
1,4-Naphthoquinone Triggers Nematode Lethality by Inducing Oxidative Stress and Activating Insulin/IGF Signaling Pathway in Caenorhabditis elegans

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results and Discussion
2.1. Chemical Screening of Nematocidal Compounds
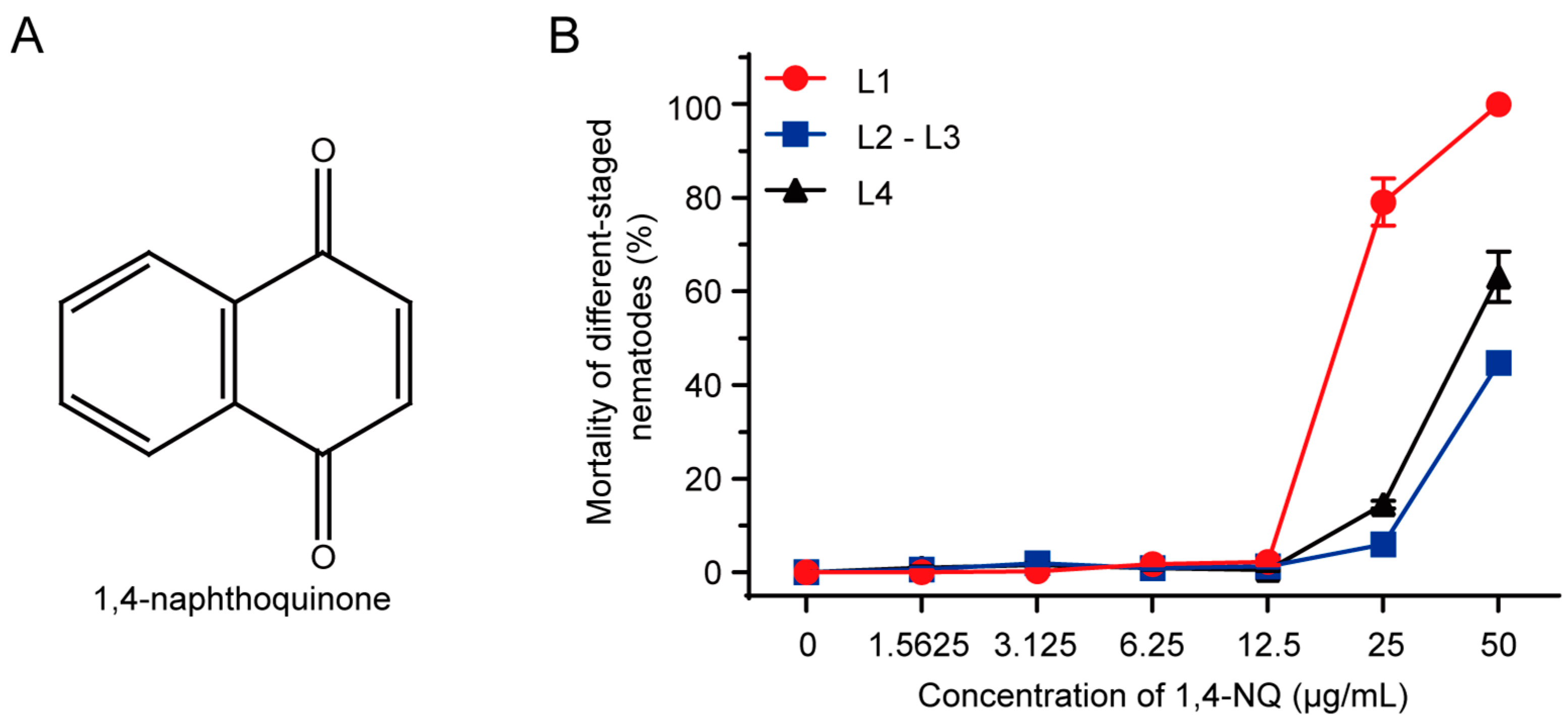
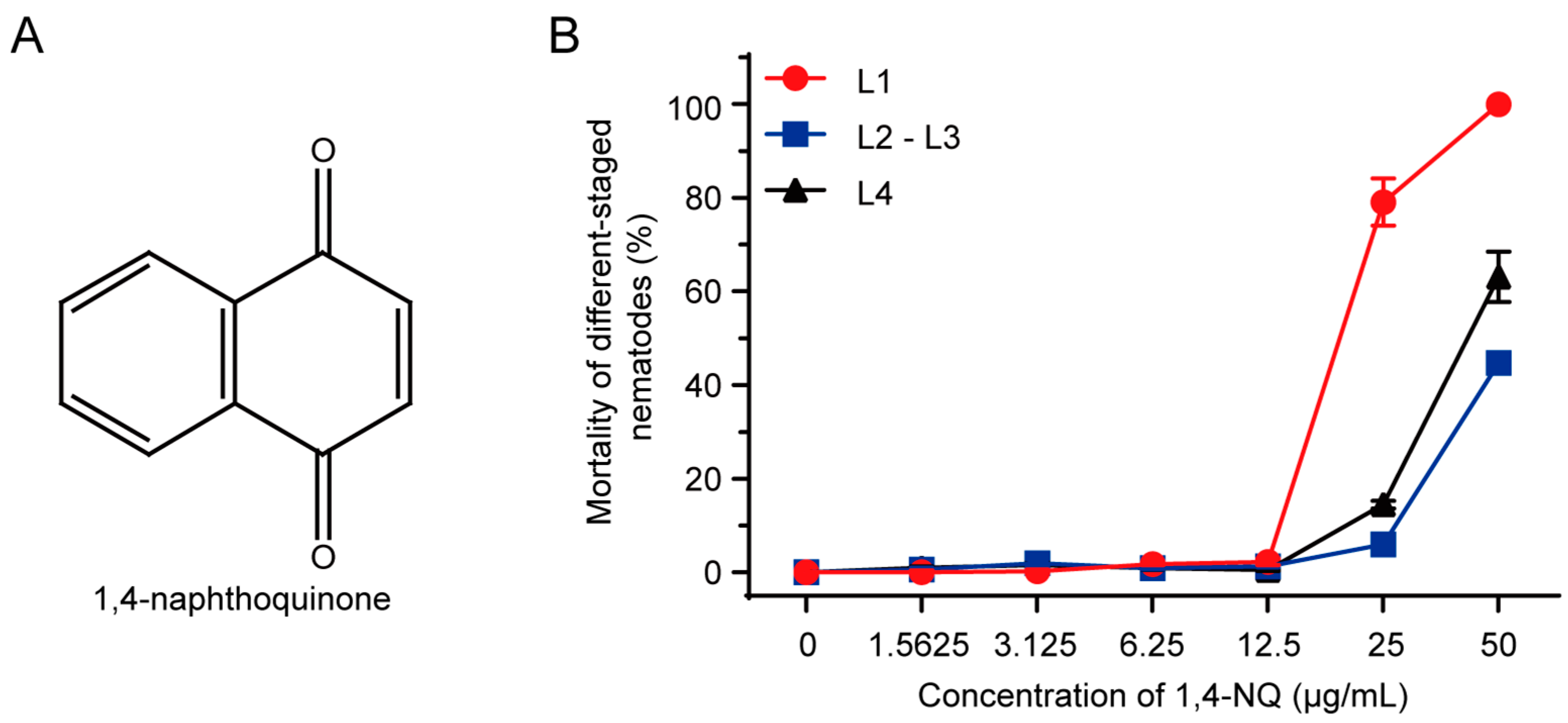
2.2. N2 L1 Was the Most Sensitive to 1,4-NQ
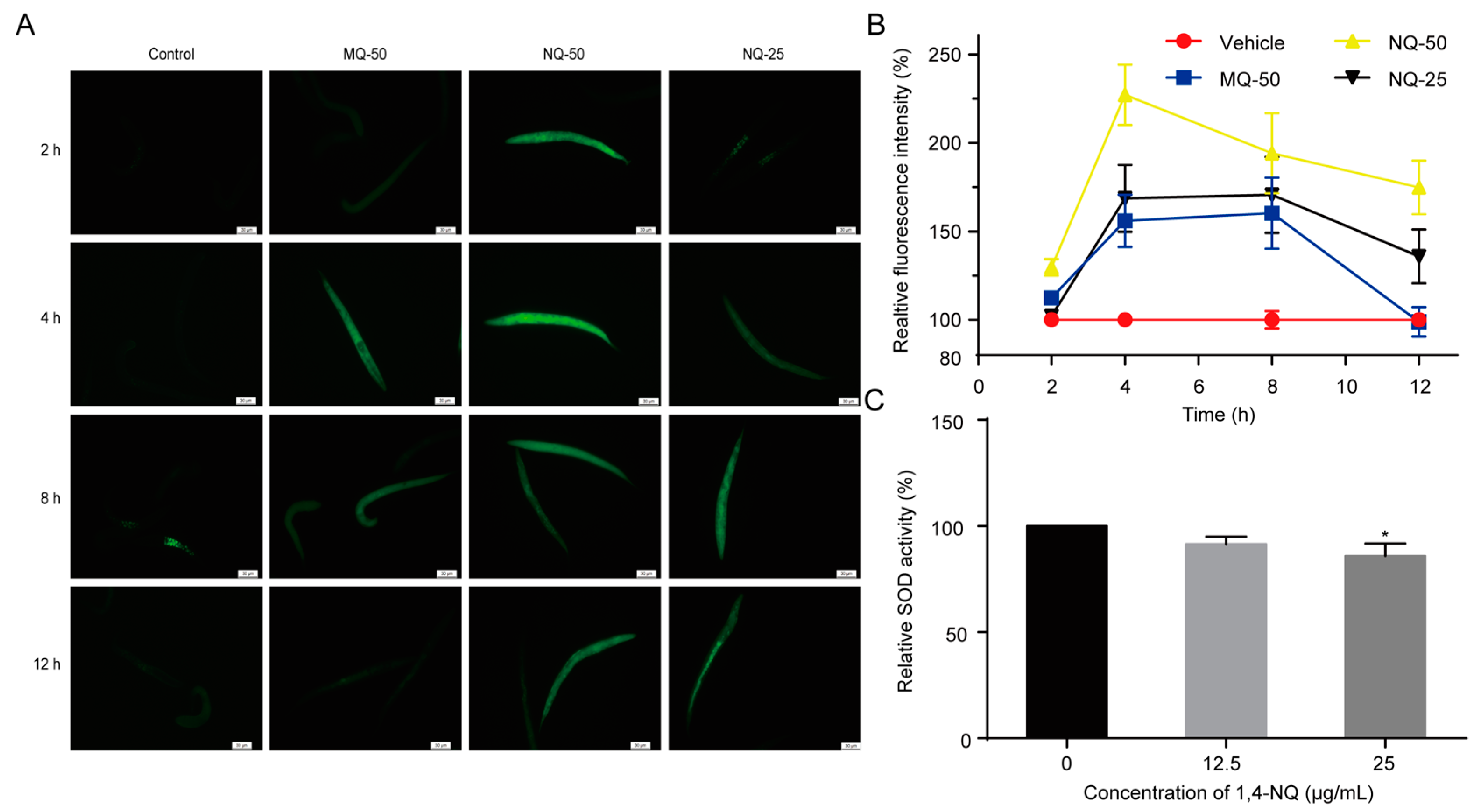
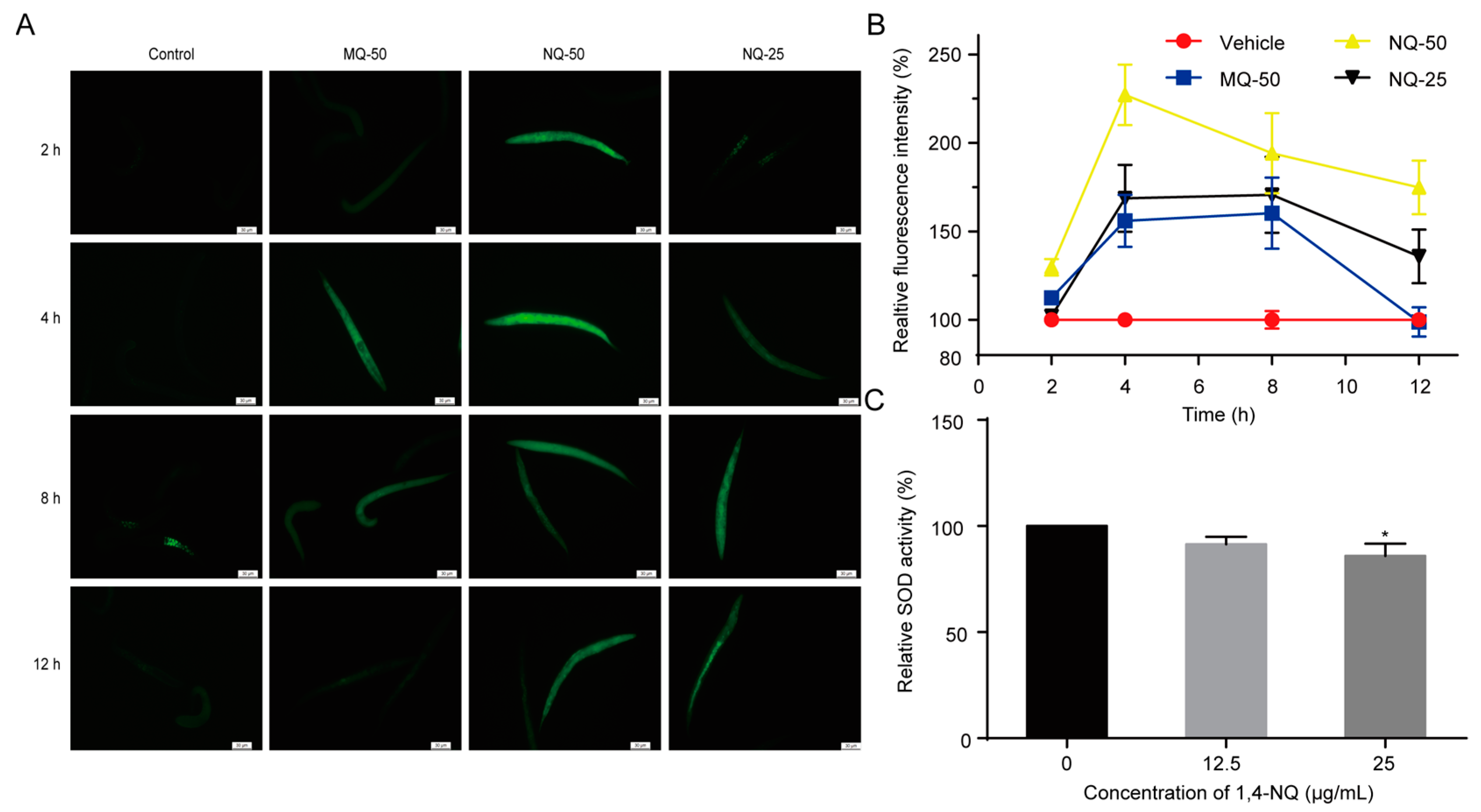
2.3. 1,4-NQ Stimulates ROS Production and Suppresses Elimination of ROS in C. elegans
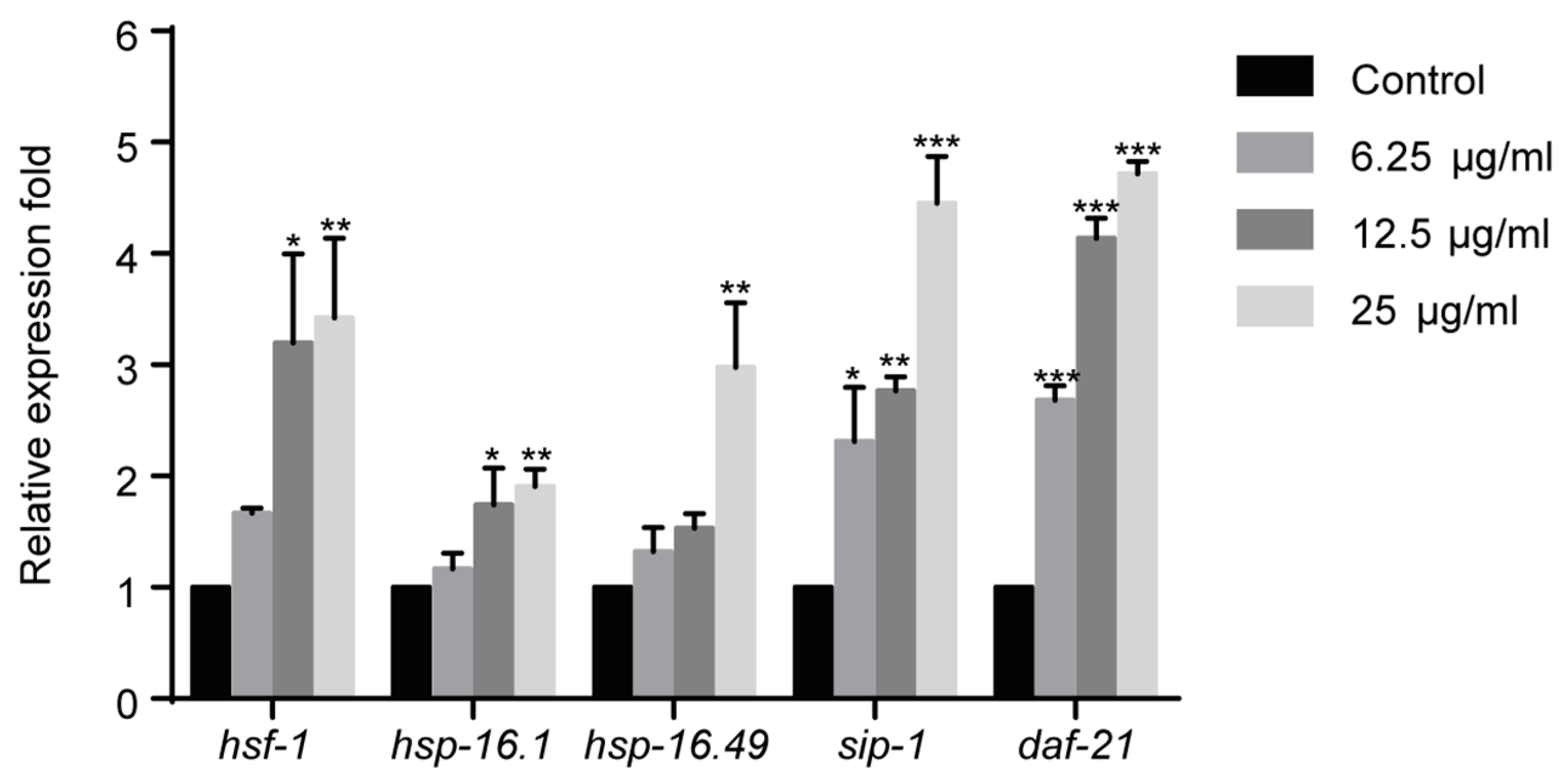
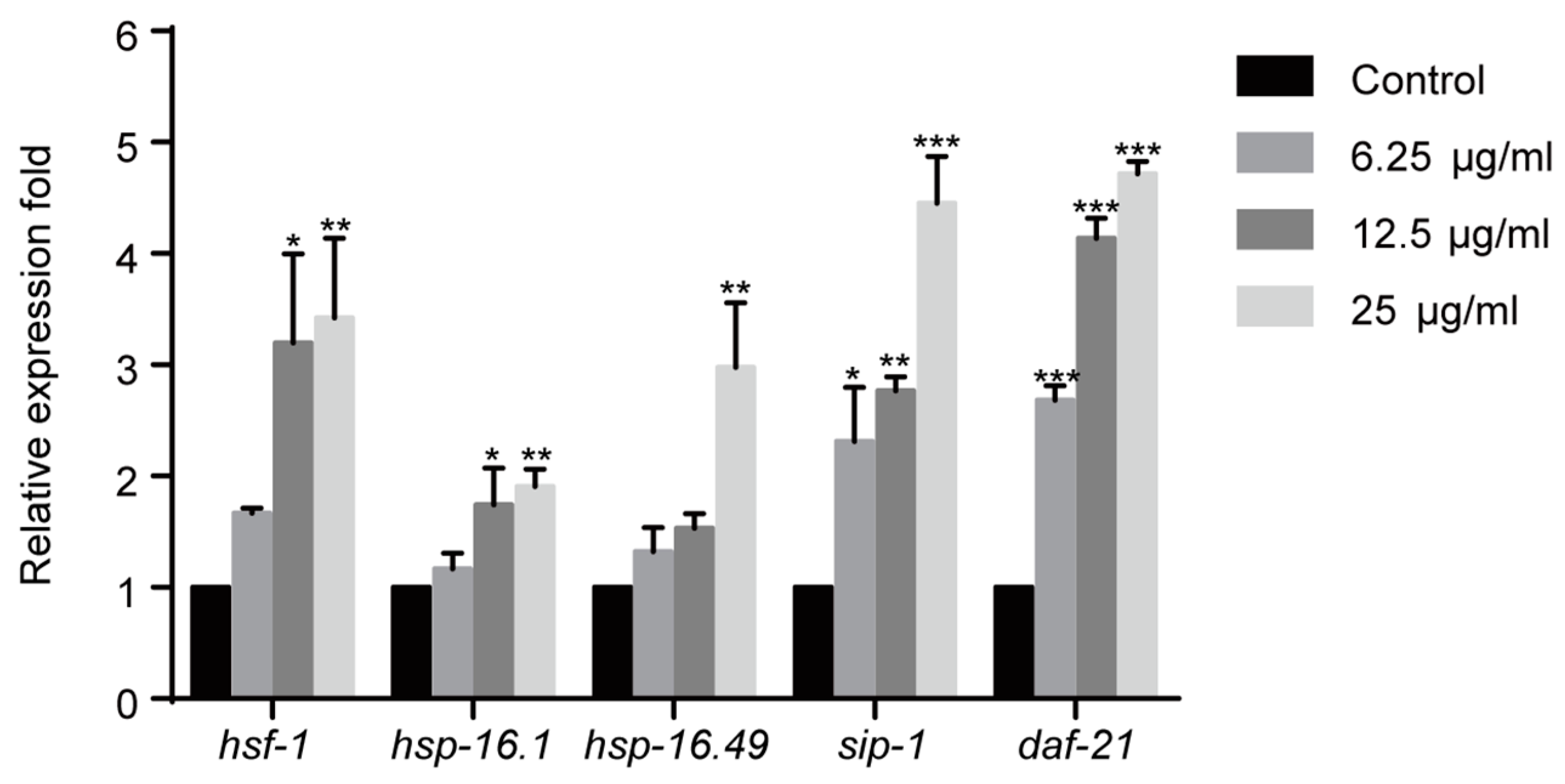
2.4. 1,4-NQ Activates HSF-1 Signaling Pathway in C. elegans
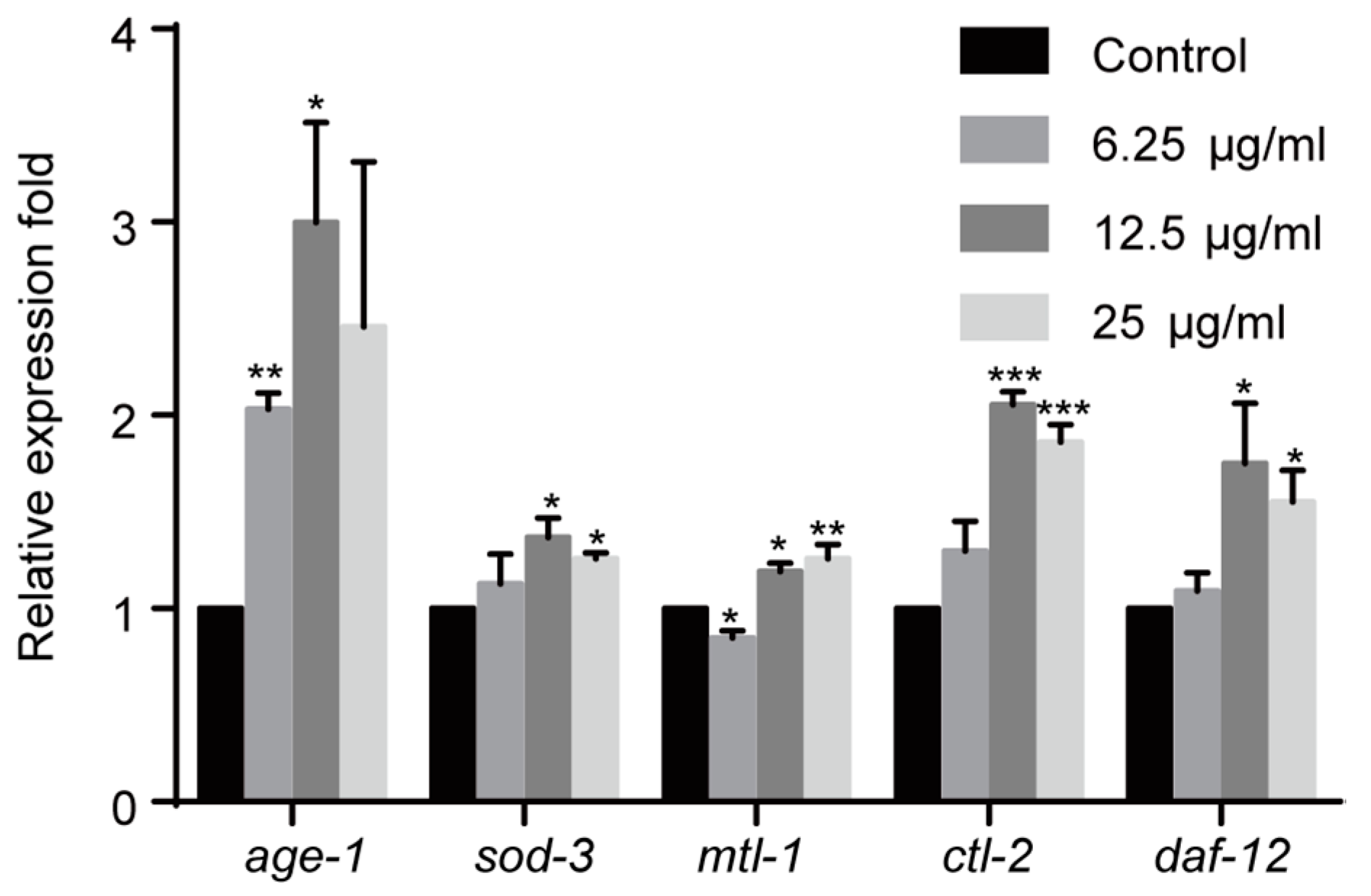
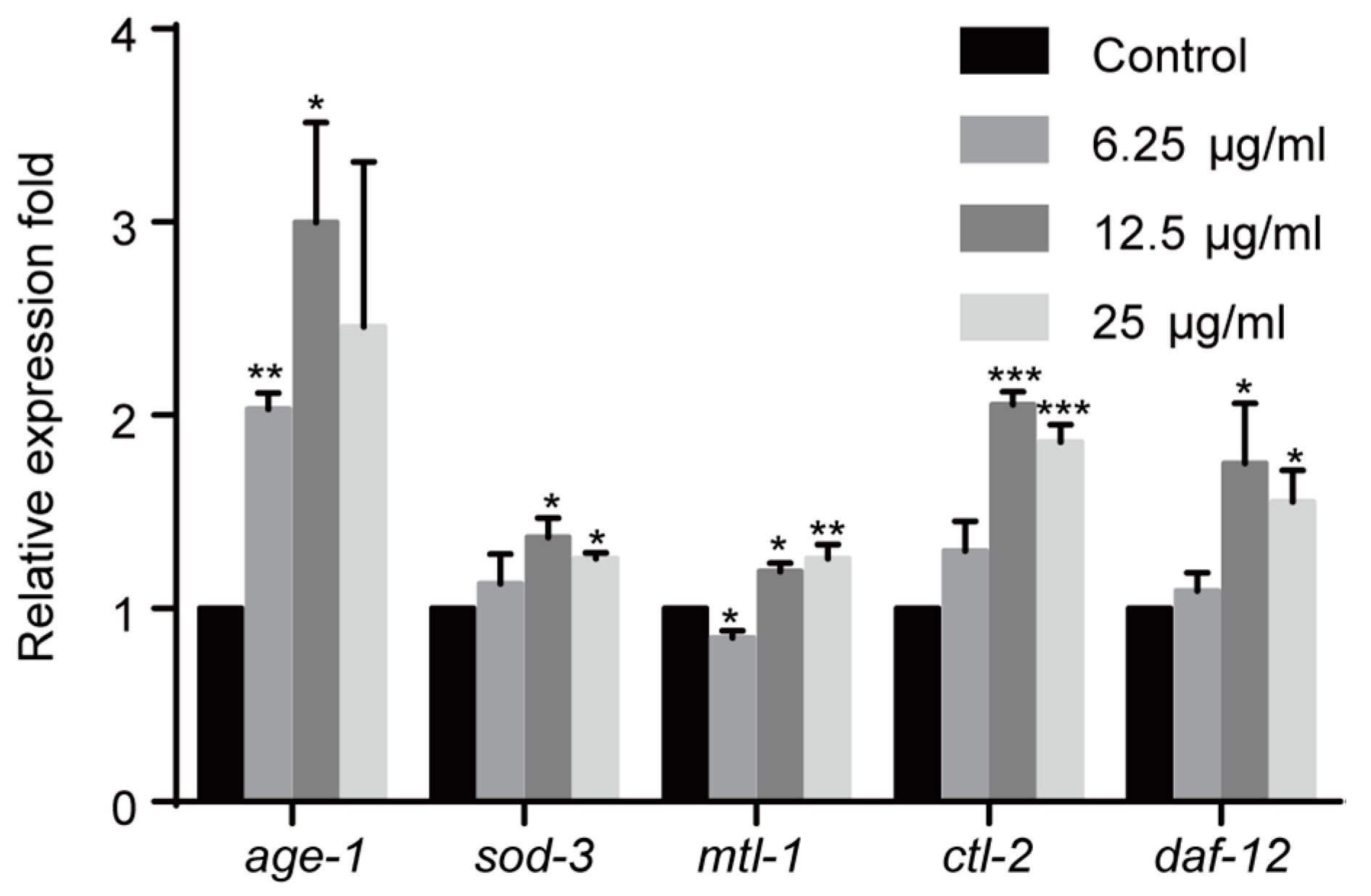
2.5. Effect of 1,4-NQ on IIS Pathway Related Gene Expressions in C. elegans
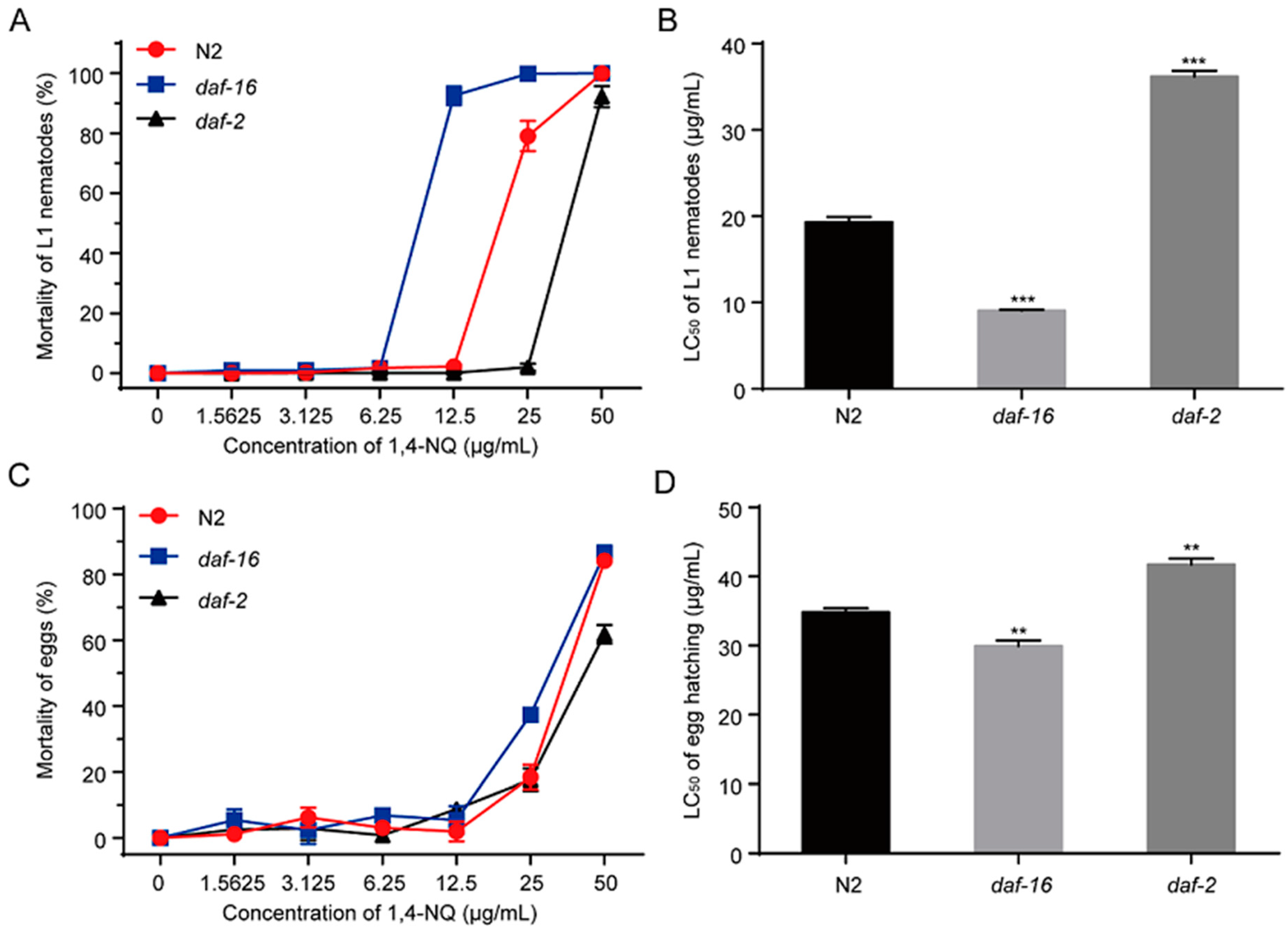
2.6. Decreased Sensitivity of daf-2 Mutants and Increased Sensitivity of daf-16 Mutants to 1,4-NQ
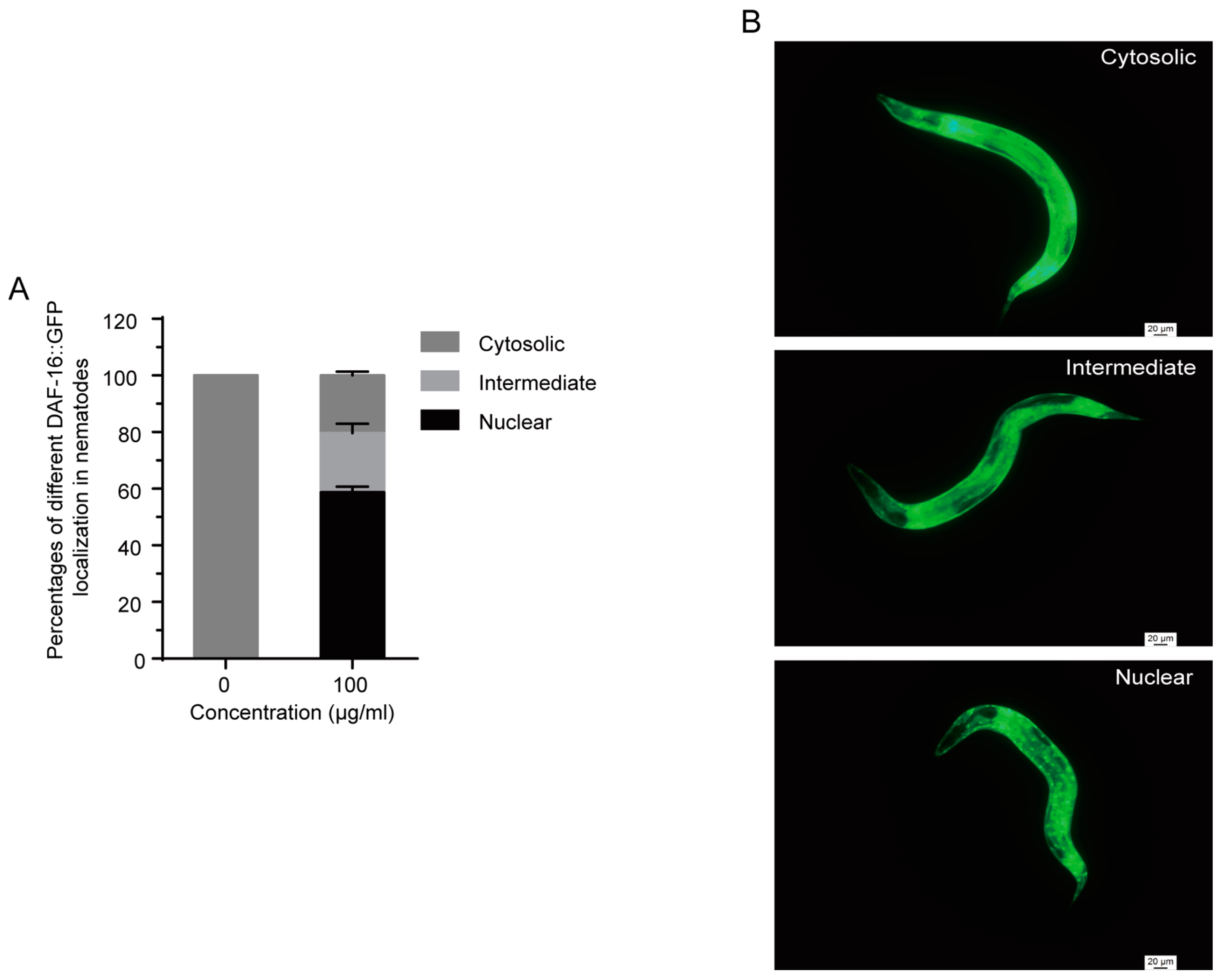
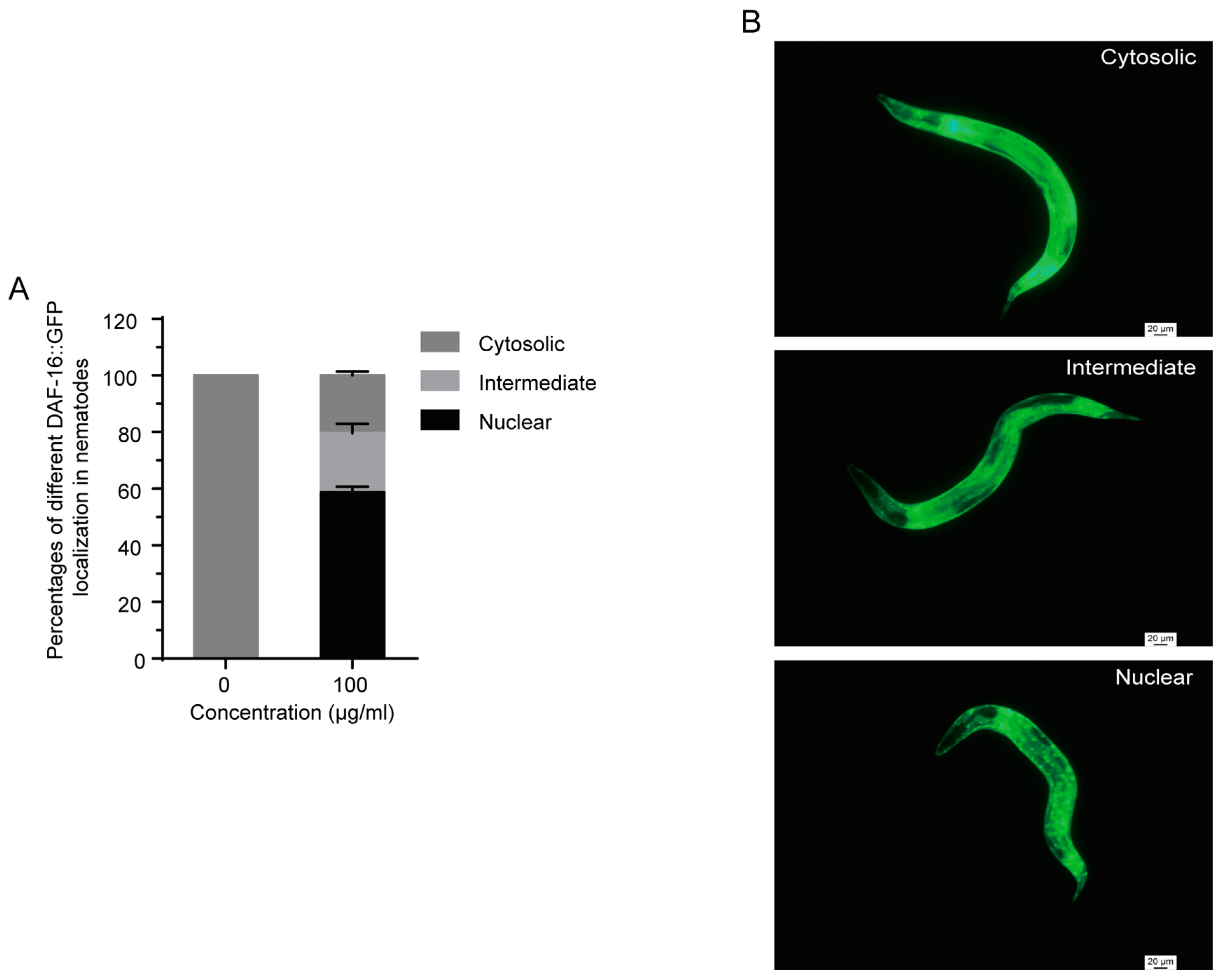
2.7. 1,4-NQ Stimulates DAF-16::GFP Localization in C. elegans
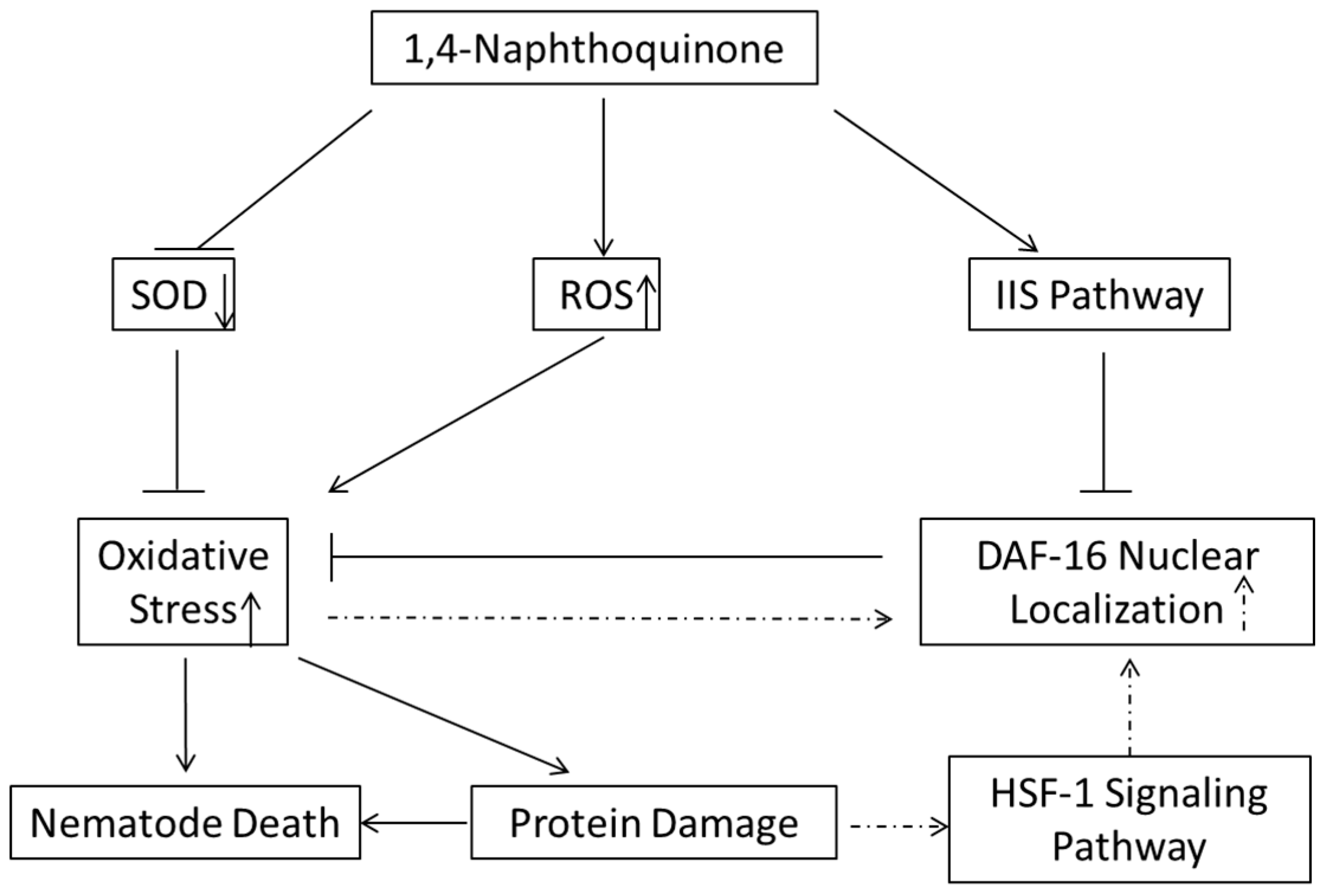
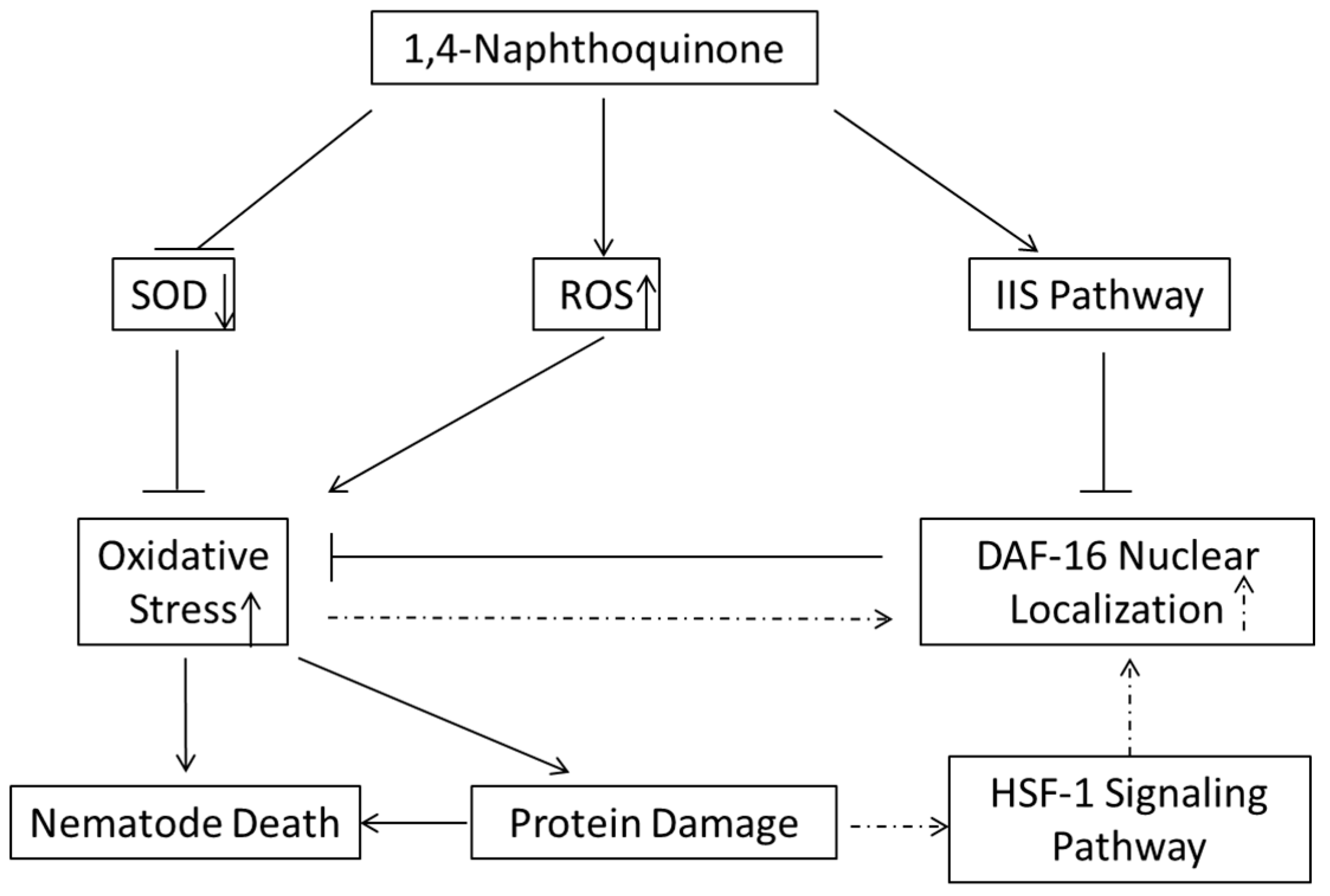
2.8. Discussion
