Neurotoxic Effects of Linalool and β-Pinene on Tribolium castaneum Herbst


1
Environmental and Computational Chemistry Group, Zaragocilla Campus, School of Pharmaceutical Sciences, University of Cartagena, Cartagena 130016, Bolivar, Colombia
2
Medical and Pharmaceutical Sciences Group, School of Health Sciences, Department of Medicine, University of Sucre, Sincelejo 700003, Sucre, Colombia
*
Author to whom correspondence should be addressed.
Molecules 2017, 22(12), 2052; https://doi.org/10.3390/molecules22122052
Submission received: 9 September 2017
/
Accepted: 21 November 2017
/
Published: 24 November 2017
Abstract
:Effective, ethical pest control requires the use of chemicals that are highly specific, safe, and ecofriendly. Linalool and β-pinene occur naturally as major constituents of the essential oils of many plant species distributed throughout the world, and thus meet these requirements. These monoterpenes were tested as repellents against Tribolium castaneum, using the area preference method, after four hours of exposure and the effect transcriptional of genes associated with neurotransmission. Changes in gene expression of acetylcholinesterase (Ace1), GABA-gated anion channel splice variant 3a6a (Rdl), GABA-gated ion channel (Grd), glutamate-gated chloride channel (Glucl), and histamine-gated chloride channel 2 (Hiscl2) were assessed and the interaction with proteins important for the insect using in silico methods was also studied. For linalool and β-pinene, the repellent concentration 50 (RC50) values were 0.11 µL/cm2 and 0.03 µL/cm2, respectively. Both compounds induced overexpression of Hiscl2 gen in adult insects, and β-pinene also promoted the overexpression of Grd and the Ace1 gene. However, β-pinene and linalool had little potential to dock on computer-generated models for GABA-gated ion channel LCCH3, nicotinic acetylcholine receptor subunits alpha1 and alpha2, and putative octopamine/tyramine receptor proteins from T. castaneum as their respective binding affinities were marginal, and therefore the repellent action probably involved mechanisms other than direct interaction with these targets. Results indicated that β-pinene was more potent than linalool in inducing insect repellency, and also had a greater capacity to generate changes in the expression of genes involved in neuronal transmission.
1. Introduction
Biopesticides are gaining increased attention and interest among those concerned with environmentally friendly, safe, and integrated crop management approaches [1,2] and, correspondingly, nature has provided us with a variety of chemotherapeutic agents [3]. Among them, essential oils (EOs) and their constituents affect biochemical processes, specifically disrupting the endocrinological balance of insects. In fact, some plants biosynthesize EOs to protect themselves from insects and may disrupt the process of morphogenesis through neurotoxicity, or by acting as growth regulators [1] and interfering with the basic metabolic and physiology of insects [4].
Terpenes are a class of plant secondary metabolites that have a distinct structure and function, and are considered to be important agents in the medicinal uses of aromatic plants [3]. Monoterpenes, a structurally diverse group of phytochemical compounds, are the major constituent of EOs. In recent years, interest in understanding the pharmacological actions of these molecules has increased. Studies have demonstrated that these naturally occurring molecules can modulate the functional properties of various types of voltage and ligand-gated ion channels, and thus can be used as alternatives to synthetic insecticides against pests on stored products [5].
Linalool and β-pinene are two alcohol and hydrocarbon volatile monoterpenes [6] reported to be major components of EOs in various aromatic species [7] (Figure 1). Linalool is produced by plants and is used in cosmetic products, perfumes, and flavorings. Furthermore, it has been shown to have antimicrobial activity and insect-repellant properties that act on the central nervous system (CNS) [6]. Linalool is slightly volatile with a pleasant aroma associated with lavender and laurel fragrances [3,8]. The aim of this study was to evaluate the effects of linalool and β-pinene in the expression of genes related to neurotransmission on T. castaneum, and assesses the interaction with proteins important for the insect using in silico methods.
2. Results
2.1. Repellent Activity
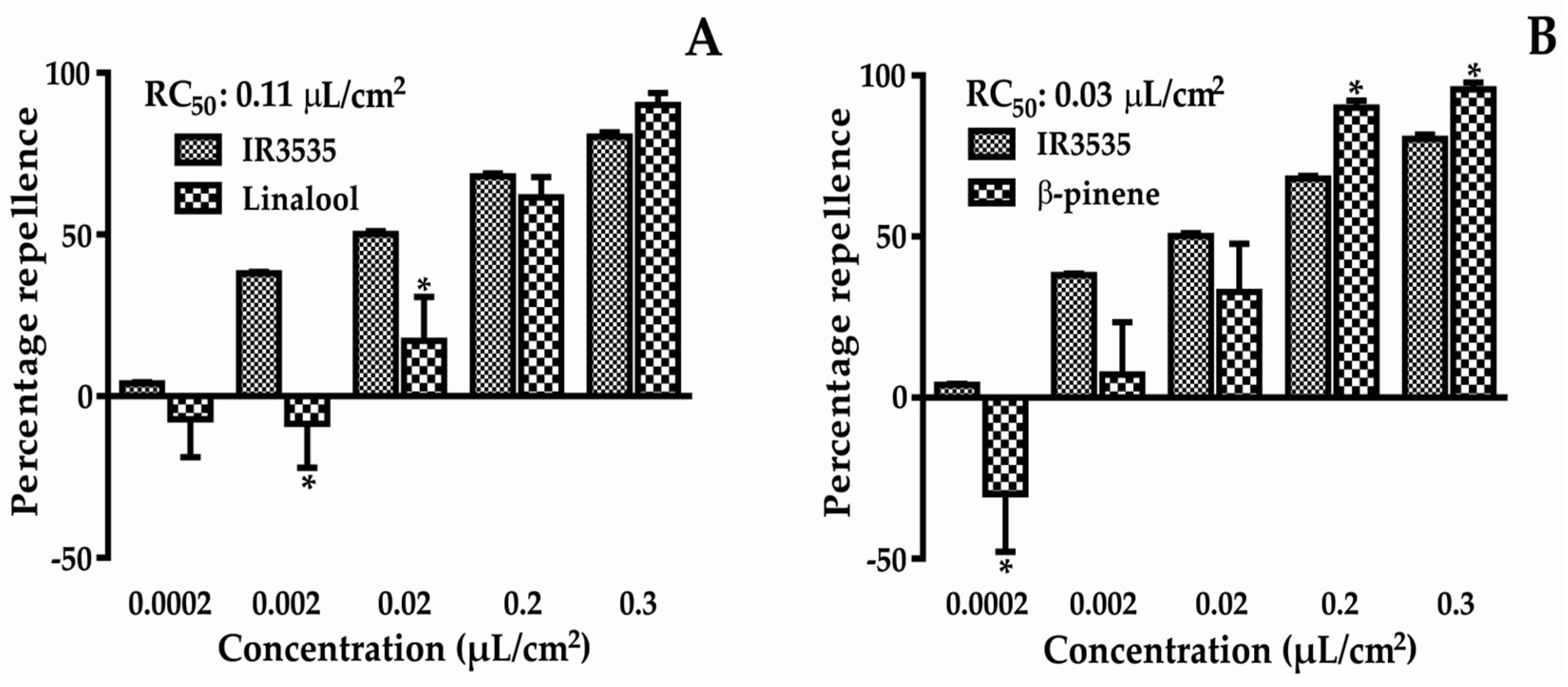
The percentage repellency results from the chemicals tested are presented in Figure 2. At the lowest concentration, attractive activity was found for both compounds. However, at higher concentrations, these compounds showed strong repellent activity, with repellent concentration 50 (RC50) values of 0.11 and 0.03 µL/cm2 for linalool and β-pinene, respectively. The RC50 of the positive control was 0.02 µL/cm2. The two-way ANOVA results showed that there was interaction between the different concentrations evaluated, therefore, the repellent effect was dependent on the concentration.
