A Guide to the Variability of Flavonoids in Brassica oleracea


Abstract
:
1. Introduction
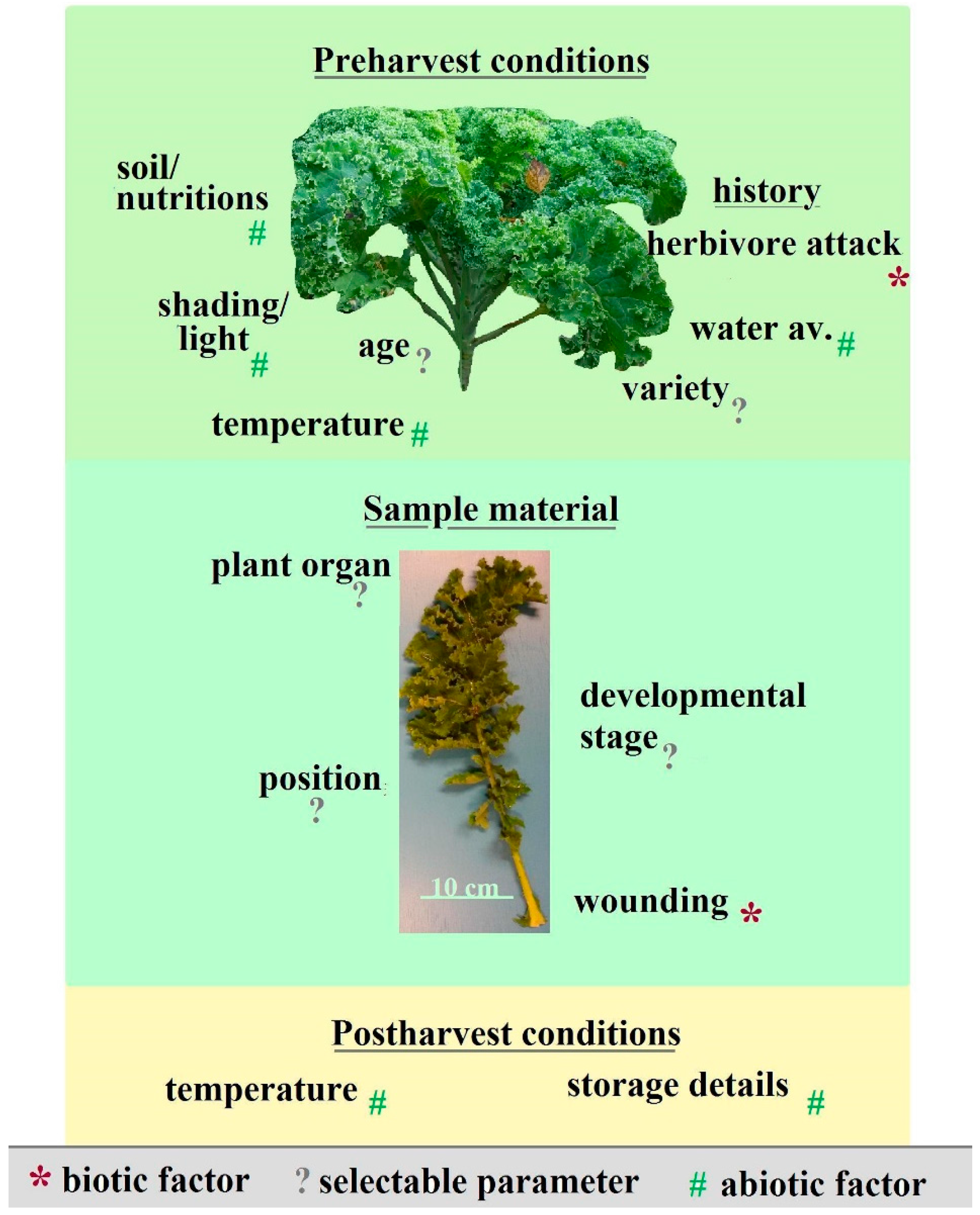
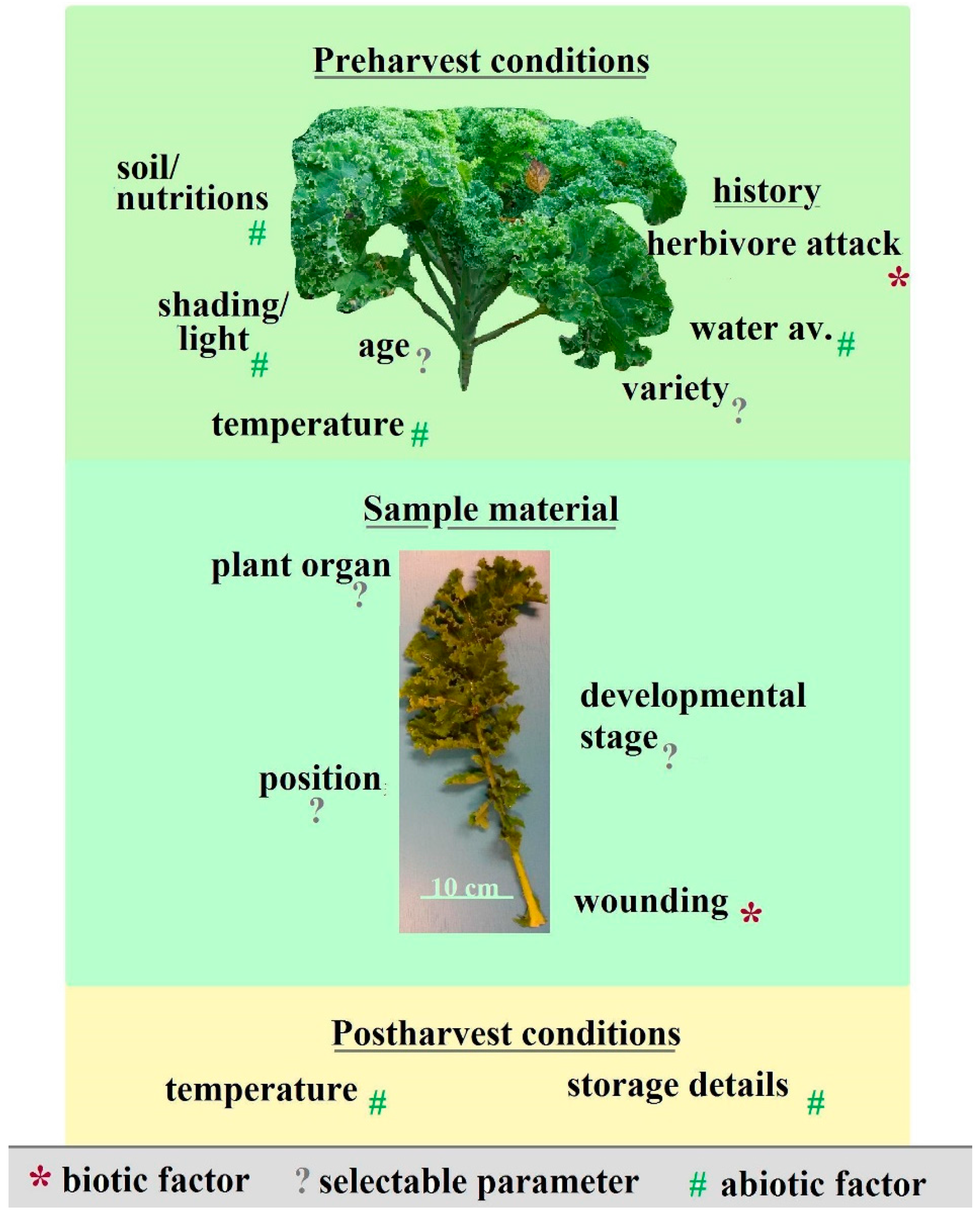
2. Data Comparability: Telling the Whole Story
3. Dealing with Complexity
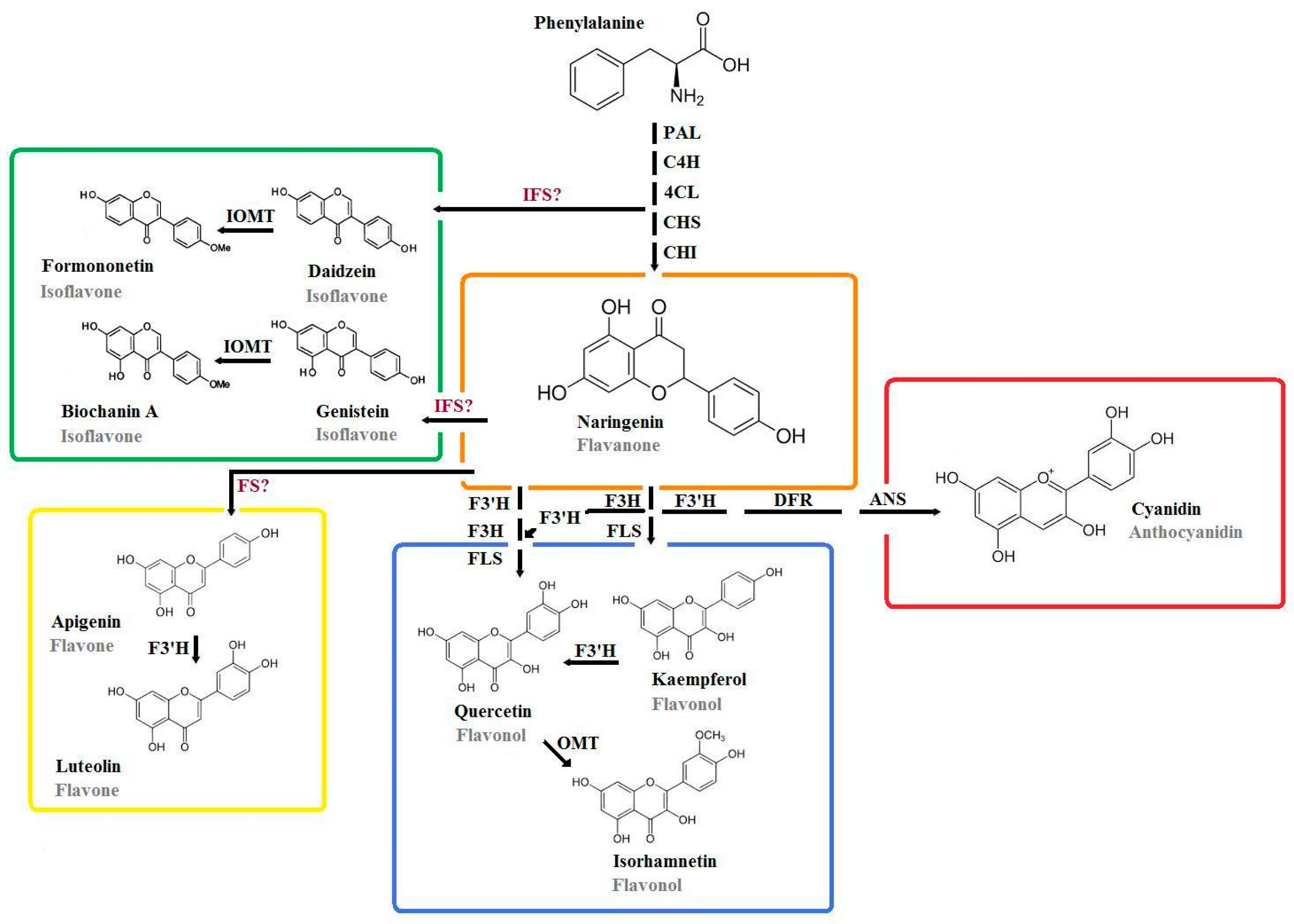
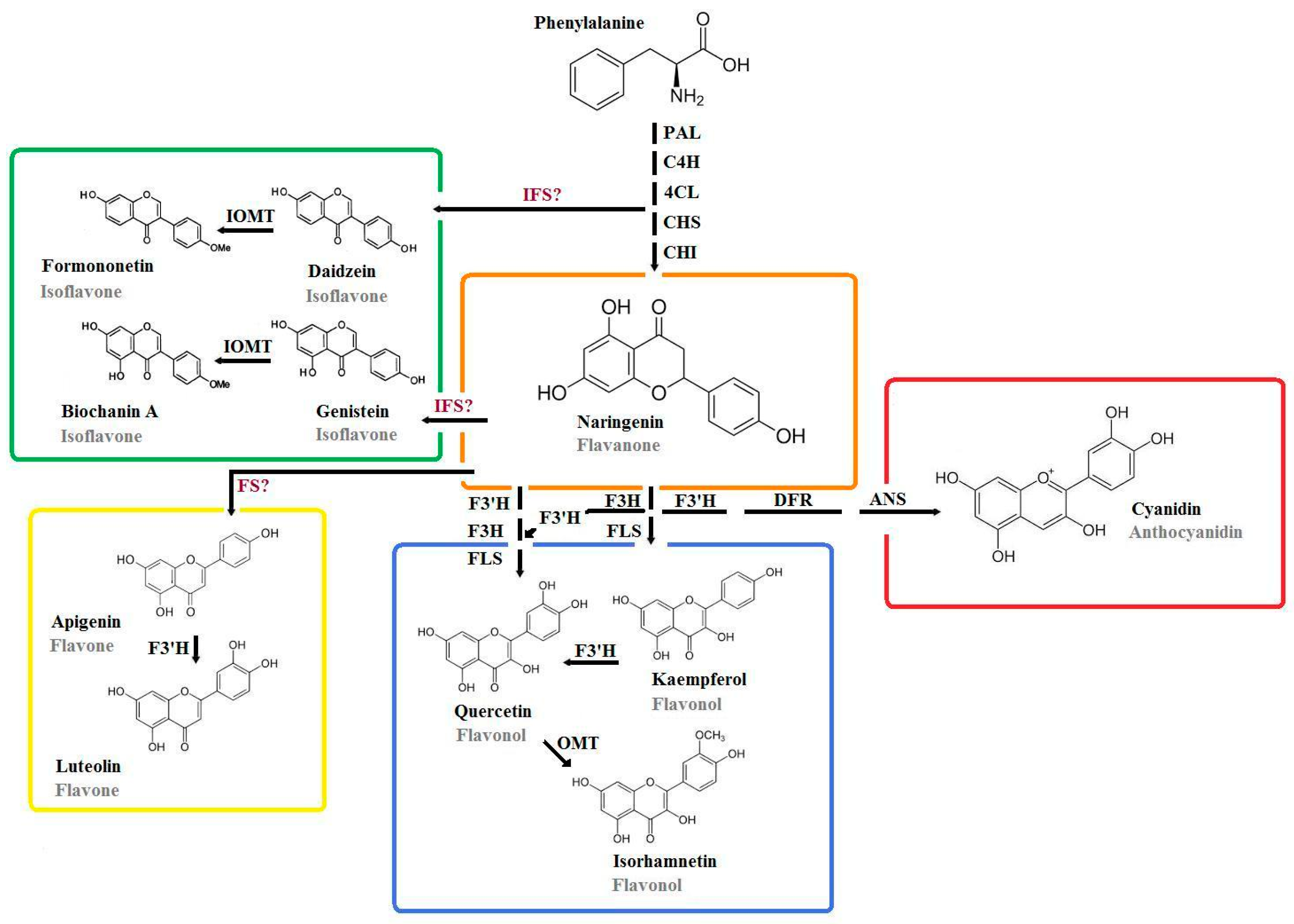
4. Specific Flavonoids of Major Interest
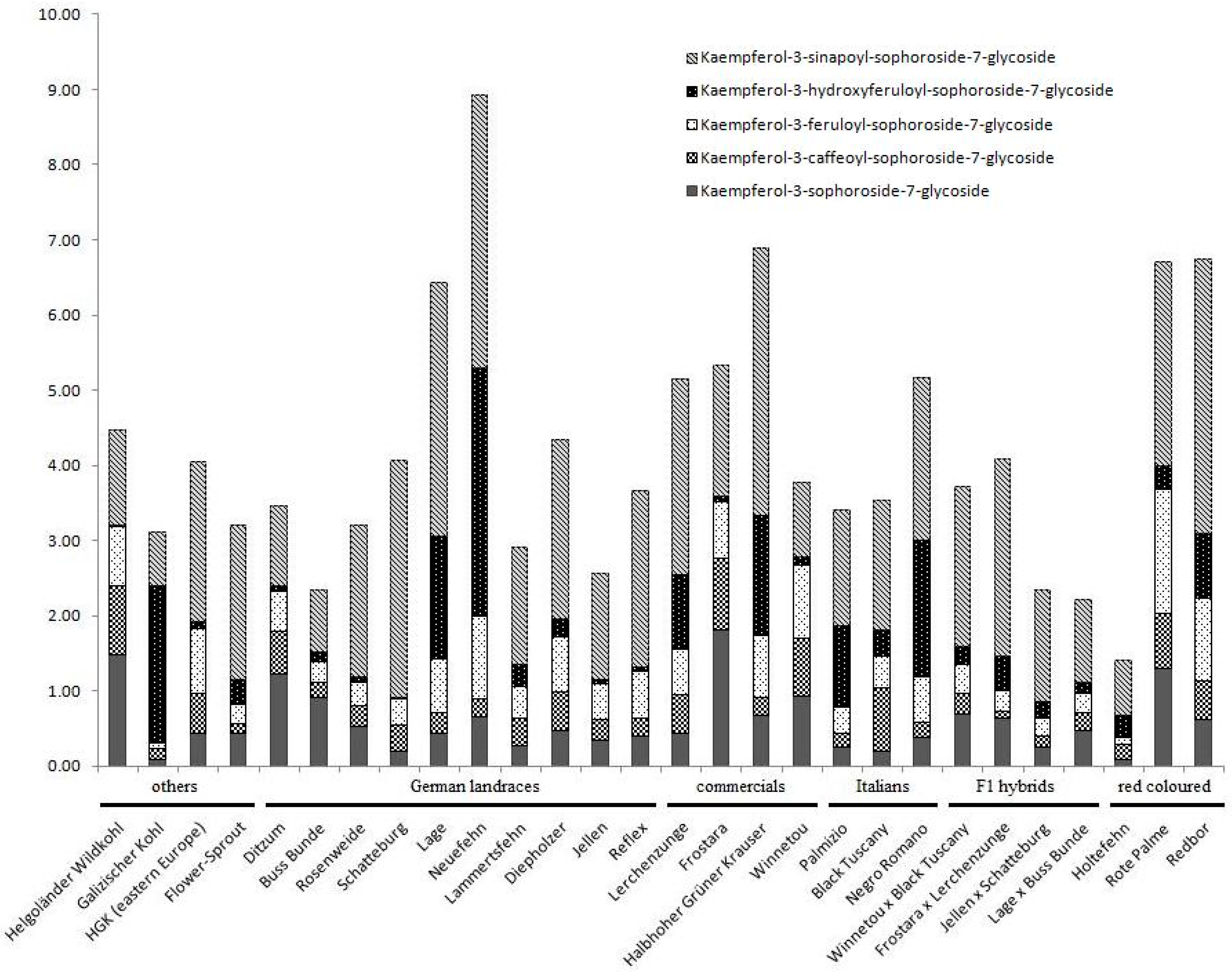
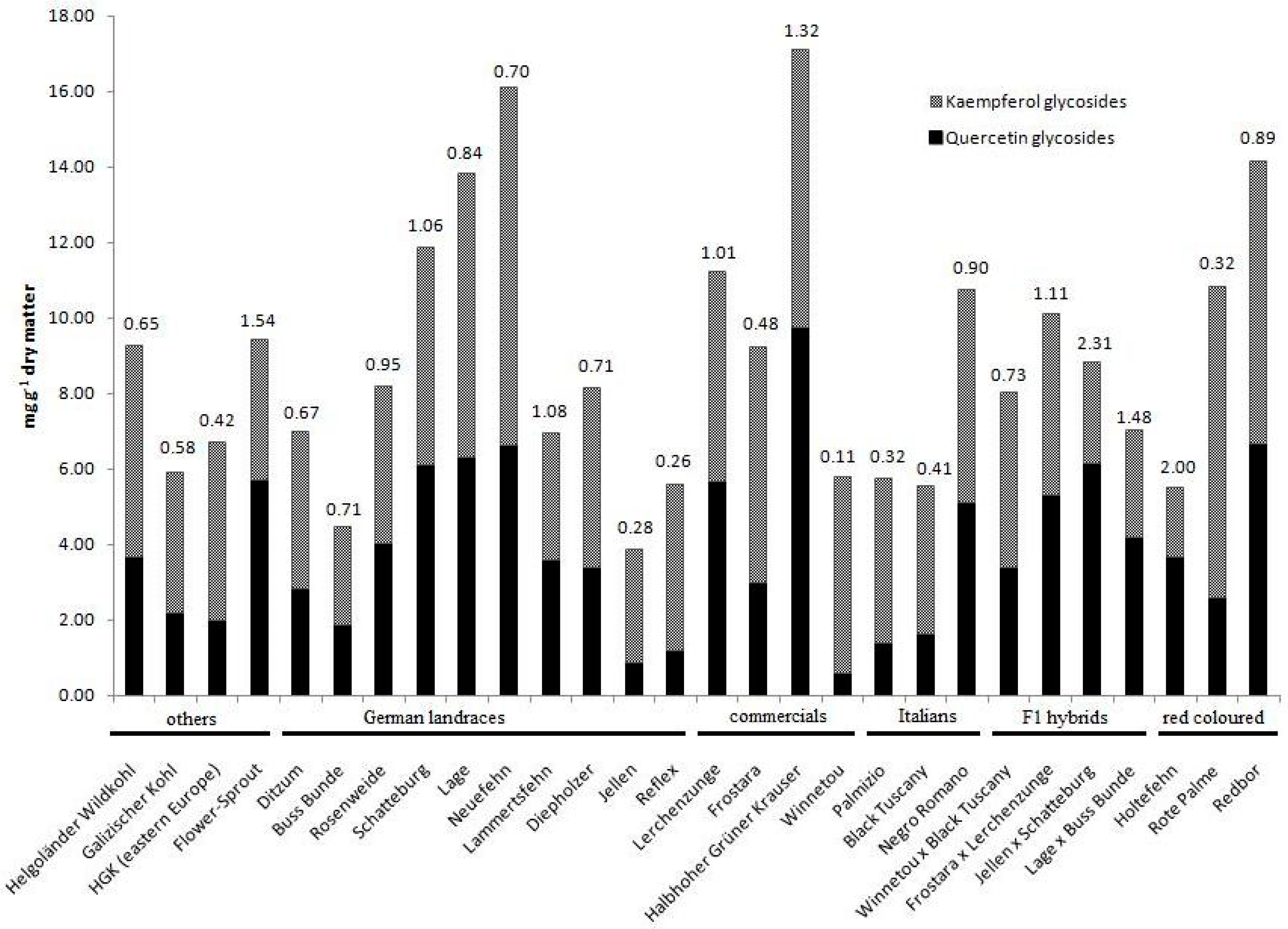
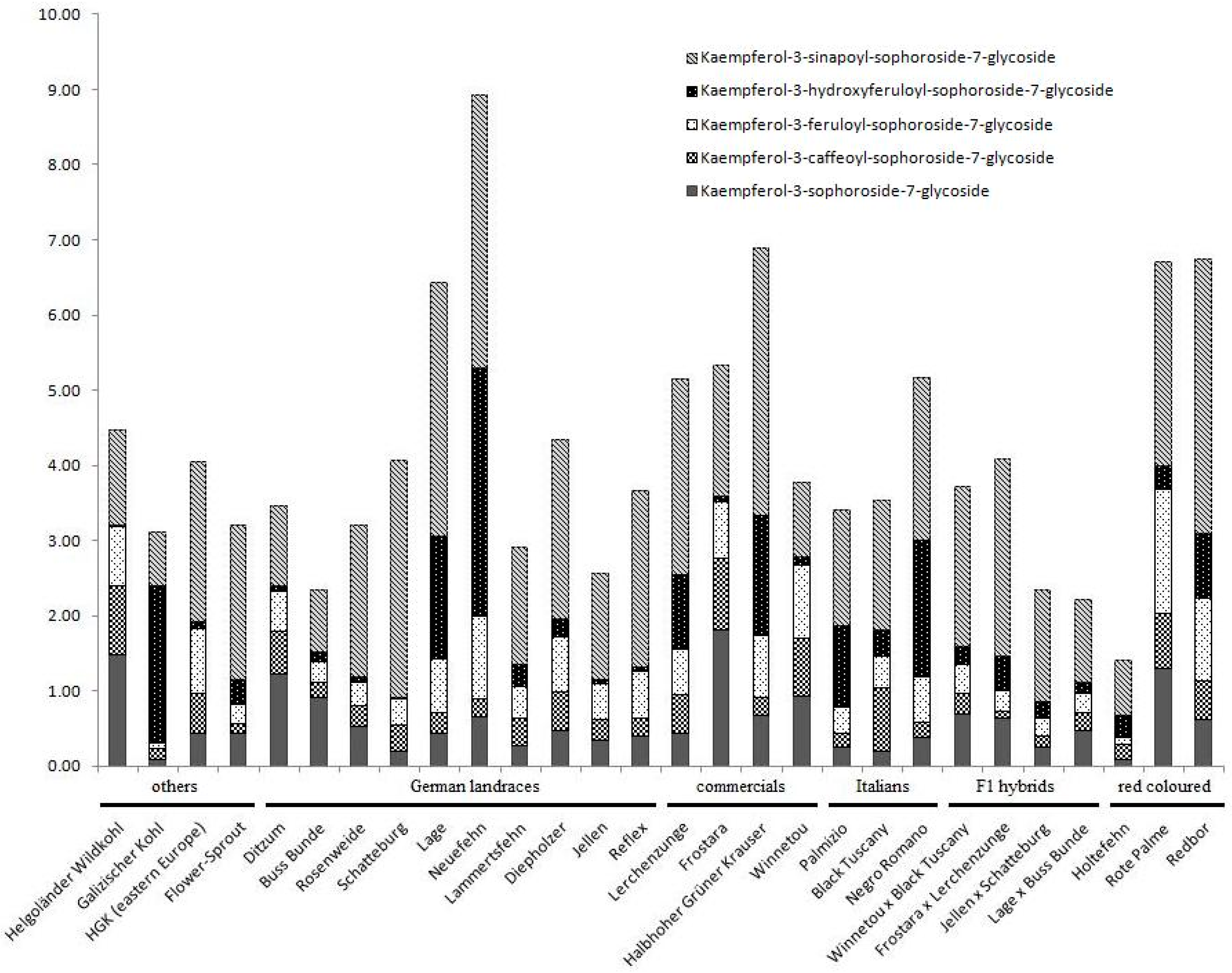
5. Compound Ratios as Plant Character
6. Future Perspectives
Supplementary Materials
Acknowledgments
Author Contributions
Conflicts of Interest
Appendix A
A1. Plant Material
A2. Flavonol and Anthocyanin Quantification
References
- FAOSTAT. Available online: www.faostat3.fao.org/home/E (accessed on 5 August 2015).
- Kushad, M.M.; Brown, A.F.; Kurilich, A.C.; Juvik, J.A.; Klein, B.P.; Wallig, M.A.; Jeffery, E.H. Variation of Glucosinolates in Vegetable Crops of Brassica oleracea. J. Agric. Food Chem. 1999, 47, 1541–1548. [Google Scholar] [CrossRef] [PubMed]
- Koh, E.; Wimalasiri, K.; Chassy, A.; Mitchell, A. Content of ascorbic acid, quercetin, kaempferol and total phenolics in commercial broccoli. J. Food Compos. Anal. 2009, 22, 637–643. [Google Scholar] [CrossRef]
- Agati, G.; Azzarello, E.; Pollastri, S.; Tattini, M. Flavonoids as antioxidants in plants: Location and functional significance. Plant Sci. 2012, 196, 67–76. [Google Scholar] [CrossRef] [PubMed]
- Yao, L.H.; Jiang, Y.M.; Shi, J.; Tomás-Barberán, F.A.; Datta, N.; Singanusong, R.; Chen, S.S. Flavonoids in foods and their health benefits. Plant. Food Hum. Nutr. 2004, 59, 113–122. [Google Scholar] [CrossRef]
- Kris-Etherton, P.; Lefevre, M.; Beecher, G.; Gross, M.; Keen, C.; Etherton, T. Bioactive compounds in nutrition and health-research methodologies for establishing biological function: The antioxidant and anti-inflammatory effects of flavonoids on atherosclerosis. Annu. Rev. Nutr. 2004, 24, 511–538. [Google Scholar] [CrossRef] [PubMed]
- Halliwell, B.; Rafter, J.; Jenner, A. Health promotion by flavonoids, tocopherols, tocotrienols, and other phenols: Direct or indirect effects? Antioxidant or not? Am. J. Clin. Nutr. 2005, 81, 268–276. [Google Scholar]
- Cartea, M.E.; Francisco, M.; Soengas, P.; Velasco, P. Phenolic Compounds in Brassica Vegetables. Molecules 2010, 16, 251–280. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Erdman, J.W., Jr.; Balentine, D.; Arab, L.; Arab, L.; Beecher, G.; Dwyer, J.T.; Folts, J.; Harnly, J.; Hollman, P.; Keen, C.L.; et al. Flavonoids and Heart Health: Proceedings of the ILSI North America Flavonoids Workshop, 31 May–1 June 2005, Washington, DC. J. Nutr. 2007, 137, 718–737. [Google Scholar]
- Zietz, M.; Annika, W.; Schmidt, S.; Rohn, S.; Schreiner, M.; Krumbein, A.; Kroh, L.W. Genotypic and Climatic Influence on the Antioxidant Activity of Flavonoids in Kale (Brassica oleracea var. sabellica). J. Agric. Food Chem. 2010, 58, 2123–2130. [Google Scholar] [CrossRef] [PubMed]
- Schmidt, S.; Zietz, M.; Schreiner, M.; Rohn, S.; Kroh, L.W.; Krumbein, A. Genotypic and climatic influences on the concentration and composition of flavonoids in kale (Brassica oleracea var. sabellica). Food Chem. 2010, 119, 1293–1299. [Google Scholar] [CrossRef]