3. Materials and Methods
3.1. Chemical Compounds
3.2. Nematode Strains and Culture Methods
3.3. Chemical Screening
3.4. ROS Measurement
3.5. Measurement of SOD Activity
3.6. Reverse Transcription-Polymerase Chain Reaction (RT--PCR) Analyses
3.7. Mutant Resistance Assay
3.8. DAF-16::GFP Nuclear Localization
3.9. Statistical Analysis
Supplementary Materials
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Fuller, V.L.; Lilley, C.J.; Urwin, P.E. Nematode resistance. New Phytol. 2008, 180, 27–44. [Google Scholar] [CrossRef] [PubMed]
- Abad, P.; Gouzy, J.; Aury, J.M.; Castagnone-Sereno, P.; Danchin, E.J.; Deleury, E.; Perfus-Barbeoch, L.; Anthouard, V.; Artiguenave, F.; Blok, V.C.; et al. . Genome sequence of the metazoan plant-parasitic nematode Meloidogyne incognita. Nat. Biotechnol. 2008, 26, 909–915. [Google Scholar] [CrossRef] [PubMed]
- Rich, J.R.; Dunn, R.A.; Noling, J.W. Nematicides: Past and present uses. Nematology 2004, 21, 1179–1200. [Google Scholar]
- Cabrera, J.A.; Menjivar, R.D.; Dababat, A.A.; Sikora, R.A. Properties and nematicide performance of avermectins. J. Phytopathol. 2013, 161, 65–69. [Google Scholar] [CrossRef]