2.2. Gene Expression
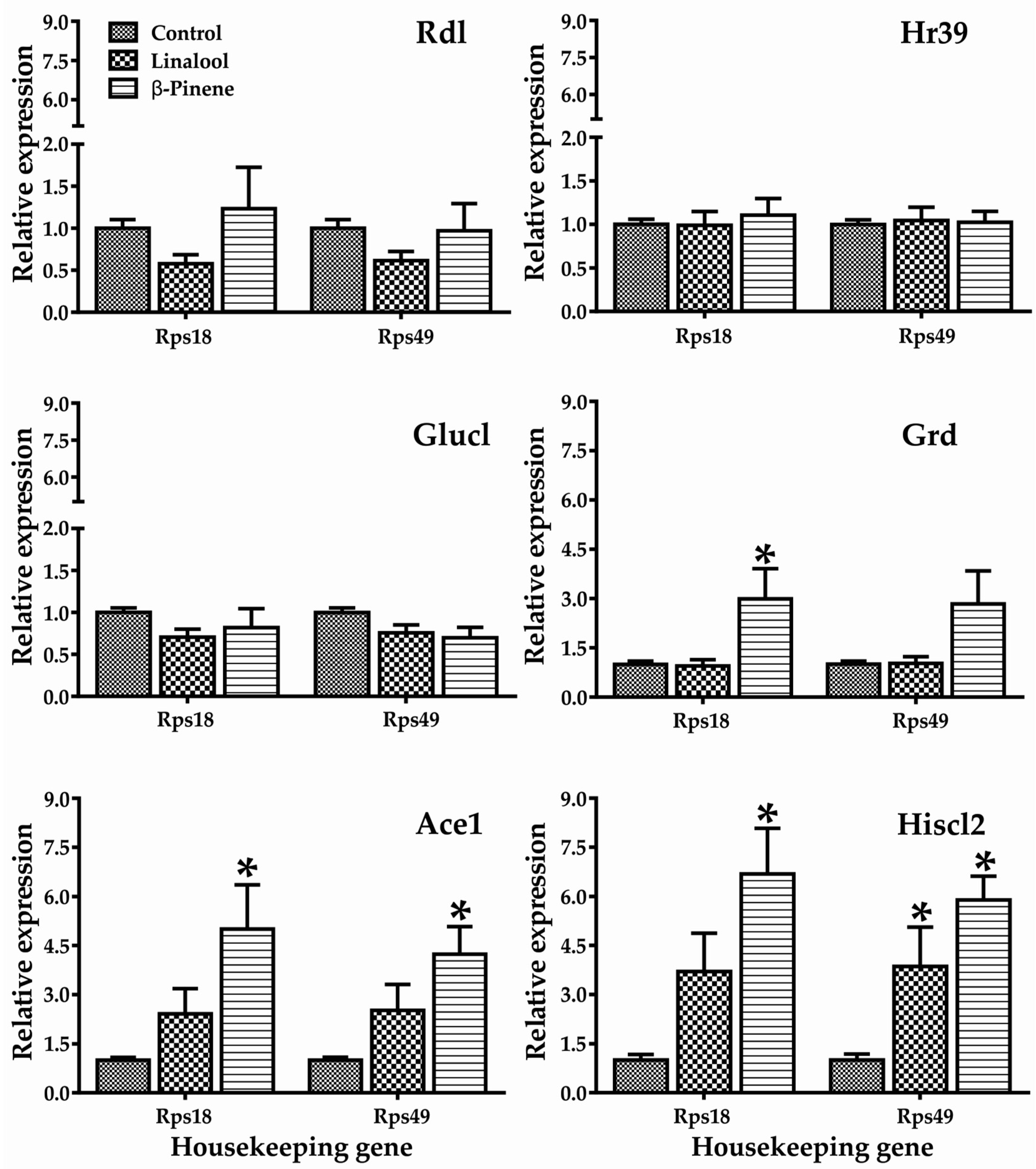
In this study, changes in the gene expression of T. castaneum adults are presented in Figure 3. Both constituents behaved differently, and showed statistically significant differences in the expression of genes such as Grd, Ace1, and Hiscl2. Although insects exposed to linalool did not show significant differences in the expression of the gene Ace1, a tendency to increase was observed in both housekeeping genes. The two-way ANOVA showed that there was no interaction between the genes, therefore, the gene expression was independent for each gene.
