Biochemical, Physiological and Transcriptomic Comparison between Burley and Flue-Cured Tobacco Seedlings in Relation to Carbohydrates and Nitrate Content

Abstract
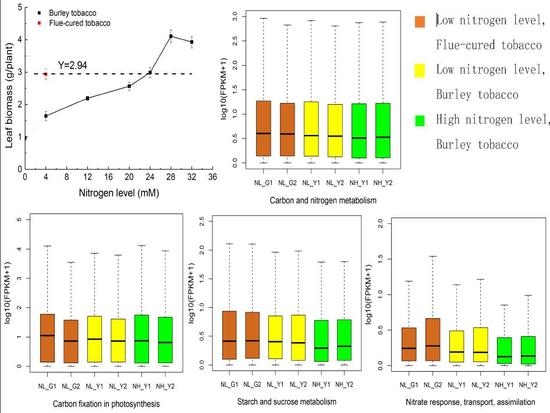
:
1. Introduction
2. Results
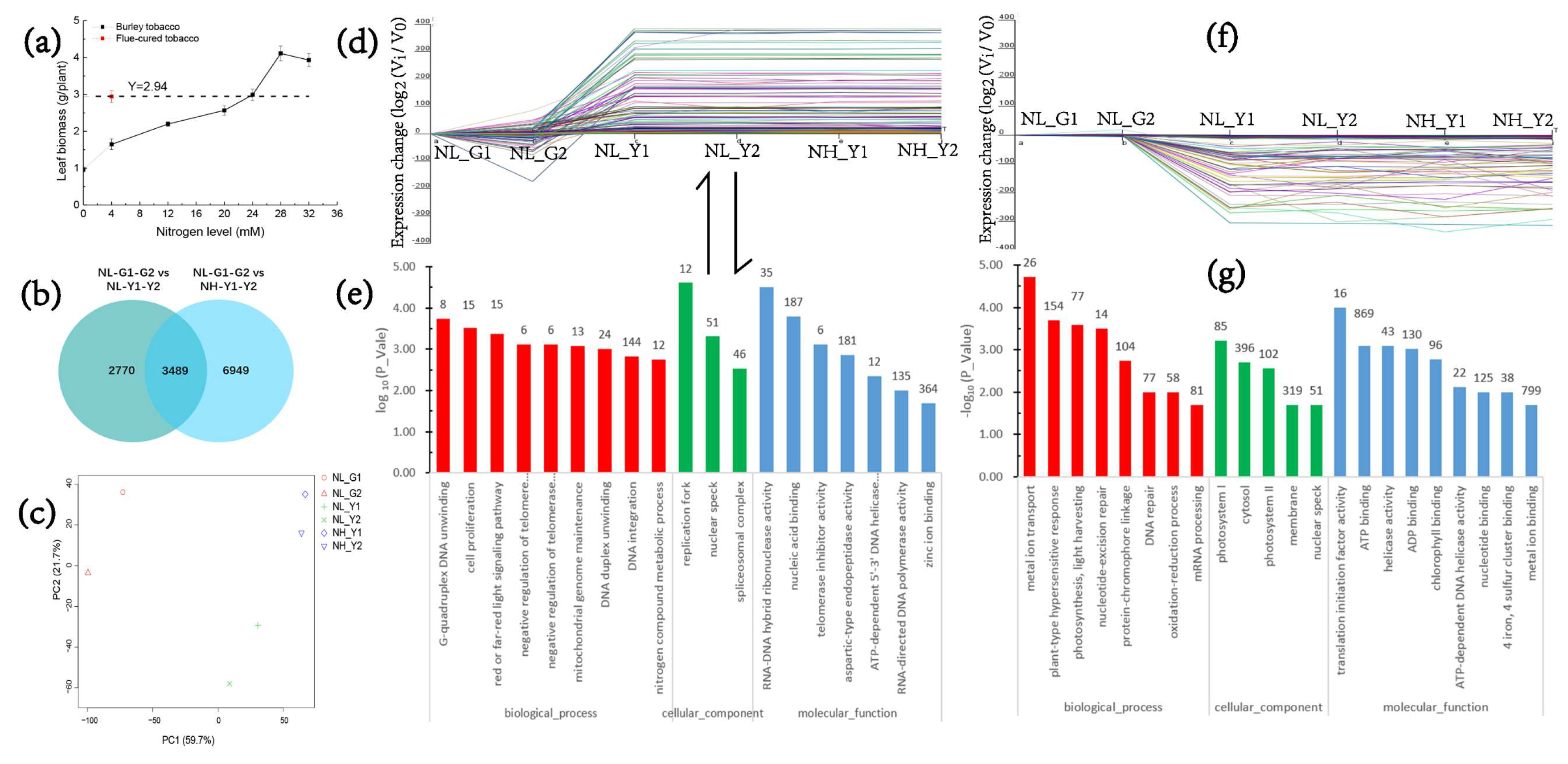
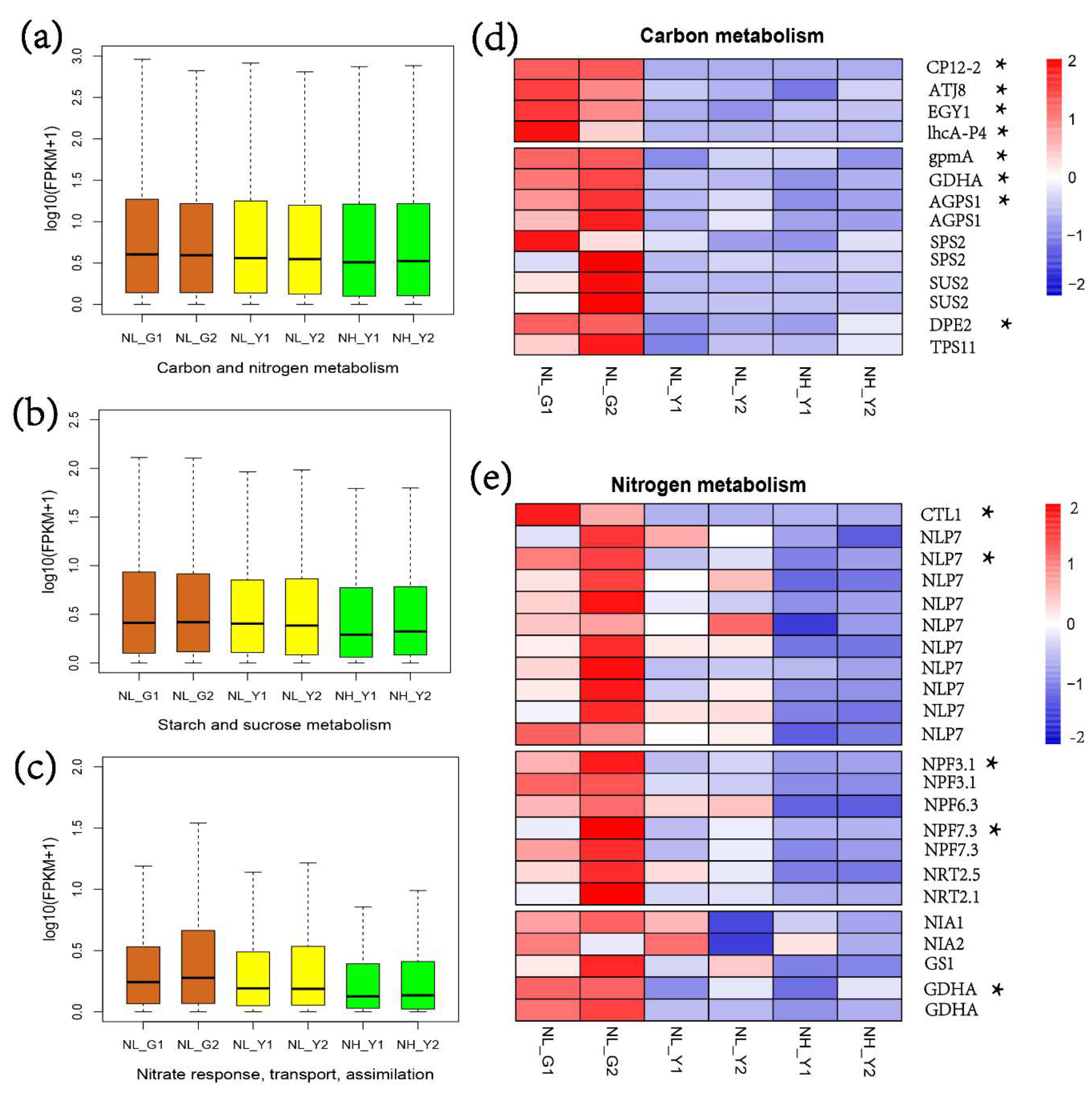
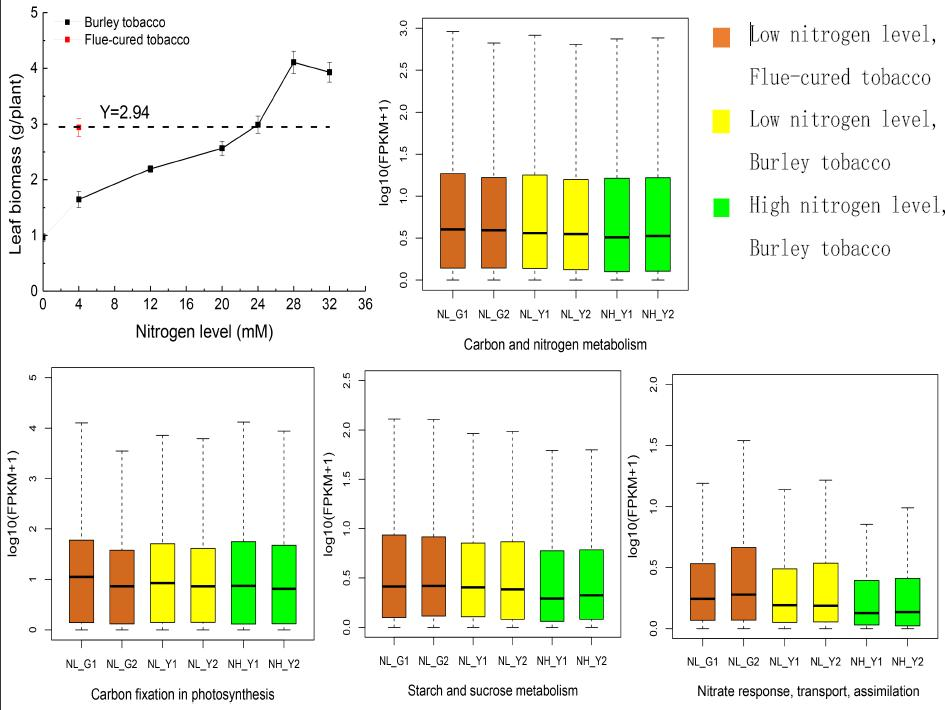
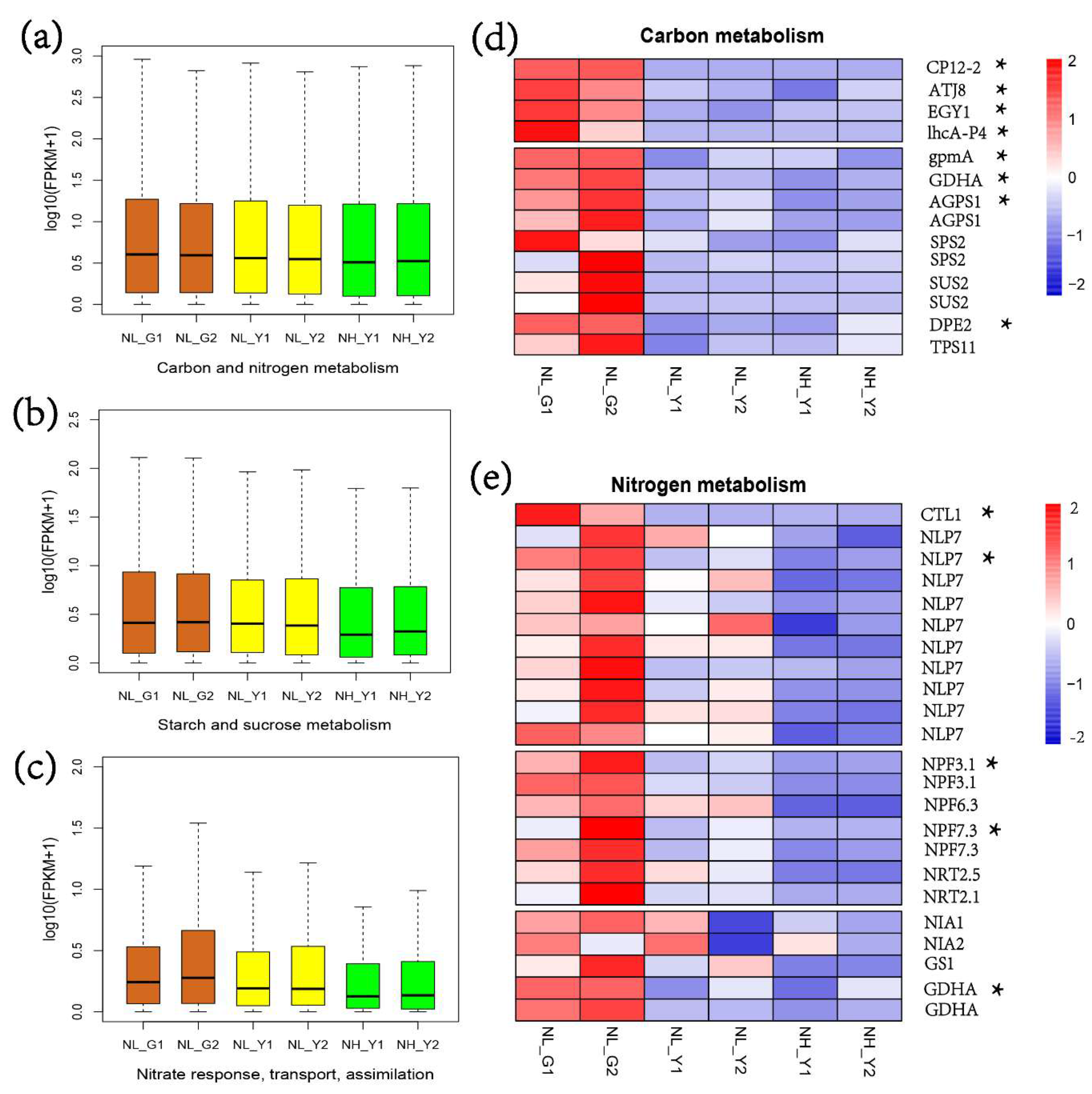
2.1. RNA-Seq Statistics, and Molecular Analysis on Nitrogen and Carbon Metabolism
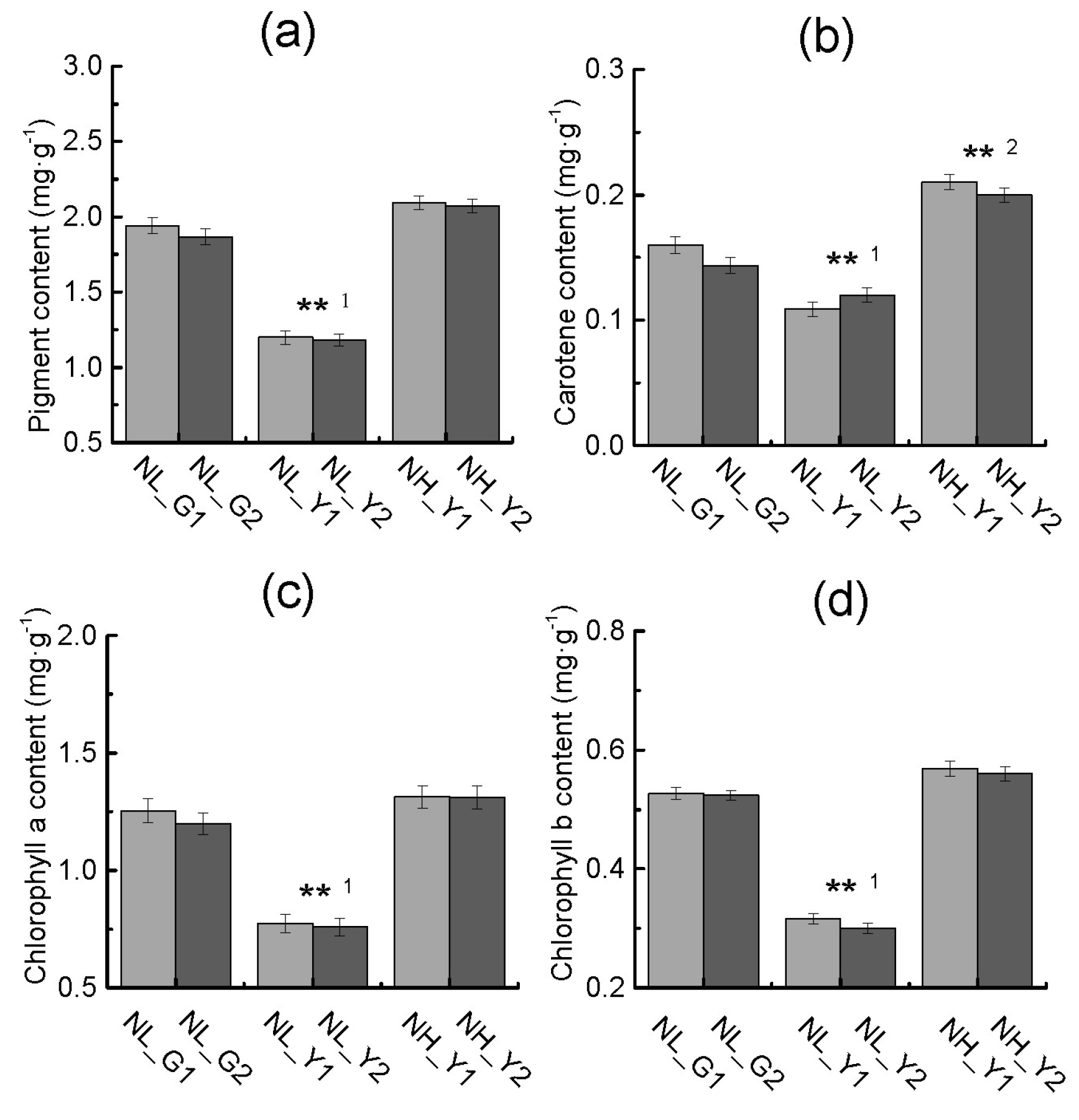
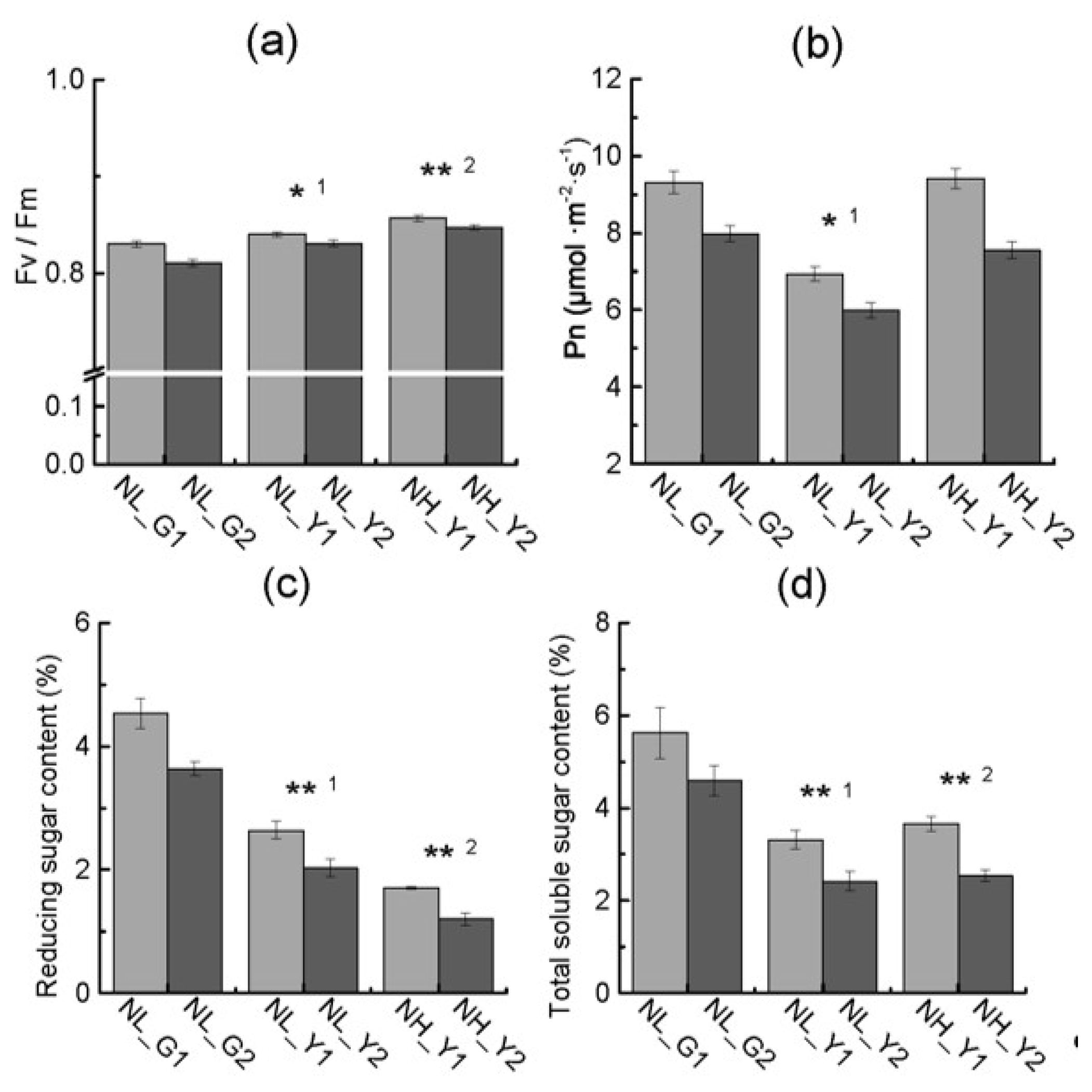
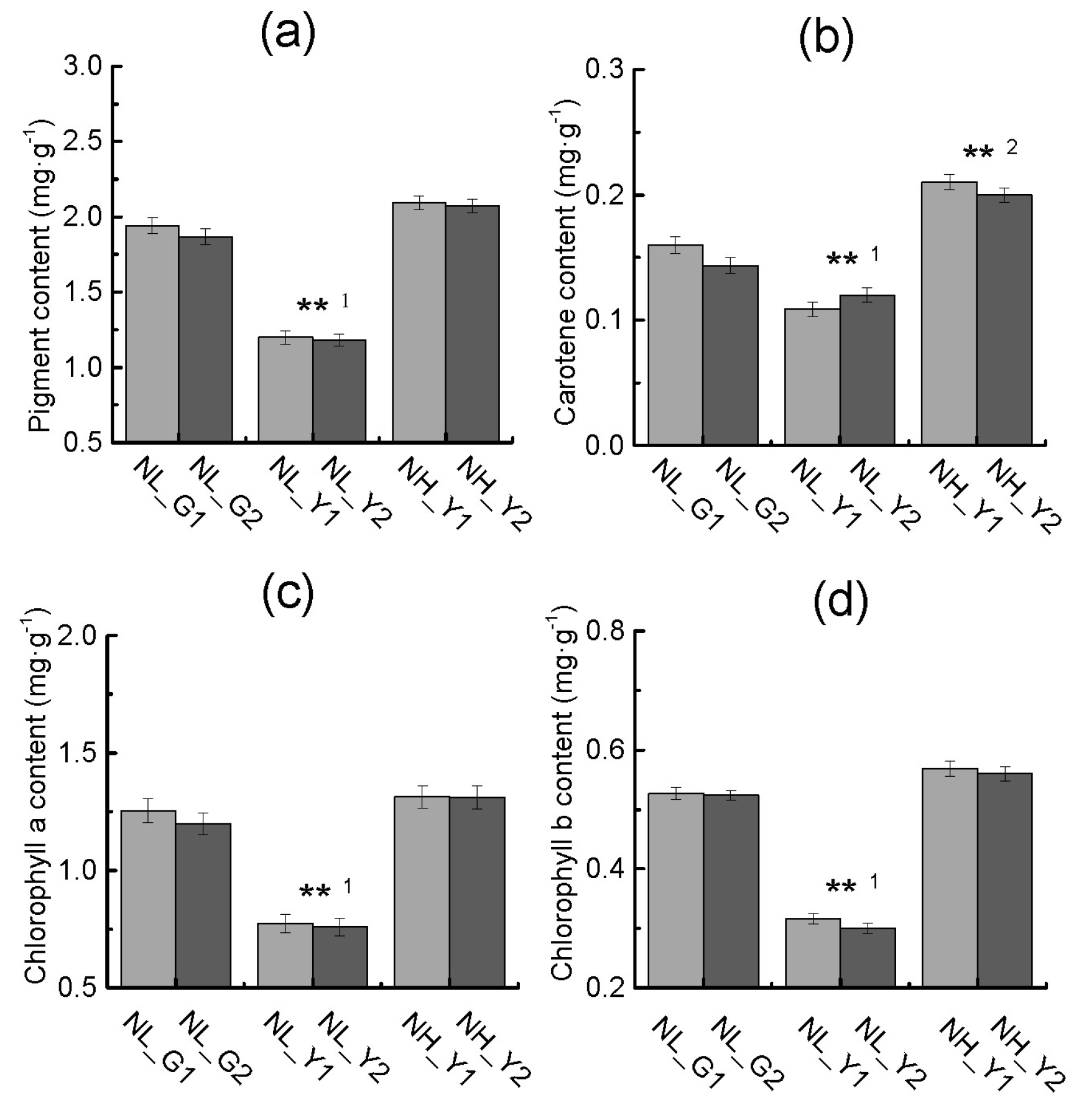
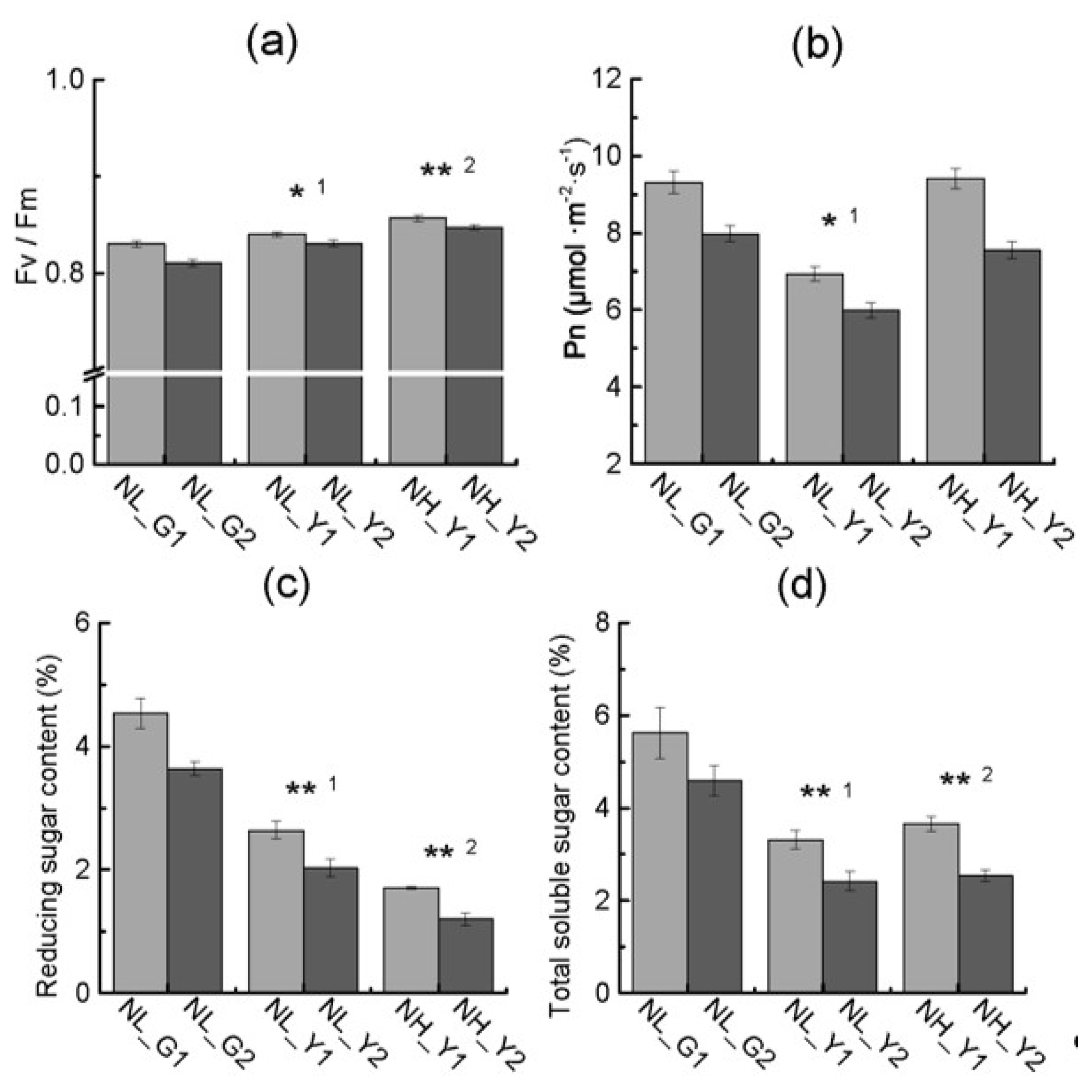
2.2. Differences in Pigment Content and Photosynthesis between the Two Types
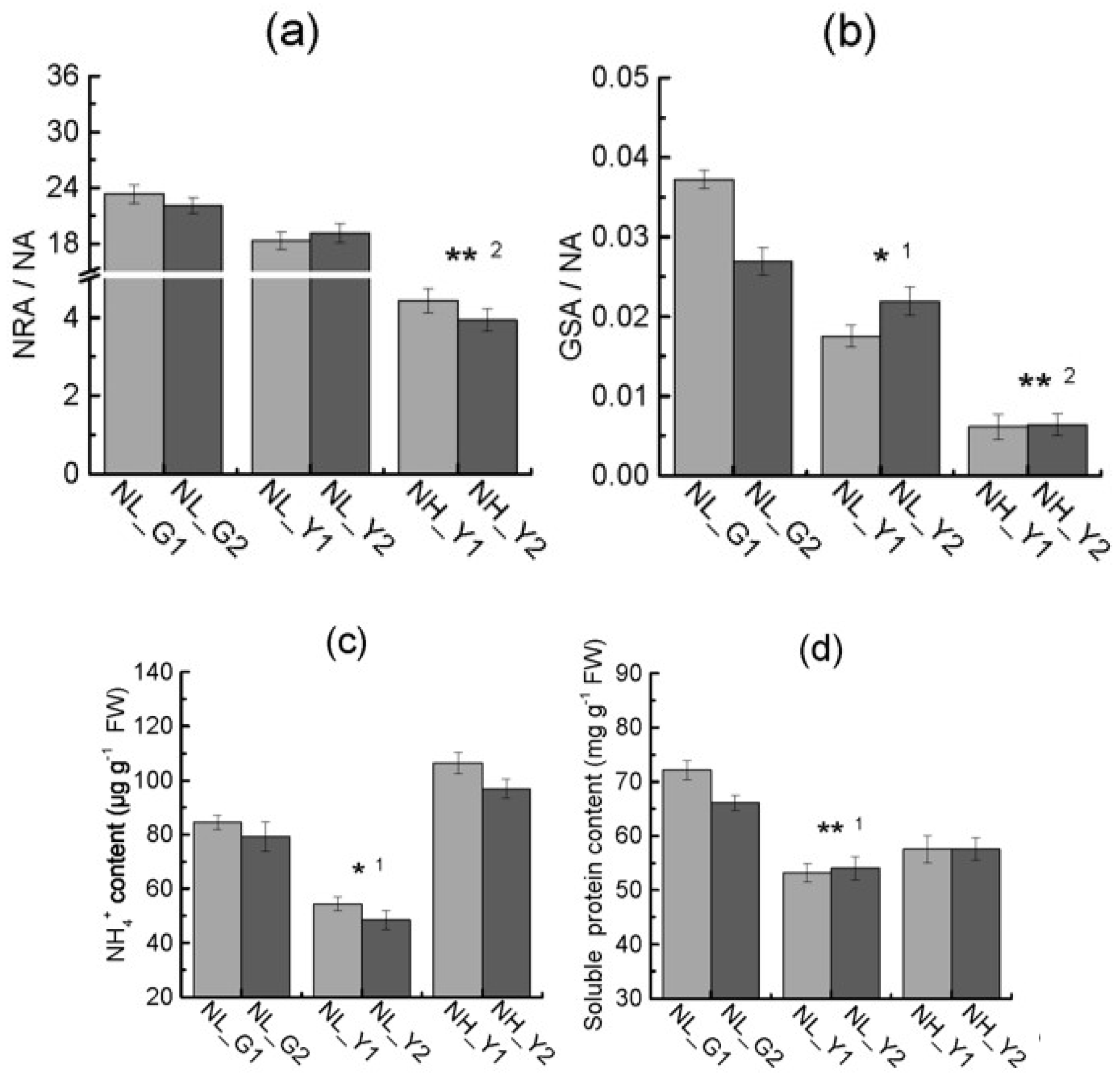
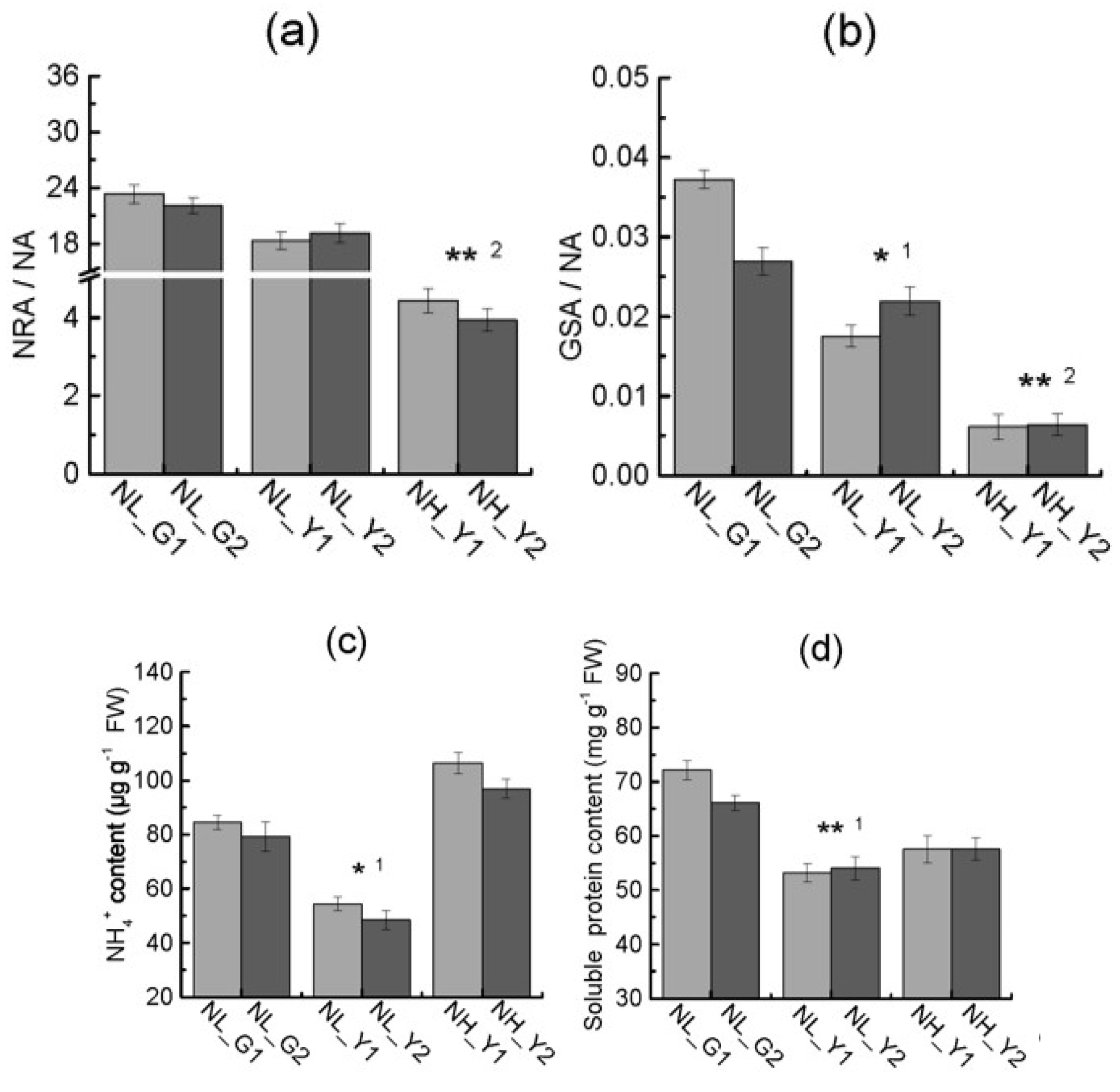
2.3. Differences in Enzymes Activities Correlated with Nitrogen Metabolism
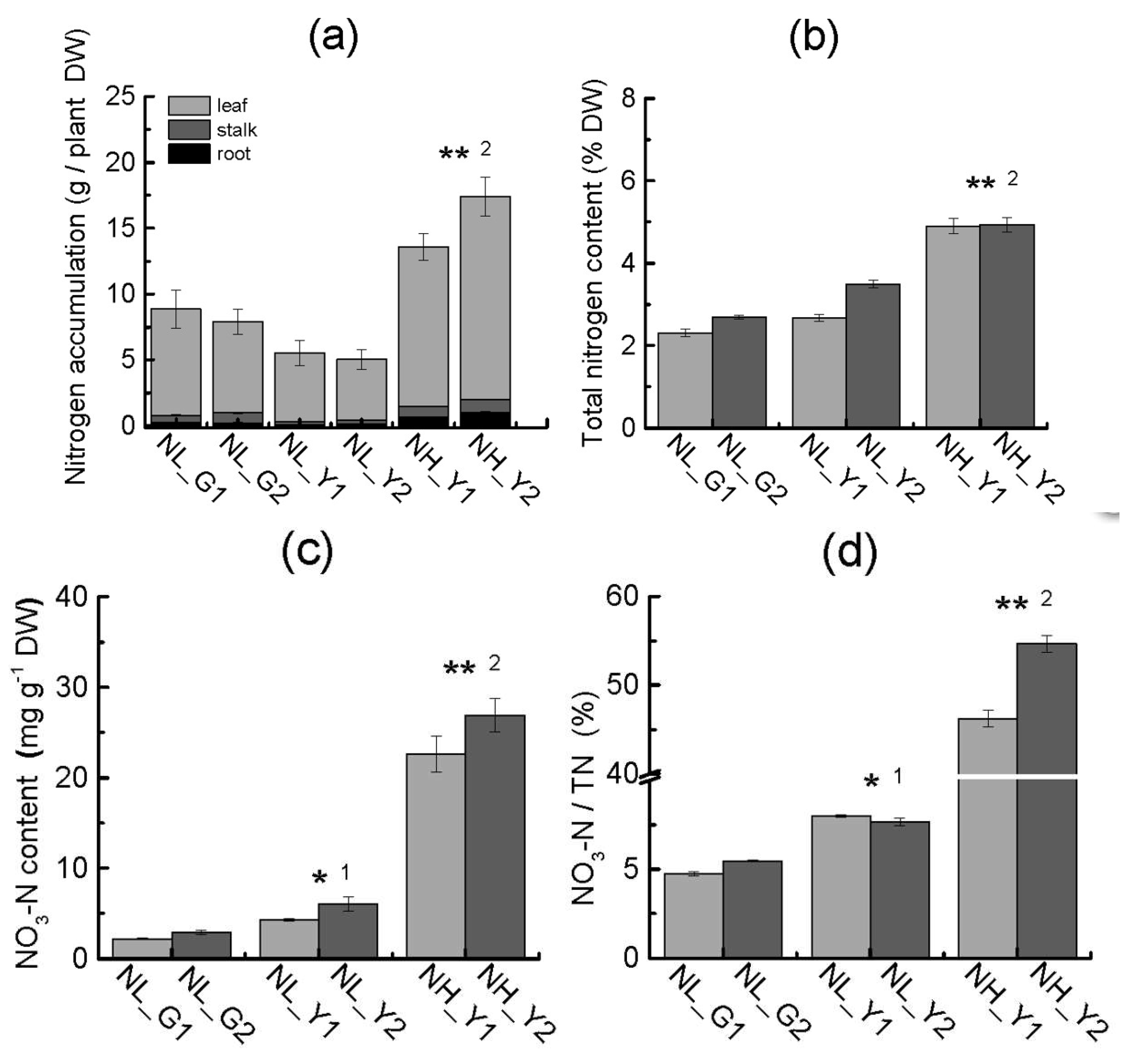
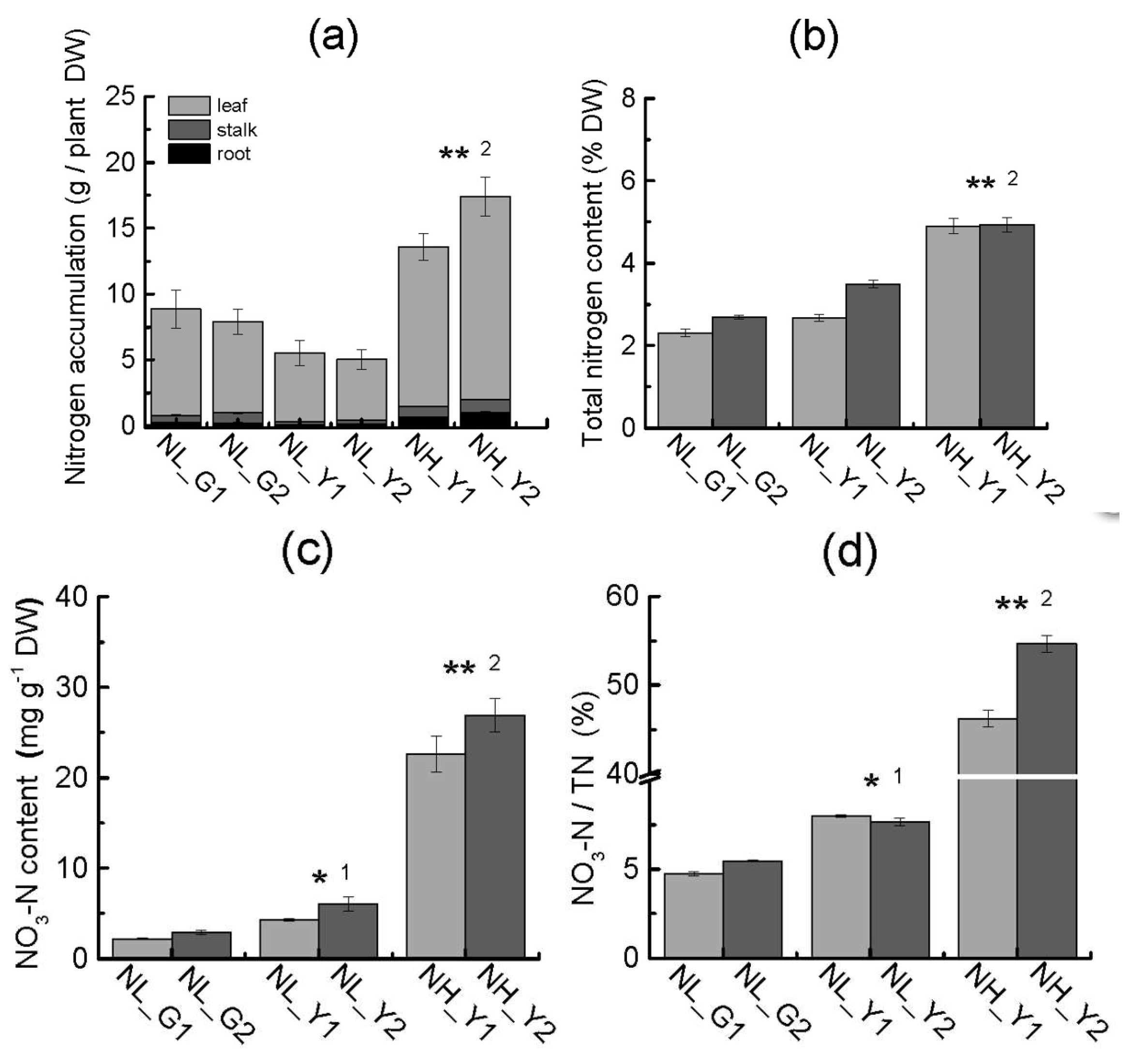
2.4. Differences in Carbon and Nitrogen Compounds between Flue-Cured and Burley Tobacco
2.5. Correlation Analysis
3. Discussion
4 Material and Methods
4.1. Plant Material and Growth Conditions
4.2. Treatments
4.3. Sampling
4.4. Measurement of NRA, GSA and Nitrate
4.5. Measurement of Pigment Content, Photosynthetic Rate (Pn), Chlorophyll a Fluorescence
4.6. Measurement of Total Nitrogen, Total Soluble Sugar, Reducing Sugar Content
4.7. RNA Extraction, Preparation of cDNA Library, and Sequencing
4.8. RNA-Seq Analysis, GO and KEGG Pathway Enrichment Analysis of Differentially Expressed Genes (DEGs)
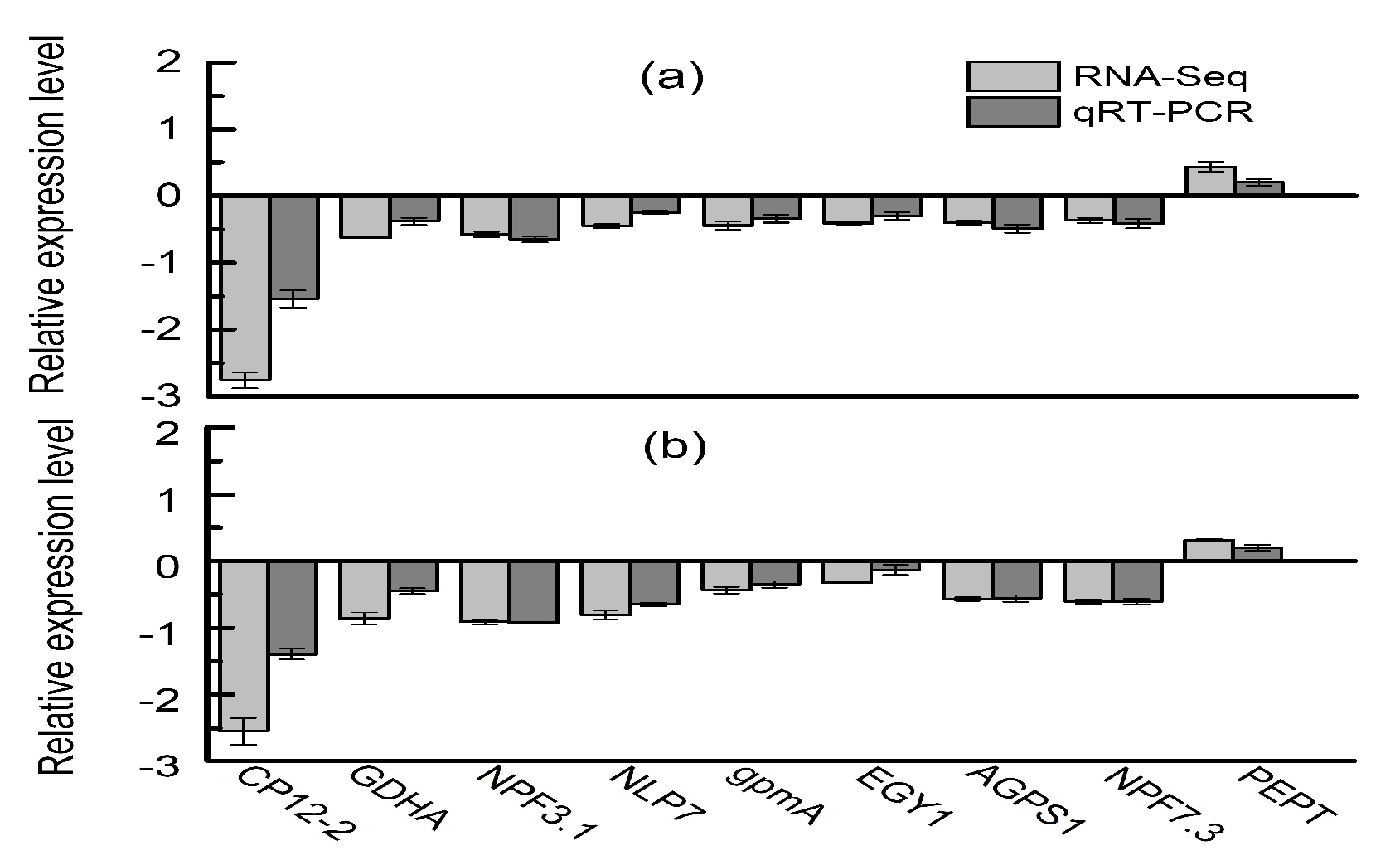
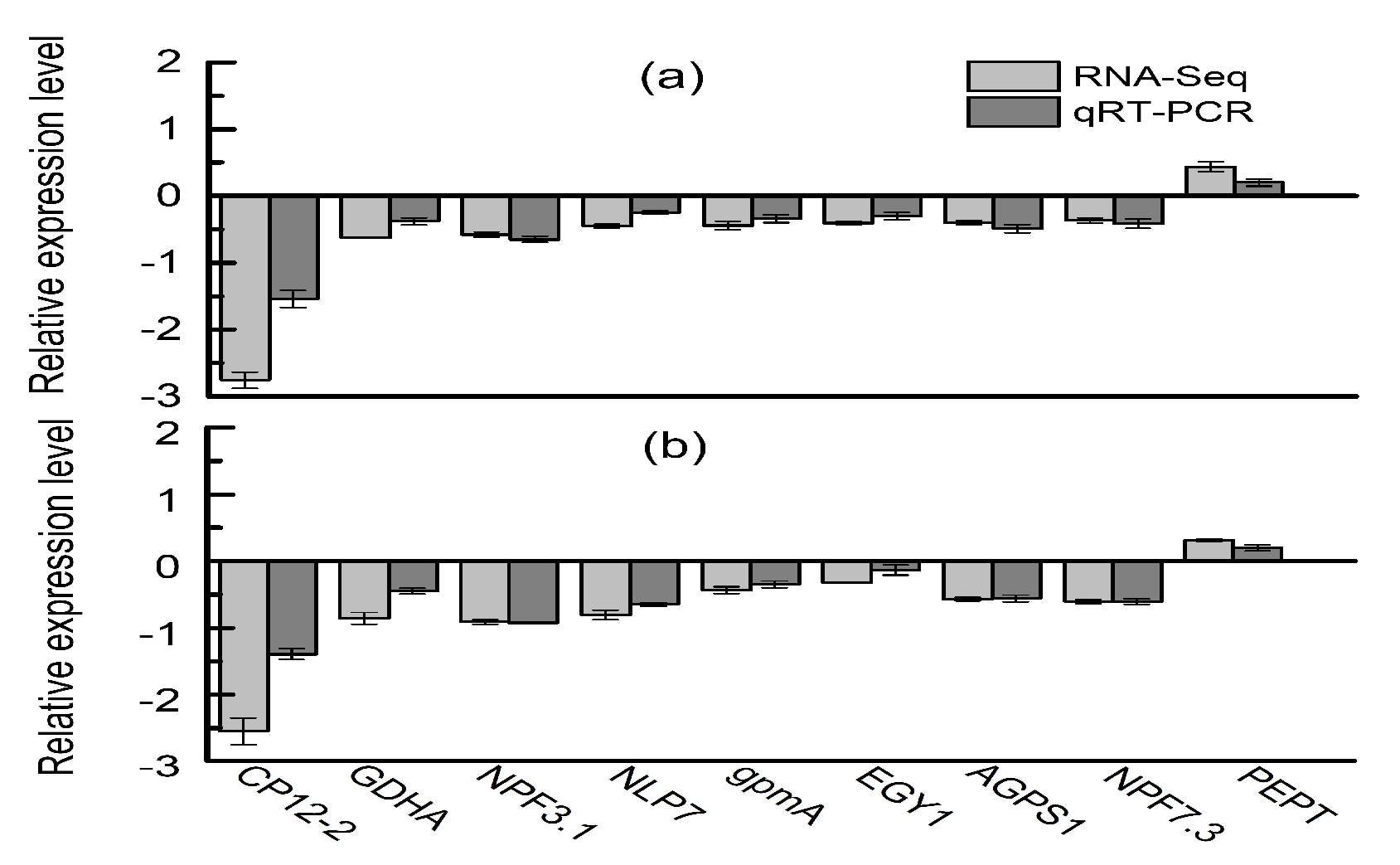
4.9. Validation by qRT-PCR Analysis
4.10. Statistical Analysis
Supplementary Materials
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Henica, F.S. The inheritance of the White Burley character in tobacco. Jpn. J. Crop Sci. 1932, 4, 281–282. [Google Scholar]