- Ntalli, N.G.; Menkissoglu-Spiroudi, U. Pesticides of Botanical Origin: A Promising Tool in Plant Protection. In Pesticides—Formulations, Effects, Fate, 1st ed.; Stoytcheva, M., Ed.; INTECH Open Access Publisher: Janeza Trdine, Rijeka, Croatia, 2011. [Google Scholar]
- Ntalli, N.G.; Caboni, P. Botanical nematicides: A review. J. Agric. Food Chem. 2012, 60, 9929–9940. [Google Scholar] [CrossRef] [PubMed]
- Akhtar, Y.; Yeoung, Y.R.; Isman, M.B. Comparative bioactivity of selected extracts from Meliaceae and some commercial botanical insecticides against two noctuid caterpillars, Trichoplusia ni and Pseudaletia unipuncta. Phytochem. Rev. 2008, 7, 77–88. [Google Scholar] [CrossRef]
- Flamini, G. Acaricides of natural origin, personal experiences and review of literature (1900–2001). Stud. Nat. Prod. Chem. 2003, 28, 381–451. [Google Scholar]
- Oka, Y.; Tkachi, N.; Shuker, S.; Yerumiyahu, U. Enhanced nematicidal activity of organic and inorganic ammonia-releasing amendments by Azadirachta indica extracts. J. Nematol. 2007, 39, 9–16. [Google Scholar] [PubMed]
- Chitwood, D.J. Phytochemical based strategies for nematode control. Annu. Rev. Phytopathol. 2002, 40, 221–249. [Google Scholar] [CrossRef] [PubMed]
- Isman, M.B. Botanical insecticides: For richer, for poorer. Pest. Manag. Sci. 2008, 64, 8–11. [Google Scholar] [CrossRef] [PubMed]
