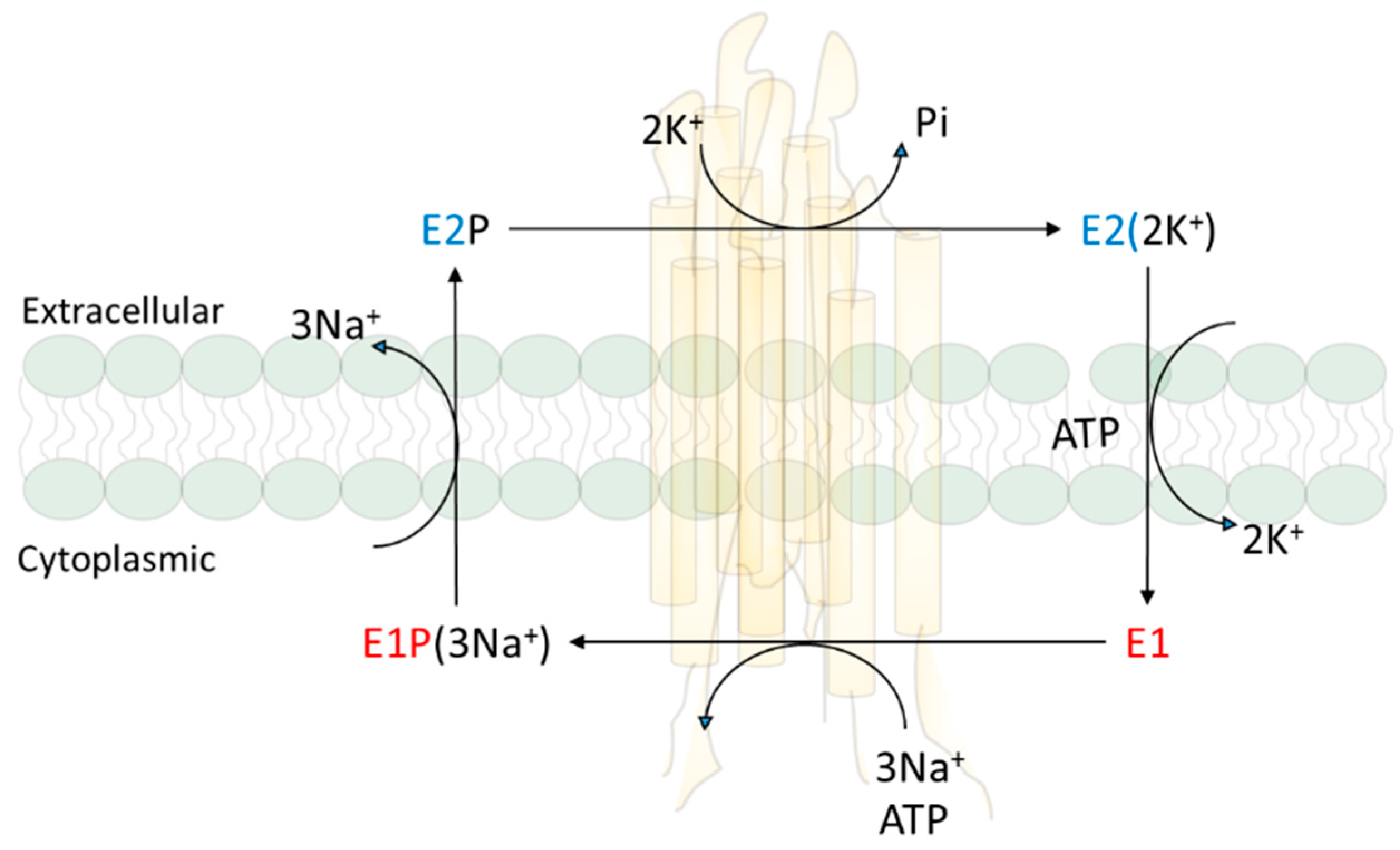
Figure 2.
Schematic presentation of NKA-mediated signal transduction through direct protein interactions. EGFR, EGF receptor; PLC, phospholipase C; PKC, protein kinase.
Figure 2.
Schematic presentation of NKA-mediated signal transduction through direct protein interactions. EGFR, EGF receptor; PLC, phospholipase C; PKC, protein kinase.

Figure 3.
Comparison of NaKtide sequences among different species in α1, α2 and α3 subunits. The amino acids being different from mammalian α1 are marked in red. Data are from NCBI data base.
Figure 3.
Comparison of NaKtide sequences among different species in α1, α2 and α3 subunits. The amino acids being different from mammalian α1 are marked in red. Data are from NCBI data base.