2.3. Homology Modeling and Validation
The best proteins obtained in the homology modeling process were loaded in Protein Model Data Base (PMDB). The validation results obtained using the different servers demonstrated the quality of the models. ProQ rated the obtained models as fairly, very, and extremely good (Table 1).
The results of the validation process obtained by RAMPAGE showed that the studied proteins had a number of residues in favored and allowed regions of over 90% (Figures S1–S13). Figures S14–S26 depict the results obtained using QMean, which were a measure of the degree of nativity of a structure for a particular protein. These results showed values with a negative sign, indicating scores lower than those obtained for the average experimental structures.
2.4. Molecular Docking
A computational approach was carried out to identify possible targets for linalool and β-pinene, with a particular focus on the T. castaneum proteins involved in neurological responses. The results are presented in Table 2. β-pinene produced better interactions than linalool, with affinity values ranging from −7.7 to −5.0 Kcal/mol. However, these affinity values were low when compared to those obtained from known inhibitors.
The 3D structure of the proteins and amino acids involved in the protein-ligand interaction are shown in Figure S27. Linalool and β-pinene bind to the same site and therefore interact with the same type of amino acids in all the studied complexes, except those formed with the carboxylic ester hydrolase and the glutamate-gated chloride channel proteins where the chemicals were coupled on different sites of the protein. The major amino acids identified in the protein-ligand interactions were ILE, LEU, THR, SER, and VAL, with a frequency of occurrence greater than 40%. The main protein–ligand interactions identified were hydrophobic and hydrogen bonding.
3. Discussion
Natural insect repellent products have become a viable alternative to synthetic repellents [12,13] as they provide less risk to the environment and to human health [7]. In this paper, two main EO compounds were studied to evaluate repellent activity, gene expression, and molecular docking. β-pinene showed greater repellent activity than linalool (Figure 2), and both had smaller RC50 values than IR3535, although these were comparable with the positive control.
β-pinene was more potent against T. castaneum than linalool. This compound is an oxygenated monoterpene which is abundantly present in the environment, and is widely found in plants as a constituent of EOs [14]. It has been reported with acute toxicity to insects of T. castaneum after three days of exposure [15]. Meanwhile, linalool is a monoterpene of 10 carbon atoms and an alcohol group [16] that has been reported as a mosquito (Aedes aegypti) [17] and beetle repellent (T. castaneum and Dominica Rhyzopertha) [18], which coincides with the effects observed in this study.
Linalool and β-pinene are insect repellents; however, the mechanisms by which these compounds function are not fully understood. It is only known that monoterpenes can act on various insect and mammalian receptors, especially at the level of the nervous system, and specifically on gamma-aminobutyric acid (GABA)-gated chloride channels; octopamine, tyramine, and nicotinic acetylcholine (nAChR) receptors; acetylcholine esterase; sodium channels; and other targets. Several different monoterpenoids have been reported to bind to ionotropic GABA receptors in humans, rodents, and insects in previous research [1,5,19]. Allelochemical defense systems in insects allow the rapid elimination of ingested toxic substances, and/or the fat bodies, therefore facilitating the detoxification of contaminants penetrating the cuticular or tracheal structures through enzymes—such as P450, Glutathione-S-transferases, and esterases—which are typically concentrated in the insect midgut [1]. However, some of the most important targets of insecticidal agents are inhibitory neurotransmitter receptors such as invertebrate gamma-aminobutyric acid (GABA)-gated chloride channels (GABACls) and glutamate-gated chloride channels (GluCls), which are related to changes in behavior [20].
Repellent activity is related to changes in the behavior of insects, which are caused by effects on the CNS. The histamine-gated ion channel is a member of the cys-loop ligand-gated ion channel (cys-loop LGIC) superfamily [21], and has two subunits—HisCl1 and HisCl2—that control the phototransduction of invertebrates [22]. Outside of phototransduction, histamine also appears to be the neurotransmitter found in some mechanosensory neurons in Drosophila as well as in lobster stomatogastric, cardiac, and olfactory neurons [23,24]. Adult insects exposed to linalool and β-pinene overexpressed Hiscl2 gen (Figure 3), most likely suggesting that the function of this gene could be one of the mechanisms involved in the repellent action of these monoterpenes. However, these compounds have little in silico ability to bind to this protein, with affinity values of −5.0 ± 0.2 and −5.7 ± 0.0 Kcal/mol for linalool and β-pinene, respectively (Table 2), leaving room for the involvement of modulatory mechanisms not related to direct binding.
Monoterpenes are capable of quickly penetrating insect cells as they are volatile and lipophilic compounds that interfere with physiological functions so their modes of action or mechanisms of resistance are especially complex [1,5]. For instance, they interact with the functioning of GABA synapses [25], or change the activity of voltage gated and/or ligand-gated ion channels in the CNS, thus impairing the activity of neuronal enzymes such as Ace [26].
Compared to β-pinene, linalool exhibited less activity in terms of gene expression, despite some potential to interact with proteins involved in synapsis (Figure S27b). On the other hand, β-pinene induces overexpression of Grd and Ace1 in T. castaneum adults (Figure 3). The ionotropic receptors of T. castaneum function as GABA-gated chloride channels (GABACls-Grd), receptor members of the family of the Cys-loop ligand-gated ion channels, which mediate fast inhibitory synaptic transmission [22,27]. On the other hand, the canonical biological function of Ace is to terminate impulse transmission at cholinergic synapses by rapidly hydrolyzing the neurotransmitter acetylcholine in animals; thus, it is the target of anticholinesterase insecticides [28]. Although it is clear that the evaluated monoterpenoids had little capacity to directly interact in silico with several proteins related to neurotransmission (Table 2, Figure S27), the activation of gene expression indicated that other alternative mechanisms were indirectly modulating their repellent activities.
The use of molecular tools together with in silico approaches assists in gaining insight into toxicity mechanisms [29,30,31]. In this investigation, in addition to evaluating gene expression, the interactions of two monoterpenes with important proteins in neurotransmission were also studied. Homology modeling was employed to construct the three-dimensional structure of proteins prior to molecular docking. The quality of the models was evaluated with QMean, ProQ, and RAMPAGE (Table 1, Figures S1–S26), and LGscore and MaxSub values implied that the obtained models were good [32]. The QMean Z-score uses structures that are solved by X-ray crystallography as a reference to estimate the absolute quality of a model [33]. For the models in this study, the QMean Z-score was in the range of −10.99 to −4.70, showing that these models varied in quality; conversely, however, the results obtained with RAMPAGE and ProQ suggested that the models had the appropriate quality [34,35,36].