- Reilly, K.; Valverde, J.; Finn, L.; Rai, D.K.; Brunton, N.; Sorensen, J.C.; Sorensen, H.; Gaffney, M. Potential of cultivar and crop management to affect phytochemical content in winter-grown sprouting broccoli (Brassica oleracea L var. italica). J. Sci. Food Agric. 2013, 94, 322–330. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sousa, C.; Lopes, G.; Pereira, D.; Taveira, M.; Valentao, P.; Seabra, R.; Pereira, J.; Baptista, P.; Ferreres, F.; Andrade, P. Screening of Antioxidant Compounds During Sprouting of Brassica oleracea L. var. costata DC. Comb. Chem. High Throughput Screen. 2007, 10, 377–386. [Google Scholar] [CrossRef] [PubMed]
- Šamec, D.; Piljac-Žegarac, J.; Bogović, M.; Habjanič, K.; Grúz, J. Antioxidant potency of white (Brassica oleracea L var. capitata) and Chinese (Brassica rapa L var. pekensis (Lour)) cabbage: The influence of development stage, cultivar choice and seed selection. Sci. Hort. 2011, 128, 78–83. [Google Scholar]
- Soengas, P.; Cartea, M.; Francisco, M.; Sotelo, T.; Velasco, P. New insights into antioxidant activity of Brassica crops. Food Chem. 2012, 134, 725–733. [Google Scholar] [CrossRef] [PubMed]
- Sousa, C.; Pereira, D.M.; Pereira, J.A.; Bento, A.; Rodrigues, M.A.; Dopico-Garcia, A.; Valentão, P.; Lopes, G.; Ferreres, F.; Seabra, R.M.; et al. Multivariate Analysis of Tronchuda Cabbage (Brassica oleracea L var. costata DC) Phenolics: Influence of Fertilizers. J. Agric. Food Chem. 2008, 56, 2231–2239. [Google Scholar] [PubMed]
- Sousa, C.; Valentão, P.; Ferreres, F.; Seabra, R.M.; Andrade, P.B. Tronchuda Cabbage (Brassica oleracea L var var costata DC): Scavenger of Reactive Nitrogen Species. J. Agric. Food Chem. 2008, 56, 4205–4211. [Google Scholar] [CrossRef] [PubMed]
- Treutter, D. Significance of flavonoids in plant resistance: A review. Environ. Chem. Lett. 2006, 4, 147–157. [Google Scholar] [CrossRef]
- Ferrieri, A.P.; Appel, H.M.; Schultz, J.C. Plant Vascular Architecture Determines the Pattern of Herbivore-Induced Systemic Responses in Arabidopsis thaliana. PLoS ONE 2015, 10, e0123899. [Google Scholar] [CrossRef] [PubMed]
- Fallovo, C.; Schreiner, M.; Schwarz, D.; Colla, G.; Krumbein, A. Phytochemical changes induced by different nitrogen supply forms and radiation levels in two leafy Brassica species. J. Agric. Food Chem. 2011, 59, 4198–4207. [Google Scholar] [CrossRef] [PubMed]
- Neugart, S.; Fiol, M.; Schreiner, M.; Rohn, S.; Zrenner, R.; Kroh, L.W.; Krumbein, A. Low and moderate photosynthetically active radiation affects the flavonol glycosides and hydroxycinnamic acid derivatives in kale (Brassica oleracea var. sabellica) dependent on two low temperatures. Plant Physiol. Biochem. 2013, 72, 161–168. [Google Scholar] [PubMed]
- Neugart, S.; Zietz, M.; Schreiner, M.; Rohn, S.; Kroh, L.W.; Krumbein, A. Structurally different flavonol glycosides and hydroxycinnamic acid derivatives respond differently to moderate UV-B radiation exposure. Physiol. Plant 2012, 145, 582–593. [Google Scholar] [CrossRef] [PubMed]
- Naguib, A.E.-M.M.; El-Baz, F.K.; Salama, Z.A.; Hanaa, H.A.E.B.; Ali, H.F.; Gaafar, A.A. Enhancement of phenolics, flavonoids and glucosinolates of Broccoli (Brassica olaracea var. italica) as antioxidants in response to organic and bio-organic fertilizers. J. Saudi Soc. Agric. Sci. 2012, 11, 135–142. [Google Scholar]
- Valverde, J.; Reilly, K.; Villacreces, S.; Gaffney, M.; Grant, J.; Brunton, N. Variation in bioactive content in broccoli (Brassica oleracea var. italica) grown under conventional and organic production systems. J. Sci. Food Agric. 2014, 95, 1163–1171. [Google Scholar] [PubMed]
- Groenbaek, M.; Jensen, S.; Neugart, S.; Schreiner, M.; Kidmose, U.; Kristensen, H.L. Influence of Cultivar and Fertilizer Approach on Curly Kale (Brassica oleracea L. var. sabellica). 1. Genetic Diversity Reflected in Agronomic Characteristics and Phytochemical Concentration. J. Agric. Food Chem. 2014, 62, 11393–11402. [Google Scholar] [PubMed]