- Yang, J.; Tian, Y.C.; Yao, X.; Cao, W.X.; Zhang, Y.S.; Zhu, Y. Hyperspectra estimation model for chlorophyll concentra tions in top leaves of rice. Acta Ecol. Sin. 2009, 29, 6561–6571. [Google Scholar]
- Shang, Z.Q. Effects of nitrogen amount on growth and development yield and quality in burley tobacco. Chin. Agric. Sci. Bull. 2007, 23, 299–301. [Google Scholar]
- Sun, W.S.; Wang, J.; Zhou, J.; Ma, Y.J.; Yang, H.J.; Xu, D.Y.; Bai, R.S.; Jiao, Z.H.; Shi, H.Z. Effect of nitrate nitrogen level in tobacco leaves on TSNAs formation during high temperature storage. Acta Tabacaria Sin. 2015, 21, 53–58. [Google Scholar]
- Reddy, K.S.; Menary, R.C. Nitrate reductase and nitrate accumulation in relation to nitrate toxicity in Boronia megastigma. Physiol. Plant. 1990, 78, 430–434. [Google Scholar] [CrossRef]
- Santamaria, P. Nitrate in vegetables: Toxicity, content, intake and EC regulation. A review. J. Sci. Food Agric. 2006, 86, 10–17. [Google Scholar] [CrossRef]
- Burton, H.; Dye, N.; Bush, L. Relationship between tobacco-specific nitrosamines and nitrite from different air-cured tobacco varieties. J. Agric. Food Chem. 1994, 42, 2007–2011. [Google Scholar] [CrossRef]
- Shi, H.Z.; Wang, R.Y.; Bush, L.P. The relationships between TSNAs and their precursors in burley tobacco from different regions and varieties. J. Food Agric. Environ. 2012, 10, 132–136. [Google Scholar]
- Lewis, R.S.; Parker, R.G.; Danehower, D.A.; Andres, K.; Jack, A.M.; Whitley, D.S.; Bush, L.P. Impact of Alleles at the Yellow Burley (Yb) Loci and Nitrogen Fertilization Rate on Nitrogen Utilization Efficiency and Tobacco-Specific Nitrosamine (TSNA) Formation in Air-Cured Tobacco. J. Agric. Food Chem. 2012, 60, 6454–6461. [Google Scholar] [CrossRef] [PubMed]
- Vieira, I.S.; Vasconcelos, E.P.; Monteiro, A.O.A. Nitrate accumulation, yield and leaf quality of turnip greens in response to nitrogen fertilization. Nutr. Cycl. Agroecosyst. 1998, 51, 249–258. [Google Scholar] [CrossRef]