The application of computational approaches is often used for the study of the toxicological profiles of new candidate insecticidal agents [37]. The docking results revealed that linalool and β-pinene had little ability to interact with the T. castaneum proteins involved in the neurotransmission process (Table 2) as the binding affinity were between −5.0 and 6.0 Kcal/mol, values far from those obtained for known inhibitors, which are around −9.0 Kcal/mol (Table 2) [9,10,11]. Interestingly, protein–ligand interactions for β-pinene and linalool also shared some residues with Ivermectin [38]; suggesting a common binding site. Although the evaluated proteins were related to the action of several chemical compounds on insects, in this in silico study, linalool and β-pinene showed very low affinity values on the proteins, indicating that repellent action probably involved alternative mechanisms.
4. Materials and Methods
4.1. Insect Rearing
The wild-type strain of Tribolium castaneum Herbst was taken from a stock colony maintained in our laboratory. The identification of T. castaneum was performed according to the morphological characteristics reported by Dönitz et al. [39]. Insects were reared on a diet of flakes and ground oats (70:30) at 26 ± 2 °C, with a 70% to 85% relative humidity and a 10:14 h light:dark photoperiod [2,6,7]. Then, two to four week old healthy adults were randomly chosen for bioassays.
4.2. Materials
Linalool (>99%) and β-pinene (>95%) were purchased from Sigma-Aldrich (Steinheim, Germany). Acetone reagent (≥99.5%) was also purchased from Sigma-Aldrich.
4.3. Repellent Activity
The experiments were carried out on adults of T. castaneum under laboratory conditions. The repellent activity of linalool and β-pinene was evaluated using the area preference method [20,40]. Test areas consisted of 9 cm Albet DP 597125 filter paper cut in half (31.8 cm2). As experimental containers, Petri dishes (9 cm in diameter) were used to house 20 T. castaneum. Linalool and β-pinene were serially dissolved in acetone (0.0002, 0.002, 0.02, 0.2 and 0.3 µL/cm2). Five hundred µL of the solution were uniformly applied to a half-filter paper disc to five different testing concentrations. The other half of the remaining filter paper was treated with 500 µL of acetone and used as the control vehicle. Then, the half discs were air-dried for about 10 min to allow solvent evaporation. During each test, 20 T. castaneum adults without regard to gender were released at the center of the disk, then covered quickly with a dish cover and placed in darkness. Counts of the insects present on each side of the filter paper were made after 4 h of exposure. As a positive control, a 15% formulation of IR3535 ethyl 3-(N-acetyl-N-butylamino)propionate [2] was used. The percent repellency (PR) of each compound/commercial repellent was then calculated using the formula Equation (1).
where Nc and Nt are the number of insects on the untreated (control) and treated areas, respectively. Three replicates were used for each tested concentration of the main compound, and each assay was repeated twice.
PR = [(Nc − Nt)/(Nc + Nt)] × 100
After the experiment was conducted, the median repellent concentration (RC50) for each compound was obtained by Probit analysis (RC50 = 0.11 µL/cm2 for linalool, and RC50 = 0.03 µL/cm2 for β-pinene) and was used for gene expression assays.
4.4. Gene Expression Assays
Adult control and exposed organisms were directly frozen with liquid nitrogen and preserved in RNAlater. Total RNA was isolated using an RNeasy Mini Kit (Qiagen, Valencia, CA, USA) according to the manufacturer’s protocols. RNA concentrations were measured using a NanoDrop 2000 spectrophotometer (Thermo Scientific, Wilmington, DE, USA) and the quality was verified by absorbance ratio 260/280 nm. cDNA was obtained through the QuantiTect® Reverse Transcription Kit (Qiagen Inc., Valencia, CA, USA) according to the manufacturer’s protocols. A real-time polymerase chain reaction (RT-PCR) was performed on a StepOne Plus real-time PCR system (Applied Biosystems, Foster City, CA, USA) apparatus. Six genes were analyzed (Table 3) [41,42,43]. Gene expression was normalized to ribosomal protein 18 (Rps18) and ribosomal protein 49 (Rps49) [44]. Primer sequences are presented in Table 3. RT-PCR was performed with a Maxima SYBR Green/ROX qPCR Master Mix (Thermo Scientific, Waltham, MA, USA). The ΔΔCT was used to calculate relative gene expression. All experiments were run in duplicate [45,46,47].
4.5. Homology Modeling and Validation
The amino acid sequences of the proteins from T. castaneum were obtained from Uniprot [48] in FASTA format with the following access numbers E7DN61, E7DN62, A8DMU1, A8DMU2, A8DMU3, A8DMU5, A8DMU7, A8DMU8, D2A6H1, A8DIP3, A8DIQ7, A8DMU9, and D6WB14. Three-dimensional structures of carboxylic ester hydrolase, carboxylic ester hydrolase 2, gamma-aminobutyric acid (GABA)-RDL, GABA-GRD, GABA-gated ion channel LCCH3, glutamate-gated chloride channel, histamine-gated chloride channel 1, histamine-gated chloride channel 2, hormone receptor in 39-like, nicotinic acetylcholine receptor subunit alpha1, nicotinic acetylcholine receptor subunit alpha2, pH sensitive chloride channel, and putative octopamine/tyramine receptor proteins were constructed by homology modeling using (PS)2-v2: Protein Structure Prediction Server [49,50], Phyre2 [51], and I-Tasser [52]. The best model generated by each prediction server was selected to validate the model quality [33]. The quality of the models was evaluated using the RAMPAGE [53], ProQ (Protein Quality Predictor), and QMEAN [31,54]. The best structure of each protein obtained after the validation procedure was subjected to structural analysis using the molecular graphics system PyMOL [33] and saved in the Protein Model DataBase (PMDB) [32]. PM0081096, PM0081123, PM0081100, PM0081098, PM0081126, PM0081097, PM0081124, PM0081099, PM0081101, PM0081125, PM0081129, PM0081127, and PM0081128 were assigned as the identifiers for the structures carboxylic ester hydrolase, carboxylic ester hydrolase2, GABA-RDL, GABA-GRD, GABA-gated ion channel LCCH3, glutamate-gated chloride channel, histamine-gated chloride channel 1, histamine-gated chloride channel 2, hormone receptor in 39-like, nicotinic acetylcholine receptor subunit alpha1, nicotinic acetylcholine receptor subunit alpha2, pH sensitive chloride channel, and putative octopamine/tyramine receptor, respectively.