- Pękal, A.; Pyrzynska, K. Evaluation of Aluminium Complexation Reaction for Flavonoid Content Assay. Food Anal. Methods 2014, 7, 1776–1782. [Google Scholar] [CrossRef]
- Hagen, S.F.; Borge, G.I.A.; Solhaug, K.A.; Bengtsson, G.B. Effect of cold storage and harvest date on bioactive compounds in curly kale (Brassica oleracea L var. acephala). Postharvest Biol. Technol. 2009, 51, 36–42. [Google Scholar] [CrossRef]
- Vallejo, F.; Tomás-Barberán, F.; García-Viguera, C. Health-Promoting Compounds in broccoli as Influenced by Refrigerated Transport and Retail Sale Period. J. Agric. Food Chem. 2003, 51, 3029–3034. [Google Scholar] [CrossRef] [PubMed]
- Bahorun, T.; Luximon-Ramma, A.; Crozier, A.; Aruoma, O.I. Total phenol, flavonoid, proanthocyanidin and vitamin C levels and antioxidant activities of Mauritian vegetables. J. Sci. Food Agric. 2004, 84, 1553–1561. [Google Scholar] [CrossRef]
- Heimler, D.; Vignolini, P.; Dini, M.G.; Vincieri, F.F.; Romani, A. Antiradical activity and polyphenol composition of local Brassicaceae edible varieties. Food Chem. 2006, 99, 464–469. [Google Scholar] [CrossRef]
- Jacob, J.A.; Mahal, H.; Mukherjee, T.; Kapoor, S. Free radical reactions with the extract of brassica family. Food Chem. 2011, 129, 1132–1138. [Google Scholar] [CrossRef] [PubMed]
- Jaiswal, A.K.; Abu-Ghannam, N.; Gupta, S. A comparative study on the polyphenolic content, antibacterial activity and antioxidant capacity of different solvent extracts of Brassica oleracea vegetables. Int. J. Food Sci. Technol. 2011, 47, 223–231. [Google Scholar] [CrossRef]
- Lola-Luz, T.; Hennequart, F.; Gaffney, M. Effect on yield, total phenolic, total flavonoid and total isothiocyanate content of two broccoli cultivars (Brassica oleracea var. italica) following the application of a commercial brown seaweed extract (Ascophyllum. nodosum). Agric. Food Sci. 2014, 23, 28–37. [Google Scholar]
- Strack, D. Phenolic compounds. In Plant Biochemistry, 1st ed.; Dey, P., Harborne, J., Eds.; Academic Press: San Diego, CA, USA, 1997; pp. 387–416. [Google Scholar]
- Pollastri, S.; Tattini, M. Flavonols: Old compounds for old roles. Ann. Bot. 2011, 108, 1225–1233. [Google Scholar] [CrossRef] [PubMed]
- Hollman, P.C.; Arts, I.C. Flavonols, flavones and flavanols—Nature, occurrence and dietary burden. J. Sci. Food Agric. 2000, 80, 1081–1093. [Google Scholar] [CrossRef]
- Beecher, G.R. Overview of dietary flavonoids: Nomenclature, occurrence and intake. J. Nutr. 2003, 133, 3248S–3254S. [Google Scholar] [PubMed]
- Heim, K.E.; Tagliaferro, A.R.; Bobilya, D.J. Flavonoid antioxidants: Chemistry, metabolism and structure-activity relationships. J. Nutr. Biochem. 2002, 13, 572–584. [Google Scholar] [CrossRef]
- Podsedek, A. Natural antioxidants and antioxidant capacity of Brassica vegetables: A review. LWT—Food Sci. Technol. 2007, 40, 1–11. [Google Scholar] [CrossRef]
- Winkel-Shirley, B. Biosynthesis of flavonoids and effects of stress. Curr. Opin. Plant Biol. 2002, 5, 218–223. [Google Scholar] [CrossRef]
- Ponce, MA.; Scervino, J.M.; Erra-Balsells, R.; Ocampo, J.A.; Godeas, A.M. Flavonoids from shoots, roots and roots exudates of Brassica alba. Phytochemistry 2004, 65, 3131–3134. [Google Scholar] [CrossRef] [PubMed]
- Martens, S.; Mithöfer, A. Flavones and flavone synthases. Phytochemistry 2005, 66, 2399–2407. [Google Scholar] [CrossRef] [PubMed]