- Sijmons, P.C.; Atkinson, H.J.; Wyss, U. Parasitic strategies of root nematodes and associated host cell responses. Annu. Rev. Phytopathol. 1994, 32, 235–259. [Google Scholar] [CrossRef]
- Burns, A.R.; Luciani, G.M.; Musso, G.; Bagg, R.; Yeo, M.; Zhang, Y.; Rajendran, L.; Glavin, J.; Hunter, R.; Redman, E.; et al. Caenorhabditis elegans is a useful model for anthelmintic discovery. Nat. Commun. 2015, 6, 7485. [Google Scholar] [CrossRef] [PubMed]
- Anastassopoulou, C.G.; Fuchs, B.B.; Mylonakis, E. Caenorhabditis elegans-based model systems for antifungal drug discovery. Curr. Pharm. Des. 2011, 17, 1225–1233. [Google Scholar] [CrossRef] [PubMed]
- Leung, C.K.; Wang, Y.; Malany, S.; Deonarine, A.; Nguyen, K.; Vasile, S.; Choe, K.P. An ultra high-throughput, whole-animal screen for small molecule modulators of a specific genetic pathway in Caenorhabditis elegans. PLoS ONE 2013, 8, e62166. [Google Scholar] [CrossRef] [PubMed]
- Thor, H.; Smith, M.T.; Hartzell, P.; Bellomo, G.; Jewell, S.A.; Orrenius, S. The metabolism of menadione (2-methyl-1,4-naphthoquinone) by isolated hepatocytes. A study of the implications of oxidative stress in intact cells. J. Biol. Chem. 1982, 257, 12419–12425. [Google Scholar] [PubMed]