4.6. Molecular Docking
Prior to molecular docking, the 3D structures of carboxylic ester hydrolase, carboxylic ester hydrolase 2, GABA-RDL, GABA-GRD, GABA-gated ion channel LCCH3, glutamate-gated chloride channel, histamine-gated chloride channel 1, histamine-gated chloride channel 2, hormone receptor in 39-like, nicotinic acetylcholine receptor subunit alpha1, nicotinic acetylcholine receptor subunit alpha2, pH sensitive chloride channel, and putative octopamine/tyramine receptor proteins from T. castaneum were subsequently optimized and minimized using atomic partial charges with the Kollman method using the SYBYL 8.1.1 package (Tripos, San Luis, MO, USA). MGLTools 1.5.0 software (Molecular Graphics Laboratory, La Jolla, CA, USA) was utilized to convert structures from PDB to PDBQT format, adding polar hydrogens and assigning Kollman partial charges. The 3D structure of the linalool and β-pinene were obtained from Pubchem [55] and optimized using Gaussian 03 by quantum chemical calculations based in density functional theory (DFT) at the 6-311G level [56]. Docking studies were carried out using AutoDock Vina [57]. Docking was performed by establishing a cube of sufficient size to cover the complete protein with a grid point spacing of 1 Å. The average binding affinity for the best poses was accepted as the binding affinity value for a particular complex [56]. The existing ligand–residue interactions on the protein were evaluated with LigandScout 3.0 (Inte:Ligand, Maria Enzersdorf, Austria, Europe) [58]. The validation of the docking process was carried out by docking GABA, octopamine/tyramine and nicotinic receptors, and their inhibitors are reported in the literature (Table 2) [9,10].
4.7. Statistical Analysis
The paired t-test was utilized to compare the mean number of insects on the treated and untreated areas of the filter paper. Repellency or attraction was established if significant differences occurred for positive or negative percentage repellency, respectively. Probit analysis was employed to calculate RC50. Normal distribution and equality between variances were checked by the Kolmogorov–Smirnov and Bartlett’s tests, respectively. Two-way analysis of variance (ANOVA) with the Bonferroni post-test was used to compare all groups (treated and control) at different time intervals. The data are presented as means ± SE and the differences between means are considered to be significant at p ≤ 0.05. GraphPad InStat 3.05 (GraphPad Software, La Jolla, CA, USA) was used for data analysis. The parameters evaluated in the statistical analysis are found in Tables S1 and S2.
5. Conclusions
In summary, the monoterpenes linalool and β-pinene showed repellent activity against T. castaneum, affecting the expression of genes related to neurotransmission such as acetylcholinesterase, GABA-gated ion channel, and histamine-gated chloride channel 2. However, each chemical compound induced a different gene expression profile. These chemicals had little ability to dock on proteins associated with the neurotransmission process in the red flour beetle.
Supplementary Materials
Supplementary Materials are available online.
Acknowledgments
The authors thank the Program to Support Research Groups (2012–2014), and Grant 02295/2013, sponsored by the Vice-Presidency for Research at the University of Cartagena as well as the Grant RC-0572-2012 (Patrimonio Autónomo Fondo Nacional de Financiamiento para la Ciencia, la Tecnología y la Innovación, Francisco José de Caldas), and the National Program for Doctoral Formation (Colciencias, 567-2012).
Author Contributions
Nerlis Pajaro-Castro, Karina Caballero-Gallardo, and Jesus Olivero-Verbel conceived and designed the experiments, performed the experiments, analyzed the data, contributed reagents/materials/analysis tools, and wrote the paper.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Rattan, R.S. Mechanism of action of insecticidal secondary metabolites of plant origin. Crop Prot. 2010, 29, 913–920. [Google Scholar] [CrossRef]
- Caballero-Gallardo, K.; Olivero-Verbel, J.; Stashenko, E.E. Repellent Activity of essential oils and some of their individual constituents against Tribolium castaneum Herbst. J. Agric. Food Chem. 2011, 59, 1690–1696. [Google Scholar] [CrossRef] [PubMed]
- Blenau, W.; Rademacher, E.; Baumann, A. Plant essential oils and formamidines as insecticides/acaricides: What are the molecular targets? Apidologie 2012, 43, 334–347. [Google Scholar] [CrossRef]