- Sakakibara, H.; Honda, Y.; Nakagawa, S.; Ashida, H.; Kanazawa, K. Simultaneous Determination of All Polyphenols in Vegetables, Fruits, and Teas. J. Agric. Food Chem. 2003, 51, 571–581. [Google Scholar] [CrossRef] [PubMed]
- Miean, K.H.; Mohamed, S. Flavonoid (Myricetin, Quercetin, Kaempferol, Luteolin, and Apigenin) Content of Edible Tropical Plants. J. Agric. Food Chem. 2001, 49, 3106–3112. [Google Scholar] [CrossRef] [PubMed]
- Cao, J.; Chen, W.; Zhang, Y.; Zhang, Y.; Zhao, X. Content of Selected Flavonoids in 100 Edible Vegetables and Fruits. Food Sci. Technol. Res. 2010, 16, 395–402. [Google Scholar] [CrossRef]
- Wu, X.; Prior, R.L. Identification and Characterization of Anthocyanins by High-Performance Liquid Chromatography-Electrospray Ionization-Tandem Mass Spectrometry in Common Foods in the United States: Vegetables, Nuts, and Grains. J. Agric. Food Chem. 2005, 53, 3101–3113. [Google Scholar] [CrossRef] [PubMed]
- Scalzo, R.L.; Genna, A.; Branca, F.; Chedin, M.; Chassaigne, H. Anthocyanin composition of cauliflower (Brassica oleracea L var. botrytis) and cabbage (B. oleracea L var. capitata) and its stability in relation to thermal treatments. Food Chem. 2008, 107, 136–144. [Google Scholar]
- Philips, D.A.; Kapulnik, Y. Plant isoflavonoids, pathogens and symbionts. Trends Microbiol. 1995, 58, 58–64. [Google Scholar] [CrossRef]
- Kuhnle, G.G.; Dell’Aquila, C.; Aspinall, S.M.; Runswick, S.A.; Joosen, A.M.; Mulligan, A.A.; Bingham, S.A. Phytoestrogen content of fruits and vegetables commonly consumed in the UK based on LC–MS and 13C-labelled standards. Food Chem. 2009, 116, 542–554. [Google Scholar] [CrossRef]
- Olsen, H.; Aaby, K.; Borge, G.I.A. Characterization and Quantification of Flavonoids and Hydroxycinnamic Acids in Curly Kale (Brassica oleracea L convar. acephala var. sabellica) by HPLC-DAD-ESI-MSn. J. Agric. Food Chem. 2009, 57, 2816–2825. [Google Scholar] [PubMed]
- Rice-Evans, C.A.; Miller, N.J.; Paganga, G. Structure-antioxidant activity relationships of flavonoids and phenolic acids. Free Radic. Biol. Med. 1996, 20, 933–956. [Google Scholar] [CrossRef]
- Tabart, J.; Kevers, C.; Pincemail, J.; Defraigne, J.-O.; Dommes, J. Comparative antioxidant capacities of phenolic compounds measured by various tests. Food Chem. 2009, 113, 1226–1233. [Google Scholar] [CrossRef]
- Vallejo, F.; Tomás-Barberán, F.; Ferreres, F. Characterisation of flavonols in broccoli (Brassica oleracea L var. italica) by liquid chromatography–UV diode-array detection-electrospray ionisation mass spectrometry. J. Chromatogr. A 2004, 1054, 181–193. [Google Scholar] [PubMed]
- Lin, L.-Z.; Harnly, J.; Zhang, R.-W.; Fan, X.-E.; Chen, H.-J. Quantitation of the Hydroxycinnamic Acid Derivatives and the Glycosides of Flavonols and Flavones by UV Absorbance after Identification by LC-MS. J. Agric. Food Chem. 2012, 60, 544–553. [Google Scholar] [CrossRef] [PubMed]
- Neugart, S.; Fiol, M.; Schreiner, M.; Rohn, S.; Zrenner, R.; Kroh, L.W.; Krumbein, A. Interaction of Moderate UV-B Exposure and Temperature on the Formation of Structurally Different Flavonol Glycosides and Hydroxycinnamic Acid Derivatives in Kale (Brassica oleracea var. sabellica). J. Agric. Food Chem. 2014, 62, 4054–4062. [Google Scholar] [CrossRef] [PubMed]
- Neugart, S.; Krumbein, A.; Zrenner, R. Influence of Light and Temperature on Gene Expression Leading to Accumulation of Specific Flavonol Glycosides and Hydroxycinnamic Acid Derivatives in Kale (Brassica oleracea var. sabellica). Front. Plant Sci. 2016, 7, 326. [Google Scholar] [CrossRef] [PubMed]