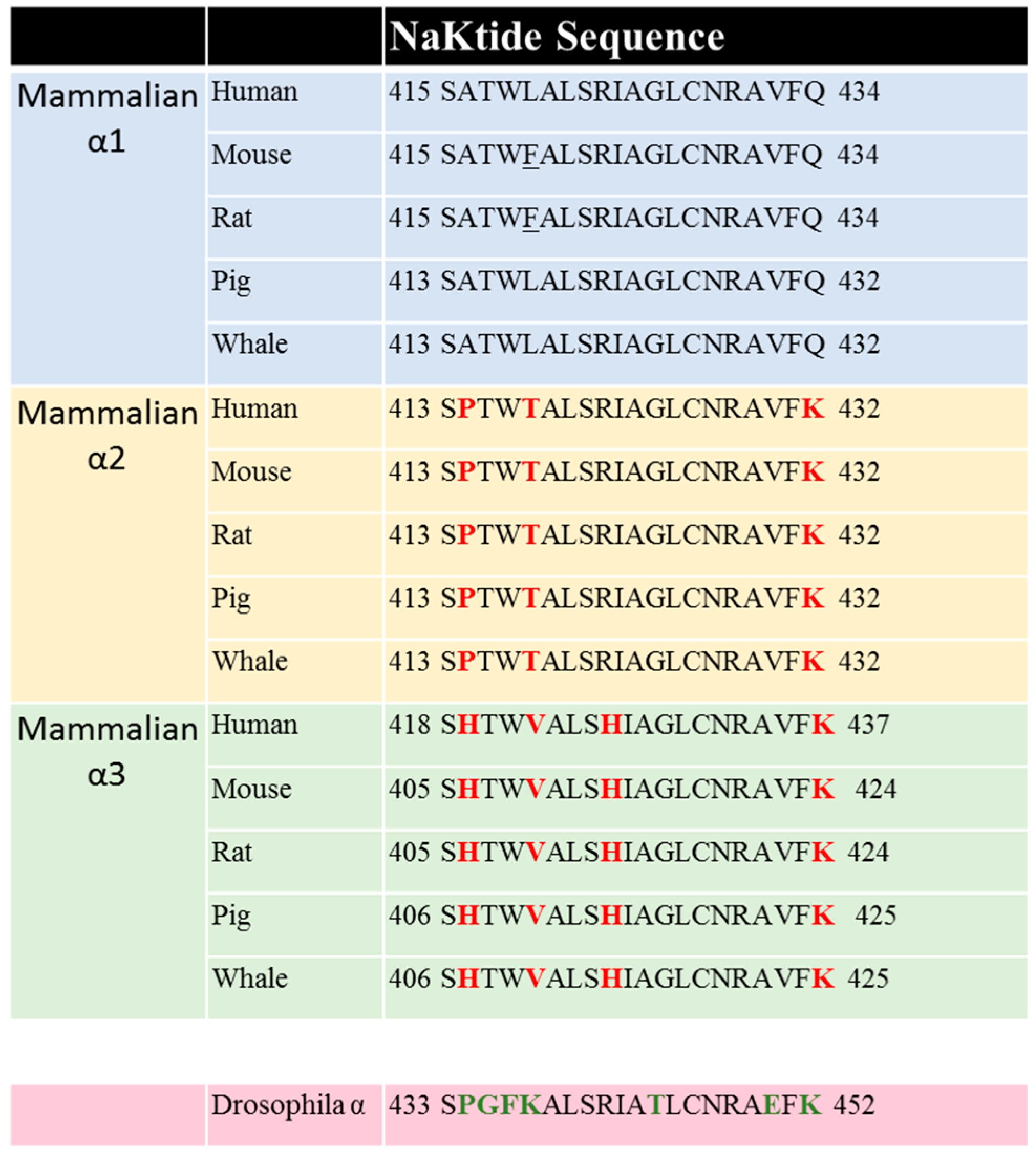
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Cui, X.; Xie, Z. Protein Interaction and Na/K-ATPase-Mediated Signal Transduction. Molecules 2017, 22, 990. https://doi.org/10.3390/molecules22060990
AMA Style
Cui X, Xie Z. Protein Interaction and Na/K-ATPase-Mediated Signal Transduction. Molecules. 2017; 22(6):990. https://doi.org/10.3390/molecules22060990
Chicago/Turabian StyleCui, Xiaoyu, and Zijian Xie. 2017. "Protein Interaction and Na/K-ATPase-Mediated Signal Transduction" Molecules 22, no. 6: 990. https://doi.org/10.3390/molecules22060990