- Klaus, V.; Hartmann, T.; Gambini, J.; Graf, P.; Stahl, W.; Hartwig, A.; Klotz, L.O. 1,4-Naphthoquinones as inducers of oxidative damage and stress signaling in HaCaT human keratinocytes. Arch. Biochem. Biophys. 2010, 496, 93–100. [Google Scholar] [CrossRef] [PubMed]
- Inbaraj, J.J.; Chignell, C.F. Cytotoxic action of juglone and plumbagin: A mechanistic study using HaCaT keratinocytes. Chem. Res. Toxicol. 2004, 17, 55–62. [Google Scholar] [CrossRef] [PubMed]
- George, E.J.; Neufeld, R.D. Degradation of fluorene in soil by fungus Phanerochaete chrysosporium. Biotechnol. Bioeng. 1989, 33, 1306–1310. [Google Scholar] [CrossRef] [PubMed]
- Circu, M.L.; Aw, T.Y. Reactive oxygen species, cellular redox systems, and apoptosis. Free Radic. Biol. Med. 2010, 48, 749–762. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Jie, G.; Zhang, J.; Zhao, B. Significant longevity-extending effects of EGCG on Caenorhabditis elegans under stress. Free Radic. Biol. Med. 2009, 46, 414–421. [Google Scholar] [CrossRef] [PubMed]
- Honda, Y.; Honda, S. The daf-2 gene network for longevity regulates oxidative stress resistance and Mn-superoxide dismutase gene expression in Caenorhabditis elegans. FASEB J. 1999, 13, 1385–1393. [Google Scholar] [PubMed]