- Pliszka, B.; Huszcza-Ciołkowska, G.; Mieleszko, E.; Czaplicki, S. Stability and antioxidative properties of acylated anthocyanins in three cultivars of red cabbage (Brassica oleracea L. var. capitata L. f. rubra). J. Sci. Food Agric. 2009, 89, 1154–1158. [Google Scholar] [CrossRef]
- Julkunen-Tiitto, R.; Nenadis, N.; Neugart, S.; Robson, M.; Agati, G.; Vepsäläinen, J.; Zipoli, G.; Nybakken, L.; Winkler, B.; Jansen, M.A.K. Assessing the response of plant flavonoids to UV radiation: An overview of appropriate techniques. Phytochem. Rev. 2014, 14, 273–297. [Google Scholar] [CrossRef]
- Moreno, D.A.; Pérez-Balibrea, S.; Ferreres, F.; Gil-Izquierdo, Á.; García-Viguera, C. Acylated anthocyanins in broccoli sprouts. Food Chem. 2010, 123, 358–363. [Google Scholar] [CrossRef]
- Sikora, E.; Cieślik, E.; Filipiak-Florkiewicz, A.; Leszczyńska, T. Effect of hydrothermal processing on phenolic acids and flavonols contents in selected Brassica vegetables. Acta Sci. Pol. Technol. Aliment. 2012, 11, 45–51. [Google Scholar] [PubMed]
- Mazur, W.; Adlercreutz, H. Natural and anthropogenic environmental oestrogens: The scientific basis for risk assessment. Pure Appl. Chem. 1998, 70, 1759–1776. [Google Scholar] [CrossRef]
- Lapcik, O.; Honys, D.; Koblovska, R.; Mackova, Z.; Vitkova, M.; Klejdus, B. Isoflavonoids are present in Arabidopsis thaliana despite the absence of any homologue to known isoflavonoid synthases. Plant Physiol. Biochem. 2006, 44, 106–114. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Reference | Brassica oleracea Variety | Preharvest Conditions | Sample Material | Post-Harvest Conditions | Method | Total Flavonoid Content | Flavonoids Quantified | |
|---|---|---|---|---|---|---|---|---|
| Variety and Cultivar | Age; Nutrition, Light, Temperature | Plant Organ | Position & Age | Temperature & Storage Details | Considered as | |||
| Bahorun et al. [29] | italica cv “Packman” italica cv “Kashmere” capitata cv “KKCross” | ? | Flower Flower Leaves | ? | Given in detail | HPLC | Quercetin equivalents | Myricetin, quercetin, kaempferol, apigenin and luteolin aglycones after acid hydrolysis |
| Heimler et al. [30] | capitata cv ?; italica cv ?; acephala cv ?; sabauda cv ?; botrytis cv “Verde di Macerata”, “Snow ball”, gemnifera cv “Zencher” | ? | Edible part | ? | Given in detail | HPLC | Catechin equivalents | Kaempferol-3-[2-sinapoylglucopyranosyl(1,2) glucopyranoside]-7-[glucopyranosyl(1,4) glucopyranoside], kaempferol-3-[-2-feruloylglucopyranosyl(1,2) glucopyranoside]-7-[glucopyranosyl(1,4) glucopyranoside], kaempferol tetraglycoside, kaempferol sinapoyl tetraglycoside, kaempferol cumaroyl tetraglycoside, kaempferol diglycoside; Quercetin-glucoside |
| Jacob et al. [31] | capitata cv?; capitata var. rubra cv? | ? | Edible parts | ? | ? | Spectrophotometry, AlCl3 | Epicatechin equivalents | Method is specific for rutin, luteolin and catechins (Pękal and Pyrzynska [26]) |
| Jaiswal et al. [32] | capitata f. alba cv?; italica cv?; gemmifera cv? | ? | Edible parts | Positions given, ages? | ? | HPLC | Quercetin equivalents | ? |
| Lola-Luz et al. [33] | italica cv “Ironman” and “Red Admiral” | Three-week range, Timepoints vary; Given in detail | Heads/florets | ? | −20 °C, two-week range | Spectrophotometry, AlCl3 | Catechin equivalents | Method is specific for rutin, luteolin and catechins (Pękal and Pyrzynska [26]) |
| Naguib et al. [23] | italica cv “Calabrese”, “Southern star” | ?; Given in detail; field cond.; location given | Florets | ? | Given in detail | Spectrophotometry, AlCl3 | Quercetin equivalents | Method is specific for rutin, luteolin and catechins (Pękal and Pyrzynska [26]) |