- Burns, I.G.; Zhang, K.; Turner, M.K.; Meacham, M.; Al-Redhiman, K.; Lynn, J.; Broadley, M.R.; Hand, P.; Pink, D. Screening for genotype and environment effects on nitrate accumulation in 24 species of young lettuce. J. Sci. Food Agric. 2011, 91, 553–562. [Google Scholar] [CrossRef] [PubMed]
- Tang, Y.F.; Sun, X.C.; Hu, C.X. Genotypic differences in nitrate uptake, translocation and assimilation of two Chinese cabbage cultivars [Brassica campestris L. ssp. Chinensis (L.). Plant Physiol. Biochem. 2013, 70, 14–20. [Google Scholar] [CrossRef] [PubMed]
- Man, H.M.; Boriel, R.; E1-Khatib, R.; Kirby, E.G. Characterization of transgenic poplar with ectopic expression of pine cytosolic glutamine synthetase under conditions of varying nitrogen availability. New Phytol. 2005, 167, 31–39. [Google Scholar] [CrossRef] [PubMed]
- Börner, T.; Mendel, R.R.; Schiemann, J. Nitrate reductase is not accumulated in chloroplast-ribosome-deficient mutants of higher plants. Planta 1986, 169, 202–207. [Google Scholar] [CrossRef] [PubMed]
- Wallsgrove, R.M.; Lea, P.J.; Miflin, B.J. Distribution of the enzymes of nitrogen assimilation within the pea leaf cell. Plant Physiol. 1979, 63, 232–236. [Google Scholar] [CrossRef] [PubMed]
- Du, J.; Shu, S.; Shao, Q.S. Mitigative effects of spermidine on photosynthesis and carbon nitrogen balance of cucumber seedlings under Ca(NO3)2 stress. J. Plant Res. 2016, 129, 79–91. [Google Scholar] [CrossRef] [PubMed]
- Groben, R.; Kaloudas, D.; Raines, C.A.; Offmann, B.; Maberly, S.C.; Gontero, B. Comparative sequence analysis of CP12, a small protein involved in the formation of a Calvin cycle complex in photosynthetic organisms. Photosynth. Res. 2010, 103, 183–194. [Google Scholar] [CrossRef] [PubMed]
- Chen, G.; Yu, R.B.; Li, N. EGY1 encodes a membrane-associated and ATP-independent metalloprotease that is required for chloroplast development. Plant J. 2004, 41, 364–375. [Google Scholar] [CrossRef] [PubMed]
- Chen, K.M.; Holmstrom, M.; Raksajit, W.; Suorsa, M.; Piippo, M.; Aro, E.M. Small chloroplasttargeted DnaJ proteins are involved in optimization of photosynthetic reactions in Arabidopsis thaliana. BMC Plant Biol. 2010, 10. [Google Scholar] [CrossRef] [PubMed]