- Li, Y.; Yu, S.; Wu, Q.; Tang, M.; Pu, Y.; Wang, D. Chronic Al2O3-nanoparticle exposure causes neurotoxic effects on locomotion behaviors by inducing severe ROS production and disruption of ROS defense mechanisms in nematode Caenorhabditis elegans. J. Hazard. Mater. 2012, 219–220, 221–230. [Google Scholar] [CrossRef] [PubMed]
- Mohri-Shiomi, A.; Garsin, D.A. Insulin signaling and the heat shock response modulate protein homeostasis in the Caenorhabditis elegans intestine during infection. J. Biol. Chem. 2008, 283, 194–201. [Google Scholar] [CrossRef] [PubMed]
- Frydman, J. Folding of newly translated proteins in vivo: The role of molecular chaperones. Annu. Rev. Biochem. 2001, 70, 603–647. [Google Scholar] [CrossRef] [PubMed]
- Lithgow, G.J. Invertebrate gerontology: The age mutations of Caenorhabditis elegans. Bioessays 1996, 18, 809–815. [Google Scholar] [CrossRef] [PubMed]
- Murphy, C.T.; Hu, P.J. Insulin/Insulin-like Growth Factor Signaling in C. elegans. Wormbook 2013. [Google Scholar] [CrossRef] [PubMed]
- Hsu, A.L.; Murphy, C.T.; Kenyon, C. Regulation of aging and age-related disease by DAF-16 and heat-shock factor. Science 2003, 300, 1142–1145. [Google Scholar] [CrossRef] [PubMed]