| Reilly et al. [12] | italica cv “TZ6002”, “TZ5055”, “TZ5052”, “TZ4043”, “Red Admiral”, “Ironman” | Given in detail | Leaf, immature Primary floret, mature primary floret, secondary floret, flower | ? | Given in detail | Spectrophotometry, AlCl3 | Catechin equivalents | Method is specific for rutin, luteolin and catechins (Pękal and Pyrzynska [26]) |
| Valverde et al. [24] | italica cv “Belstar”, “Fiesta” | Given in detail | Primary florets | ? | −20 °C, 24-h range | Spectrophotometry, AlCl3 | Catechin equivalents | Method is specific for rutin, luteolin and catechins (Pękal and Pyrzynska [26]) |
| Flavonoid (Subclass) | Physiological Function | Relevance in Medical Research | Content in B. oleracea [Method; Variety] | Intraspecific Variation Factor | References |
|---|---|---|---|---|---|
| Apigenin and luteolin (Flavone) | Nodulation and general defence mechanisms 1 Resistance to mycorrhization 2 | Phytoestrogen with antibacterial and anti-inflammatory functions; apoptosis-inducer 3 | Occurrence contradictory discussed 4–6; ~3-30 mg/kg fw [HPLC; alba, botrytis, capitata] 7 | Apigenin ~2 Luteolin ~ 2.5 7 | 1 Winkel-Shirley [40] 2 Ponce et al. [41] 3 Martens and Mithöfer [42] 4−6 Bahorum et al. [29]; Sakakibara et al. [43]; Miean and Mohamed [44] 7 Cao et al. [45] |
| Cyanidin (Anthocyanidin) | Pigmentation of flowers and fruits for recruitment of pollinators and seed dispersers 1 | Antioxidant, anti-inflammatory, antimicrobial & anticarcinogenic activities, positive effect on visual performance & neuroprotection 2 | 23 cyanidin derivative forms [HPLC; capitata f rubra] 3; ~40–750 mg/kg fw [HPLC; botrytis; capitata f rubra] 4 | 18; qualitative dominance shift in derivative forms 4 | 1 Winkel-Shirley [40] 2 Erdman et al. [9] 3 Wu and Prior [46] 4 Scalzo et al. [47] |
| Daidzein, genistein, glycitein, biochanin A and formononetin (Isoflavone) | Root bacteria interaction including symbionts and pathogenic microorganism 1 | Suggested to exert coronary benefits, directly reduce atherosclerosis and lower LDL-cholesterol 2 | Max ~10 µg/100g fw [LC/MS/MS; botrytis; capitata; capitata f alba; capitata f rubra; italica; gemmifera; sabellica; saubada] 3 | 13 (for all listed isoflavones together) 3 | 1 Philips and Kapulnik [48] 2 Erdman et al. [9] 3 Kuhnle et al. [49] |
| Kaempferol (Flavonol) | Prevent oxidative stress in chloroplasts 1; ROS reduction 1; photoprotection 1; free radical scavenging capacity 2 | Prevents coronary heart disease and chronic inflammation, suppresses cell proliferation in gut cancer lines, atherosclerosis prevention and growth inhibition of bacteria lines (gram-positive and gram-negative bacteria) 2 | ~60 mg/100 g fw [HPLC; sabellica] 4 | Qualitative dominance shift in glycosides2; ontogenetic dependent variation with subgroup specific patterns3 | 1 Pollastri and Tattini [35] 2 Cartea et al. [8] 3 Soengas et al. [15] 4 Olsen et al. [50] |
| Quercetin (Flavonol) | See kaempferol; chelate transition metal ions, auxin gradient regulation1 | See kaempferol | ~45 mg/100 g fw [HPLC; sabellica] 4 | Qualitative dominance shift in derivative forms 2; ontogenetic dependent variation with subgroup specific patterns 3 | 1 Pollastri and Tattini [35] 2 Cartea et al. [8] 3 Soengas et al. [15] 4 Olsen et al. [50] |
| A—Anthocyanins | |
| Anthocyanins (C—Cyanidin) | B. oleracea Variety Verification |
| C-3-(caffeoyl)-(p-coumaroyl)-diglycoside-5-glycoside | var. capitata f. rubra 3 |