- Jansson, S. A guide to the Lhc genes and their relatives in Arabidopsis. Trends Plant Sci. 1999, 4, 236–240. [Google Scholar] [CrossRef]
- Chourey, P.; Taliercio, E.; Carlson, S.; Ruan, Y.L. Genetic evidence that the two isozymes of sucrose synthase present in developing maize endosperm are critical one for cell wall integrity and the other for starch biosynthesis. Mol. Gen. Genet. 1998, 259, 88–96. [Google Scholar] [PubMed]
- Chen, S.; Hajirezaei, M.; Börnke, F. Differential expression of sucrose-phosphate synthase isoenzymes in tobacco reflects their functional specialization during dark-governed starch mobilization in source leaves. Plant Physiol. 2005, 139, 1163–1174. [Google Scholar] [CrossRef] [PubMed]
- Volkert, K.; Debast, S.; Voll, L.M.; Voll, H.; Schie, I.; Hofmann, J.; Schneider, S.; Börnke, F. Loss of the two major leaf isoforms of sucrose-phosphate synthase in Arabidopsis thaliana limits sucrose synthesis and nocturnal starch degradation but does not alter carbon partitioning during photosynthesis. J. Exp. Bot. 2014, 65, 5217–5229. [Google Scholar] [CrossRef] [PubMed]
- Munoz, F.J.; Zorzano, M.T.M.; Alonso-Casajús, N.; Baroja-Fernandez, E.; Etxeberria, E.; Pozueta-Romero, J. New enzymes, new pathways and an alternative view on starch biosynthesis in both photosynthetic and heterotrophic tissues of plants. Biocatal. Biotrans. 2006, 24, 63–76. [Google Scholar] [CrossRef]
- Sun, W.S.; Yang, J.J.; Zhou, J.; Xu, D.; Bai, R.; Ma, Y.; Zhang, G.; Shi, H. Effect of different nitrogen forms on nitrate nitrogen content and TSNAs formation in tobacco. Acta Tabacaria Sin. 2015, 21, 78–84. [Google Scholar]
- Zhong, R.Q.; Kays, S.; Schroeder, B.; Ye, Z. Mutation of a chitinase-like gene causes ectopic deposition of lignin, aberrant cell shapes, and overproduction of ethylene. Plant Cell 2002, 14, 165–179. [Google Scholar] [CrossRef] [PubMed]