- Roh, J.Y.; Sim, S.J.; Yi, J.; Park, K.; Chung, K.H.; Ryu, D.Y.; Choi, J. Ecotoxicity of silver nanoparticles on the soil nematode Caenorhabditis elegans using functional ecotoxicogenomics. Environ. Sci. Technol. 2009, 43, 3933–3940. [Google Scholar] [CrossRef] [PubMed]
- Accili, D.; Arden, K.C. FoxOs at the crossroads of cellular metabolism, differentiation, and transformation. Cell 2004, 117, 421–426. [Google Scholar] [CrossRef]
- Henderson, S.T.; Johnson, T.E. daf-16 integrates developmental and environmental inputs to mediate aging in the nematode Caenorhabditis elegans. Curr. Biol. 2001, 11, 1975–1980. [Google Scholar] [CrossRef]
- Vanfleteren, J.R. Oxidative stress and ageing in Caenorhabditis elegans. Biochem. J. 1993, 292, 605–608. [Google Scholar] [CrossRef] [PubMed]
- Kobayashi, Y.; Furukawa-Hibi, Y.; Chen, C.; Horio, Y.; Isobe, K.; Ikeda, K.; Motoyama, N. SIRT1 is critical regulator of FOXO-mediated transcription in response to oxidative stress. Int. J. Mol. Med. 2005, 16, 237–244. [Google Scholar] [CrossRef] [PubMed]
- Stiernagle, T. Maintenance of C. elegans. Wormbook 2006. [Google Scholar] [CrossRef] [PubMed]
- Lewis, J.A.; Fleming, J.T. Basic culture methods. Methods Cell Biol. 1995, 48, 3–29. [Google Scholar] [PubMed]