| C-3-(caffeoyl)-diglycoside-5-glycoside | var. capitata f. rubra 3 |
| C-3-(caffeoyl)-diglycoside-5-glycoside | var. capitata f. rubra 3 |
| C-3-(feruloyl)-(feruloyl)-diglycoside-5-glycoside | var. capitata f. rubra 3 |
| C-3-(feruloyl)-diglycoside-5-glycoside | var. botrytis itálica 1 var. capitata f. rubra 3 |
| C-3-(glycopyranosyl)-(feruloyl)-diglycoside-5-glycoside | var. capitata f. rubra 3 |
| C-3-(glycopyranosyl)-(sinapoyl)-diglycoside-5-glycoside | var. capitata f. rubra 3 |
| C-3-(p-coumaroyl)-(sinapoyl)-diglycoside-5-(malonyl)-glycoside | var. botrytis italic 1 |
| C-3-(p-coumaroyl)-(sinapoyl)-diglycoside-5-glycoside | var. botrytis itálica 1 var. capitata f. rubra 3 |
| C-3-(p-coumaroyl)-diglycoside-5-glycoside | var. botrytis italic 1 |
| C-3-(p-hydroxybenzoyl)-(oxaloyl)-diglycoside-5-glycoside | var. capitata f. rubra 3 |
| C-3-(sinapoyl)-(feruloyl)-diglycoside-5-(malonyl)-glycoside | var. botrytis italic 1 |
| C-3-(sinapoyl)-(feruloyl)-diglycoside-5-glycoside | var. botrytis itálica 1 var. capitata f. rubra 3 var. sabellica 4 |
| C-3-(sinapoyl)-(sinapoyl)-diglycoside-5-(malonyl)-glycoside | var. botrytis italic 1 |
| C-3-(sinapoyl)-(sinapoyl)-diglycoside-5-glycoside | var. botrytis itálica 1 var. capitata f. rubra 3 var. sabellica 4 |
| C-3-(sinapoyl)-diglycoside-5-(sinapoyl)-glycoside | var. capitata f. rubra 3 |
| C-3-(sinapoyl)-diglycoside-5-glycoside | var. botrytis itálica 1 var. capitata f. rubra 3 |
| C-3-(sinapoyl)-diglycoside-5-xyloside | var. capitata f. rubra 3 |
| C-3-(sinapoyl)-glycoside-5-glycoside | var. capitata f. rubra 3 |
| C-3-(sinapoyl)-triglycoside-5-glycoside | var. botrytis italic 1 |
| C-3,5-diglycoside | var. botrytis itálica 1 var. capitata f. rubra 3 |
| C-3-diglycoside | var. capitata f. rubra 3 |
| C-3-diglycoside-5-glycoside | var. botrytis itálica 1 var. capitata f. rubra 3 |
| C-3-diglycoside-5-xyloside | var. capitata f. rubra 3 |
| C-3-(6-feruloyl)-sophoroside-5-(6-sinapyl)-glycoside | var. botrytis2 and var. capitata 2 |
| C-3-(6-feruloyl)-sophoroside-5-glycoside | var. botrytis2 and var. capitata 2 |
| C-3-(6-p-coumaryl)-sophoroside-5-(6-sinapyl)-glycoside | var. botrytis2 and var. capitata 2 |
| C-3-(6-p-coumaryl)-sophoroside-5-glycoside | var. botrytis2 and var. capitata 2 |
| C-3-(6-sinapyl)-sophoroside-5-(6-sinapyl)-glycoside | var. botrytis2 and var. capitata 2 |
| C-3-(6-sinapyl)-sophoroside-5-glycoside | var. botrytis2 and var. capitata 2 |
| C-3-glycoside-5-glycoside | var. capitata 2 |
| C-3-sophoroside-5-glycoside | var. botrytis 2 and var. capitata 2 |
| B—Flavonols | |
| Flavonol (Q—Quercetin; K—Kaempferol) | B. oleracea Variety Verification (as Reviewed by Cartea et al. [8]) |
| Q-3-O-sophorotrioside-7-O-sophoroside | var. acephala, var. botrytis italica |
| Q-3-O-sophorotrioside-7-glycoside | var. acephala, var. botrytis italica |
| Q-3-O-sophoroside-7-O-glycoside | var. capitata f. alba, var. acephala, var. botrytis, var. botrytis italica, var. costata |
| Q-3,7-di-O-glycoside | var. capitata f. alba, var. acephala, var. botrytis italica |
| Q-3-O-sophoroside | var. capitata f. alba, var. acephala, var. botrytis italica |
| Q-3-O-glycoside | var. botrytis italica, var. costata |
| Q-3-O-(caffeoyl)-sophorotrioside-7-O-glycoside | var. botrytis italica |
| Q-3-O-(sinapoyl)-sophorotrioside-7-O-glycoside | var. botrytis italica |
| Q-3-O-(feruloyl)-sophorotrioside-7-O-glycoside | var. botrytis italica |
| Q-3-O-(p-coumaroyl)-sophorotrioside-7-O-glycoside | var. botrytis italica |