- Ai, S.Y.; Yao, J.W.; Huang, X.H.; Luo, W.; Ke, Y.; Ling, D. Study on the nitrate reduction characteristic of vegetables. Plant Nutr. Fertil. Sci. 2002, 8, 40–43. [Google Scholar]
- Lin, S.H.; Kuo, H.F.; Canivenc, G.; Lin, C.S.; Lepetit, M.; Hsu, P.K.; Tillard, P.; Lin, H.L.; Wang, Y.Y.; Tsai, C.B.; et al. Mutation of the Arabidopsis NRT1.5 nitrate transporter causes defective root-to-shoot nitrate transport. Plant Cell 2008, 20, 2514–2528. [Google Scholar] [CrossRef] [PubMed]
- Li, W.; Wang, Y.; Okamoto, M.; Crawford, N.M.; Siddiqi, M.Y.; Glass, A.D. Dissection of the AtNRT2.1: AtNRT2.2 inducible high-affinity nitrate transporter gene cluster. Plant Physiol. 2007, 143, 425–433. [Google Scholar] [CrossRef] [PubMed]
- Chen, B.M.; Wang, Z.H.; Li, S.X. Effects of nitrate supply on plant growth, nitrate accumulation, metabolic nitrate concentration and nitrate reductase activity in three leafy vegetables. Plant Sci. 2004, 167, 635–643. [Google Scholar] [CrossRef]
- Yanagisawa, S. Transcription factors involved in controlling the expression of nitrate reductase genes in higher plants. Plant Sci. 2014, 229, 167–171. [Google Scholar] [CrossRef] [PubMed]
- Yu, L.H.; Wu, J.; Tang, H.; Yuan, Y.; Wang, S.M.; Wang, Y.P.; Zhu, Q.S.; Li, S.G.; Xiang, C.B. Overexpression of Arabidopsis NLP7 improves plant growth under both nitrogen-limiting and -sufficient conditions by enhancing nitrogen and carbon assimilation. Sci. Rep. 2016, 6, 27795. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.F.; Chang, D.; Sun, J.; Yang, H.J.; Wang, J.; Shi, H.Z. Difference of nitrogen metabolism between flue-cured tobacco and burley tobacco seedlings. Tob. Sci. Technol. 2017, 50, 6–11. [Google Scholar]
- Li, H.S.; Sun, Q.; Zhao, S.J.; Zhang, W.H. Principle and Technology of Plant Physiological and Biochemical Experiments; Higher Education Press: Beijing, China, 2000. [Google Scholar]
- O’Neal, D.; Joy, K. Glutamine synthetase of pea leaves. I. Purification, stabilization, and pH optima. Plant Physiol. 1974, 54, 773–779. [Google Scholar] [CrossRef]