- Oh, S.W.; Mukhopadhyay, A.; Svrzikapa, N.; Jiang, F.; Davis, R.J.; Tissenbaum, H.A. JNK regulates lifespan in Caenorhabditis elegans by modulating nuclear translocation of forkhead transcription factor/DAF-16. Proc. Natl. Acad. Sci. USA 2005, 102, 4494–4499. [Google Scholar] [CrossRef] [PubMed]
Sample Availability: Sample of the compound 1,4-naphthoquinone is available from the authors. |

© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, J.; Zeng, G.; Huang, X.; Wang, Z.; Tan, N. 1,4-Naphthoquinone Triggers Nematode Lethality by Inducing Oxidative Stress and Activating Insulin/IGF Signaling Pathway in Caenorhabditis elegans. Molecules 2017, 22, 798. https://doi.org/10.3390/molecules22050798
Wang J, Zeng G, Huang X, Wang Z, Tan N. 1,4-Naphthoquinone Triggers Nematode Lethality by Inducing Oxidative Stress and Activating Insulin/IGF Signaling Pathway in Caenorhabditis elegans. Molecules. 2017; 22(5):798. https://doi.org/10.3390/molecules22050798
Chicago/Turabian StyleWang, Jia, Guangzhi Zeng, Xiaobing Huang, Zhe Wang, and Ninghua Tan. 2017. "1,4-Naphthoquinone Triggers Nematode Lethality by Inducing Oxidative Stress and Activating Insulin/IGF Signaling Pathway in Caenorhabditis elegans" Molecules 22, no. 5: 798. https://doi.org/10.3390/molecules22050798