| Q-3-O-(caffeoyl)-sophoroside-7-O-glycoside | var. capitata f. alba, var. acephala, var. botrytis italica |
| Q-3-O-(methoxycaffeoyl)-sophoroside-7-O-glycoside | var. capitata f. alba, var. acephala |
| Q-3-O-(sinapoyl)-sophoroside-7-O-glycoside | var. capitata f. alba, var. acephala, var. botrytis |
| Q-3-O-(p-coumaroyl)-sophoroside-7-O-glycoside | var. botrytis italica |
| Q-3-O-(feruloyl)-sophoroside | var. capitata f. alba, var. acephala, var. botrytis italica |
| K-3-O-tetraglycoside-7-O-sophoroside | var. costata |
| K-3-O-sophorotrioside-7-O-sophoroside | var. capitata f. alba, var. acephala, var. botrytis, var. botrytis italica, var. costata |
| K-3-O-sohorotrioside-7-O-glycoside | var. botrytis, var. botrytis italica, var. costata |
| K-3-O-sophoroside-7-O-diglycoside | var. botrytis, var. botrytis italica, var. costata |
| K-3-O-sophoroside-7-O-glycoside | var. capitata f. alba, var. acephala, var. botrytis, var. botrytis italica, var. costata |
| K-3,7-di-O-glycoside | var. capitata f. alba, var. acephala, var. botrytis italica |
| K-3-O-sophoroside | var. capitata f. alba, var. acephala, var. botrytis italica |
| K-7-O-glycoside | var. capitata f. alba, var. acephala, var. botrytis |
| K-3-O-glycoside | var. botrytis italica, var. costata |
| K-3-O-(caffeoyl)-sophorotrioside-7-O-sophoroside | var. botrytis italica |
| K-3-O-(methoxycaffeoyl)-sophorotrioside-7-O-sophoroside | var. botrytis italica |
| K-O-(sinapoyl)-sophorotrioside-7-O-sophoroside | var. botrytis italica |
| K-O-(feruloyl)-sophorotrioside-7-O-sophoroside | var. botrytis italica |
| K-3-O-(p-coumaroyl)-sophorotrioside-7-O-sophoroside | var. botrytis italica |
| K-3-O-(caffeoyl)-sophorotrioside-7-O-glycoside | var. botrytis italica |
| K-3-O-(methoxycaffeoyl)-sophorotrioside-7-O-glycoside | var. botrytis italica |
| K-O-(sinapoyl)-sophorotrioside-7-O-glycoside | var. botrytis italica |
| K-O-(feruloyl)-sophorotrioside-7-O-glycoside | var. botrytis italica |
| K-3-O-(caffeoyl)sophoroside-7-O-glycoside | var. capitata f. alba, var. acephala, var. botrytis, var. botrytis italica, var. costata |
| K-3-O-(methoxycaffeoyl)-sophoroside-7-O-glycoside | var. capitata f. alba, var. acephala, var. costata |
| K-3-O-(sinapoyl)-sophoroside-7-O-glycoside | var. capitata f. alba, var. acephala, var. botrytis, var. costata |
| K-3-O-(feruloyl)-sophoroside-7-O-glycoside | var. capitata f. alba, var. acephala, var. botrytis, var. costata |
| K-3-O-(p-coumaroyl)-sophoroside-7-O-glycoside | var. capitata f. alba, var. acephala |
| K-3-O-(methoxycaffeoyl)-sophoroside | var. capitata f. alba, var. acephala, var. botrytis italica |
| K-3-O-(sinapoyl)-sophoroside | var. capitata f. alba, var. acephala, var. costata |
| K-3-O-(feruloyl)-sophorotrioside | var. costata |
| K-3-O-(feruloyl)-sophoroside | var. capitata f. alba, var. acephala |
| K-3-O-(p-coumaroyl)-sophoroside | var. capitata f. alba, var. acephala |
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license ( http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mageney, V.; Neugart, S.; Albach, D.C. A Guide to the Variability of Flavonoids in Brassica oleracea. Molecules 2017, 22, 252. https://doi.org/10.3390/molecules22020252
Mageney V, Neugart S, Albach DC. A Guide to the Variability of Flavonoids in Brassica oleracea. Molecules. 2017; 22(2):252. https://doi.org/10.3390/molecules22020252
Chicago/Turabian StyleMageney, Vera, Susanne Neugart, and Dirk C. Albach. 2017. "A Guide to the Variability of Flavonoids in Brassica oleracea" Molecules 22, no. 2: 252. https://doi.org/10.3390/molecules22020252