- Cataldo, D.A.; Haroon, M.; Schrader, L.E.; Youngs, V.L. Rapid cplorimetric Deternination of Nitrate in Plant-Tissure by Nitration of Salicylic Acid. Commun. Soil Sci. Plant Anal. 1975, 6, 71–80. [Google Scholar] [CrossRef]
- Wintermans, J.F.G.M.; De Mots, A. Spectrophotometric characteristics of chlorophylls a and b and their pheophytins in ethanol. Biochim. Biophys. Acta 1965, 109, 448–453. [Google Scholar] [CrossRef]
- Liu, Y.F.; Han, X.R.; Zhan, X.M.; Yang, J.F.; Wang, Y.Z.; Song, Q.B.; Chen, X. Regulation of Calcium on Peanut Photosynthesis Under Low Night Temperature Stress. J. Integr. Agric. 2013, 12, 2172–2178. [Google Scholar] [CrossRef]
- Schreiber, U.; Bilger, W.; Neubauer, C. Chlorophyll Fluorescence as a Nonintrusive Indicator for Rapid Assessment of in Vivo Photosynthesis. In Ecophysiology of Photosynthesis; Schulze, E.-D., Caldwell, M.M., Eds.; Springer Study Edition: Berlin, Heidelberg, 1995; pp. 49–70. [Google Scholar]
- Patel, R.K.; Jain, M. NGS QC Toolkit: A Toolkit for Quality Control of Next Generation Sequencing Data. PLoS ONE 2012, 7, e30619. [Google Scholar] [CrossRef] [PubMed]
- Langmead, B.; Salzberg, S.L. Fast gapped-read alignment with Bowtie 2. Nat. Methods 2012, 9, 357–359. [Google Scholar] [CrossRef] [PubMed]
- Kim, D.; Pertea, G.; Trapnell, C.; Pimentel, H.; Kelley, R.; Salzberg, S.L. TopHat2: Accurate alignment of transcriptomes in the presence of insertions, deletions and gene fusions. Genome Biol. 2013, 14. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Trapnell, C.; Roberts, A.; Goff, L.; Pertea, G.; Kim, D.; Kelley, D.R.; Pimentel, H.; Salzberg, S.L.; Rinn, J.L.; Pachter, L. Differential gene and transcript expression analysis of RNA-seq experiments with TopHat and Cufflinks. Nat. Protoc. 2012, 7, 562–578. [Google Scholar] [CrossRef] [PubMed]
- Anders, S.; Pyl, P.T.; Huber, W. HTSeq-a Python framework to work with high-throughput sequencing data. Bioinformatics 2015, 31, 166–169. [Google Scholar] [CrossRef] [PubMed]