- Khater, H.F. Prospects of botanical biopesticides in insect pest management. Pharmacologia 2012, 3, 641–656. [Google Scholar]
- López, M.D.; Pascual-Villalobos, M.J. Are monoterpenoids and phenylpropanoids efficient inhibitors of acetylcholinesterase from stored product insect strains? Flavour Fragr. J. 2015, 30, 108–112. [Google Scholar] [CrossRef]
- Caballero-Gallardo, K.; Olivero-Verbel, J.; Stashenko, E.E. Repellency and toxicity of essential oils from Cymbopogon martinii, Cymbopogon flexuosus and Lippia origanoides cultivated in Colombia against Tribolium castaneum. J. Stored Prod. Res. 2012, 50, 62–65. [Google Scholar] [CrossRef]
- Caballero-Gallardo, K.; Pino-Benitez, N.; Pajaro-Castro, N.; Stashenko, E.; Olivero-Verbel, J. Plants cultivated in Choco, Colombia, as source of repellents against Tribolium castaneum (Herbst). J. Asia Pac. Entomol. 2014, 17, 753–759. [Google Scholar] [CrossRef]
- Beier, R.C.; Byrd, J.A.; Kubena, L.F.; Hume, M.E.; McReynolds, J.L.; Anderson, R.C.; Nisbet, D.J. Evaluation of linalool, a natural antimicrobial and insecticidal essential oil from basil: Effects on poultry. Poult. Sci. 2014, 93, 267–272. [Google Scholar] [CrossRef] [PubMed]
- Lynagh, T.; Lynch, J. Molecular mechanisms of Cys-loop ion channel receptor modulation by ivermectin. Front. Mol. Neurosci. 2012, 5, 1–11. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Elamathi, N.; Verma, V.; Sharma, V.P.; Sreehari, U.; Raghavendra, K. Neonicotinoids in vector control: In silico approach. Asian J. Biomed. Pharm. Sci. 2014, 4, 25–29. [Google Scholar]
- Kastner, K.W.; Shoue, D.A.; Estiu, G.L.; Wolford, J.; Fuerst, M.F.; Markley, L.D.; Izaguirre, J.A.; McDowell, M.A. Characterization of the Anopheles gambiae octopamine receptor and discovery of potential agonists and antagonists using a combined computational-experimental approach. Malar. J. 2014, 13, 434. [Google Scholar] [CrossRef] [PubMed]
- Kedia, A.; Prakash, B.; Mishra, P.; Singh, P.; Dubey, N. Botanicals as ECO friendly biorational alternatives of synthetic pesticides against Callosobruchus spp. (Coleoptera: Bruchidae)—A review. J. Food Sci. Technol. 2015, 52, 1239–1257. [Google Scholar] [CrossRef] [PubMed]
- Sola, P.; Mvumi, B.M.; Ogendo, J.O.; Mponda, O.; Kamanula, J.F.; Nyirenda, S.P.; Belmain, S.R.; Stevenson, P.C. Botanical pesticide production, trade and regulatory mechanisms in sub-Saharan Africa: Making a case for plant-based pesticidal products. Food Secur. 2014, 6, 369–384. [Google Scholar] [CrossRef]
- Chowhan, N.; Singh, H.; Batish, D.; Kaur, S.; Ahuja, N.; Kohli, R. β-Pinene inhibited germination and early growth involves membrane peroxidation. Protoplasma 2013, 250, 691–700. [Google Scholar] [CrossRef] [PubMed]
- García, M.; Donadel, O.J.; Ardanaz, C.E.; Tonn, C.E.; Sosa, M.E. Toxic and repellent effects of Baccharis salicifolia essential oil on Tribolium castaneum. Pest Manag. Sci. 2005, 61, 612–618. [Google Scholar] [CrossRef] [PubMed]
- Tripathi, A.K.; Upadhyay, S.; Bhuiyan, M.; Bhattacharya, P.R. A review on prospects of essential oils as biopesticide in insect-pest management. J. Pharmacogn. Phytother. 2009, 1, 52–63. [Google Scholar]
- Müller, G.C.; Junnila, A.; Butler, J.; Kravchenko, V.D.; Revay, E.E.; Weiss, R.W.; Schlein, Y. Efficacy of the botanical repellents geraniol, linalool, and citronella against mosquitoes. J. Vector Ecol. 2009, 34, 2–8. [Google Scholar] [CrossRef] [PubMed]
- Ukeh, D.A.; Umoetok, S.B.A. Repellent effects of five monoterpenoid odours against Tribolium castaneum (Herbst) and Rhyzopertha dominica (F.) in Calabar, Nigeria. Crop Prot. 2011, 30, 1351–1355. [Google Scholar] [CrossRef]
- Re, L.; Barocci, S.; Sonnino, S.; Mencarelli, A.; Vivani, C.; Paolucci, G.; Scarpantonio, A.; Rinaldi, L.; Mosca, E. Linalool modifies the nicotinic receptor–ion channel kinetics at the mouse neuromuscular junction. Pharmacol. Res. 2000, 42, 177–181. [Google Scholar] [CrossRef] [PubMed]
- Tong, F.; Coats, J.R. Effects of monoterpenoid insecticides on [3H]-TBOB binding in house fly GABA receptor and 36Cl− uptake in American cockroach ventral nerve cord. Pest Biochem. Physiol. 2010, 98, 317–324. [Google Scholar] [CrossRef]
- Jones, A.; Sattelle, D. The cys-loop ligand-gated ion channel gene superfamily of the red flour beetle, Tribolium castaneum. BMC Genom. 2007, 8, 327. [Google Scholar] [CrossRef] [PubMed]
- Bloomquist, J.R. Chloride channels as tools for developing selective insecticides. Arch. Insect Biochem. Physiol. 2003, 54, 145–156. [Google Scholar] [CrossRef] [PubMed]