- Anders, S.; Huber, W. Differential Expression of RNA-Seq Data at the Gene Level—The DESeq Package; EMBL: Heidelberg, Germany, 2013. [Google Scholar]
- Koonin, E.V.; Fedorova, N.D.; Jackson, J.D.; Jacobs, A.R.; Krylov, D.M.; Makarova, K.S.; Mazumder, R.; Mekhedov, S.L.; Nikolskaya, A.N.; Rao, B.S.; et al. A comprehensive evolutionary classification of proteins encoded in complete eukaryotic genomes. Genome Biol. 2004, 5. [Google Scholar] [CrossRef] [PubMed]
- Apweiler, R.; Bairoch, A.; Wu, C.H.; Barker, W.C.; Boeckmann, B.; Ferro, S.; Gasteiger, E.; Huang, H.Z.; Lopez, R.; Magrane, M.; et al. UniProt: The Universal Protein knowledgebase. Nucleic Acids Res. 2004, 32, D115–D119. [Google Scholar] [CrossRef] [PubMed]
- Kanehisa, M.; Goto, S.; Kawashima, S.; Okuno, Y.; Hattori, M. The KEGG resource for deciphering the genome. Nucleic Acids Res. 2004, 32, D277–D280. [Google Scholar] [CrossRef] [PubMed]
- Ashburner, M.; Ball, C.A.; Blake, J.A.; David, B.; Butler, H.; Cherry, J.M.; Davis, A.P.; Dolinski, K.; Dwight, S.S.; Eppig, J.T.; et al. Gene Ontology: Tool for the unification of biology. Nat. Genet. 2000, 25, 25–29. [Google Scholar] [CrossRef] [PubMed]
- Ernst, J.; Bar-Joseph, Z. STEM: A tool for the analysis of short time series gene expression data. BMC Bioinform. 2006, 7. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Q.; Guo, J.J.; He, C.T.; Shen, C.; Huang, Y.Y.; Chen, J.X.; Guo, J.H.; Yuan, J.G.; Yang, Z.Y. Comparative Transcriptome Analysis between Low- and High-Cadmium-Accumulating Genotypes of Pakchoi (Brassica chinensis L.) in Response to Cadmium Stress. Environ. Sci. Technol. 2016, 50, 6485–6494. [Google Scholar] [CrossRef] [PubMed]
- Britton, N.F.; Lin, X.H.; Safer, H.M.; Schneider, M.V.; Singh, M.; Tramontano, A. RNA-Seq Data Analysis-A Practical Approach; CRC press: London, UK, 2014. [Google Scholar]