- Zheng, Y.; Hirschberg, B.; Yuan, J.; Wang, A.P.; Hunt, D.C.; Ludmerer, S.W.; Schmatz, D.M.; Cully, D.F. Identification of two novel Drosophila melanogaster histamine-gated chloride channel subunits expressed in the eye. J. Biol. Chem. 2002, 277, 2000–2005. [Google Scholar] [CrossRef] [PubMed]
- Wolstenholme, A.J. Glutamate-gated chloride channels. J. Biol. Chem. 2012, 287, 40232–40238. [Google Scholar] [CrossRef] [PubMed]
- Priestley, C.M.; Williamson, E.M.; Wafford, K.A.; Satelle, D.B. Thymol, a constituent of thyme essential oils, is a positive modulator of human GABA and a homo-oligosteric GABA receptor from Drosophila melanogaster. Br. J. Pharmacol. 2003, 140, 1363–1372. [Google Scholar] [CrossRef] [PubMed]
- Guzmán-Gutiérrez, S.L.; Bonilla-Jaime, H.; Gómez-Cansino, R.; Reyes-Chilpa, R. Linalool and β-pinene exert their antidepressant-like activity through the monoaminergic pathway. Life Sci. 2015, 128, 24–29. [Google Scholar] [CrossRef] [PubMed]
- Akiyoshi, Y.; Ju, X.-L.; Furutani, S.; Matsuda, K.; Ozoe, Y. Electrophysiological evidence for 4-isobutyl-3-isopropylbicyclophosphorothionate as a selective blocker of insect GABA-gated chloride channels. Bioorg. Med. Chem. Lett. 2013, 23, 3373–3376. [Google Scholar] [CrossRef] [PubMed]
- Lu, Y.; Park, Y.; Gao, X.; Zhang, X.; Yao, J.; Pang, Y.-P.; Jiang, H.; Zhu, K.Y. Cholinergic and non-cholinergic functions of two acetylcholinesterase genes revealed by gene-silencing in Tribolium castaneum. Sci. Rep. 2012, 2. [Google Scholar] [CrossRef] [PubMed]
- Mamidala, P.; Wijeratne, A.J.; Wijeratne, S.; Kornacker, K.; Sudhamalla, B.; Rivera-Vega, L.J.; Hoelmer, A.; Meulia, T.; Jones, S.C.; Mittapalli, O. RNA-Seq and molecular docking reveal multi-level pesticide resistance in the bed bug. BMC Genom. 2012, 13, 6. [Google Scholar] [CrossRef] [PubMed]
- Keppanan, R.; Sivaperumal, S.; Chadra Kanta, D.; Akutse, K.S.; Wang, L. Molecular docking of protease from Metarhizium anisopliae and their toxic effect against model insect Galleria mellonella. Pestic. Biochem. Physiol. 2017, 138, 8–14. [Google Scholar] [CrossRef] [PubMed]
- Aloui, S.; Raboudi, F.; Ghazouani, T.; Salghi, R.; Hamdaoui, M.H.; Fattouch, S. Use of molecular and in silico bioinformatic tools to investigate pesticide binding to insect (Lepidoptera) phenoloxidases (PO): Insights to toxicological aspects. J. Environ. Sci. Health Part B 2014, 49, 654–660. [Google Scholar] [CrossRef] [PubMed]
- Wallner, B.; Elofsson, A. Can correct protein models be identified? Protein Sci. 2003, 12, 1073–1086. [Google Scholar] [CrossRef] [PubMed]
- Sarkar, S.; Gupta, S.; Chakraborty, W.; Senapati, S.; Gachhui, R. Homology modeling, molecular docking and molecular dynamics studies of the catalytic domain of chitin deacetylase from Cryptococcus laurentii strain RY1. Int. J. Biol. Macromol. 2017, 104, 1682–1691. [Google Scholar] [CrossRef] [PubMed]
- Akhter, M.; Tasleem, M.; Mumtaz Alam, M.; Ali, S. In silico approach for bioremediation of arsenic by structure prediction and docking studies of arsenite oxidase from Pseudomonas stutzeri TS44. Int. Biodeterior. Biodegrad. 2017, 122, 82–91. [Google Scholar] [CrossRef]
- Aloy, P.; Pichaud, M.; Russell, R.B. Protein complexes: Structure prediction challenges for the 21st century. Curr. Opin. Struct. Biol. 2005, 15, 15–22. [Google Scholar] [CrossRef] [PubMed]
- Chivian, D.; Kim, D.E.; Malmstrom, L.; Schonbrun, J.; Rohl, C.A.; Baker, D. Prediction of CASP6 structures using automated Robetta protocols. Proteins 2005, 61, 157–166. [Google Scholar] [CrossRef] [PubMed]
- Alejandro, S.-P.; Maria Natalia Dias Soeiro, C.; Lisvey, G.-M.; Reider, Y.-B. Current Computational approaches towards the rational design of new insecticidal agents. Curr. Comput. Aided Drug Des. 2011, 7, 304–314. [Google Scholar]
- Cornejo, I.; Andrini, O.; Niemeyer, M.I.; Marabolí, V.; González-Nilo, F.D.; Teulon, J.; Sepúlveda, F.V.; Cid, L.P. Identification and functional expression of a glutamate- and avermectin-gated chloride channel from Caligus rogercresseyi, a Southern Hemisphere sea louse affecting farmed fish. PLoS Pathog. 2014, 10, e1004402. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dönitz, J.; Grossmann, D.; Schild, I.; Schmitt-Engel, C.; Bradler, S.; Prpic, N.; Bucher, G. TrOn: An Anatomical Ontology for the Beetle Tribolium castaneum. PLoS ONE 2013, 8, e70695. [Google Scholar] [CrossRef] [PubMed]
- Lord, J.C.; Hartzer, K.; Toutges, M.; Oppert, B. Evaluation of quantitative PCR reference genes for gene expression studies in Tribolium castaneum after fungal challenge. J. Microbiol. Methods 2010, 80, 219–221. [Google Scholar] [CrossRef] [PubMed]