- Jin, J.J.; Sun, Y.W.; Qu, J.; Syah, R.; Lim, C.; Alfiko, Y.; Rahman, N.; Suwanto, A.; Yue, G.H.; Wong, L.; et al. Transcriptome and functional analysis reveals hybrid vigor for oil biosynthesis in oil palm. Sci. Rep. 2017, 7, 439. [Google Scholar] [CrossRef] [PubMed]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 22DDCT method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]
Sample Availability: Samples of the compounds NL_G1, NL_G2, NL_Y1 and NH_Y1 are available from the authors. |

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Parameter | NRA | GSA | NO3-N | TN | NO3-N/TN | Pigment | Pn |
|---|---|---|---|---|---|---|---|
| Total soluble sugar content | 0.362 | 0.292 | −0.452 | −0.554 | −0.486 | 0.433 | 0.777 |
| Reducing sugar content | −0.119 | −0.086 | −0.805 | −0.879 * | −0.825 * | 0.019 | 0.368 |
| Leaf biomass | 0.631 | 0.796 | 0.226 | 0.014 | 0.201 | 0.822 * | 0.738 |
| Parameter | NRA/NA | GSA/NA | Pigment/NA | Pn/NA |
|---|---|---|---|---|
| NO3-N | −0.986 ** | −0.828 * | −0.923 ** | −0.970 ** |
| NO3-N/TN | −0.989 ** | −0.849 * | −0.936 ** | −0.973 ** |
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, Y.; Yang, H.; Chang, D.; Lin, S.; Feng, Y.; Li, J.; Shi, H. Biochemical, Physiological and Transcriptomic Comparison between Burley and Flue-Cured Tobacco Seedlings in Relation to Carbohydrates and Nitrate Content. Molecules 2017, 22, 2126. https://doi.org/10.3390/molecules22122126
Li Y, Yang H, Chang D, Lin S, Feng Y, Li J, Shi H. Biochemical, Physiological and Transcriptomic Comparison between Burley and Flue-Cured Tobacco Seedlings in Relation to Carbohydrates and Nitrate Content. Molecules. 2017; 22(12):2126. https://doi.org/10.3390/molecules22122126
Chicago/Turabian StyleLi, Yafei, Huijuan Yang, Dong Chang, Shuzhen Lin, Yuqing Feng, Jingjing Li, and Hongzhi Shi. 2017. "Biochemical, Physiological and Transcriptomic Comparison between Burley and Flue-Cured Tobacco Seedlings in Relation to Carbohydrates and Nitrate Content" Molecules 22, no. 12: 2126. https://doi.org/10.3390/molecules22122126