- Vuerinckx, K.; Verlinden, H.; Lindemans, M.; Broeck, J.V.; Huybrechts, R. Characterization of an allatotropin-like peptide receptor in the red flour beetle, Tribolium castaneum. Insect Biochem. Mol. Biol. 2011, 41, 815–822. [Google Scholar] [CrossRef] [PubMed]
- Jentsch, T.J.; Neagoe, I.; Scheel, O. CLC chloride channels and transporters. Curr. Opin. Neurobiol. 2005, 15, 319–325. [Google Scholar] [CrossRef] [PubMed]
- Orhan, G.; Fahlke, C.; Alekov, A.K. Anion- and Proton-Dependent Gating of ClC-4 Anion/Proton Transporter under Uncoupling Conditions. Biophys. J. 2011, 100, 1233–1241. [Google Scholar] [CrossRef] [PubMed]
- Lu, Y.; Pang, Y.-P.; Park, Y.; Gao, X.; Yao, J.; Zhang, X.; Zhu, K.Y. Genome organization, phylogenies, expression patterns, and three-dimensional protein models of two acetylcholinesterase genes from the red flour beetle. PLoS ONE 2012, 7, e32288. [Google Scholar] [CrossRef] [PubMed]
- Arya, M.; Shergill, I.S.; Williamson, M.; Gommersall, L.; Arya, N.; Patel, H.R. Basic principles of real-time quantitative PCR. Expert Rev. Mol. Diagn. 2005, 5, 209–219. [Google Scholar] [CrossRef] [PubMed]
- Valasek, M.A.; Repa, J.J. The power of real-time PCR. Adv. Physiol. Educ. 2005, 29, 151–159. [Google Scholar] [CrossRef] [PubMed]
- Wang, F.; Wang, J.; Liu, D.; Su, Y. Normalizing genes for real-time polymerase chain reaction in epithelial and nonepithelial cells of mouse small intestine. Anal. Biochem. 2010, 399, 211–217. [Google Scholar] [CrossRef] [PubMed]
- UniProt, C. Reorganizing the protein space at the Universal Protein Resource (UniProt). Nucleic Acids Res. 2012, 40, D71–D75. [Google Scholar]
- Chen, C.-C.; Hwang, J.-K.; Yang, J.-M. (PS)2-v2: Template-based protein structure prediction server. BMC Bioinform. 2009, 10, 366. [Google Scholar] [CrossRef] [PubMed]
- Chen, C.-C.; Hwang, J.-K.; Yang, J.-M. (PS)2: Protein structure prediction server. Nucleic Acids Res. 2006, 34, W152–W157. [Google Scholar] [CrossRef] [PubMed]
- Kelley, L.A.; Mezulis, S.; Yates, C.M.; Wass, M.N.; Sternberg, M.J.E. The Phyre2 web portal for protein modeling, prediction and analysis. Nat. Protoc. 2015, 10, 845–858. [Google Scholar] [CrossRef] [PubMed]
- Roy, A.; Kucukural, A.; Zhang, Y. I-TASSER: A unified platform for automated protein structure and function prediction. Nat. Protoc. 2010, 5, 725–738. [Google Scholar] [CrossRef] [PubMed]
- Lovell, S.C.; Davis, I.W.; Arendall, W.B.; de Bakker, P.I.W.; Word, J.M.; Prisant, M.G.; Richardson, J.S.; Richardson, D.C. Structure validation by Cα geometry: ϕ, ψ, and Cβ deviation. Proteins 2003, 50, 437–450. [Google Scholar] [CrossRef] [PubMed]
- Benkert, P.; Tosatto, S.C.E.; Schomburg, D. QMEAN: A comprehensive scoring function for model quality assessment. Proteins 2008, 71, 261–277. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.; Thiessen, P.A.; Bolton, E.E.; Chen, J.; Fu, G.; Gindulyte, A.; Han, L.Y.; He, J.E.; He, S.Q.; Shoemaker, B.A.; et al. PubChem Substance and Compound databases. Nucleic Acids Res. 2016, 44, D1202–D1213. [Google Scholar] [CrossRef] [PubMed]
- Pájaro-Castro, N.; Flechas, M.; Ocazionez, R.; Staschenko, E.; Olivero-Verbel, J. Potential interaction of components from essential oils with dengue virus proteins. Boletín Latinoamericano y del Caribe de Plantas Medicinales y Aromáticas 2015, 14, 141–155. [Google Scholar]
- Trott, O.; Olson, A.J. AutoDock Vina: Improving the speed and accuracy of docking with a new scoring function, efficient optimization and multithreading. J. Comput. Chem. 2010, 31, 455–461. [Google Scholar] [CrossRef] [PubMed]
- Wolber, G.; Langer, T. LigandScout: 3-D Pharmacophores derived from protein-bound ligands and their use as virtual screening filters. J. Chem. Inf. Model. 2005, 45, 160–169. [Google Scholar] [CrossRef] [PubMed]
Sample Availability: Samples of the compounds linalool and β-pinene are available from the authors. |
Figure 1.
2D chemical structures of linalool (A) and β-pinene (B).
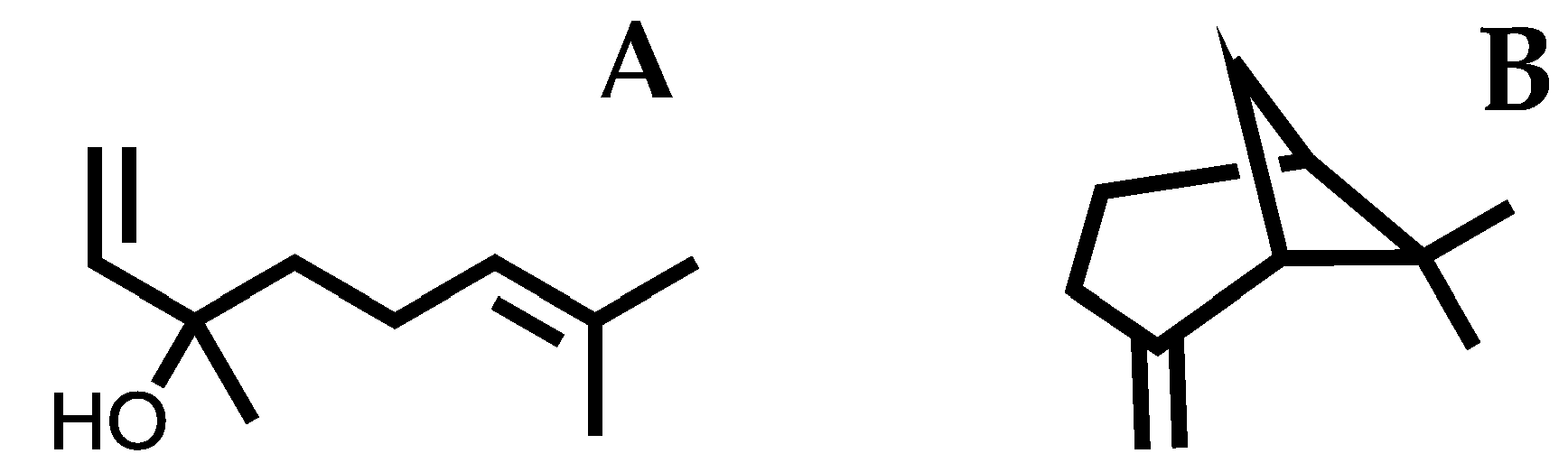
Figure 2.
Percentage repellence of linalool (A) and β-pinene (B) against T. castaneum. * Statistically significant compared to control (p < 0.05). (A) t-value = 1.050; F-value = 4.411; (B) t-value = 2.646; F-value = 4.133.
Figure 2.
Percentage repellence of linalool (A) and β-pinene (B) against T. castaneum. * Statistically significant compared to control (p < 0.05). (A) t-value = 1.050; F-value = 4.411; (B) t-value = 2.646; F-value = 4.133.
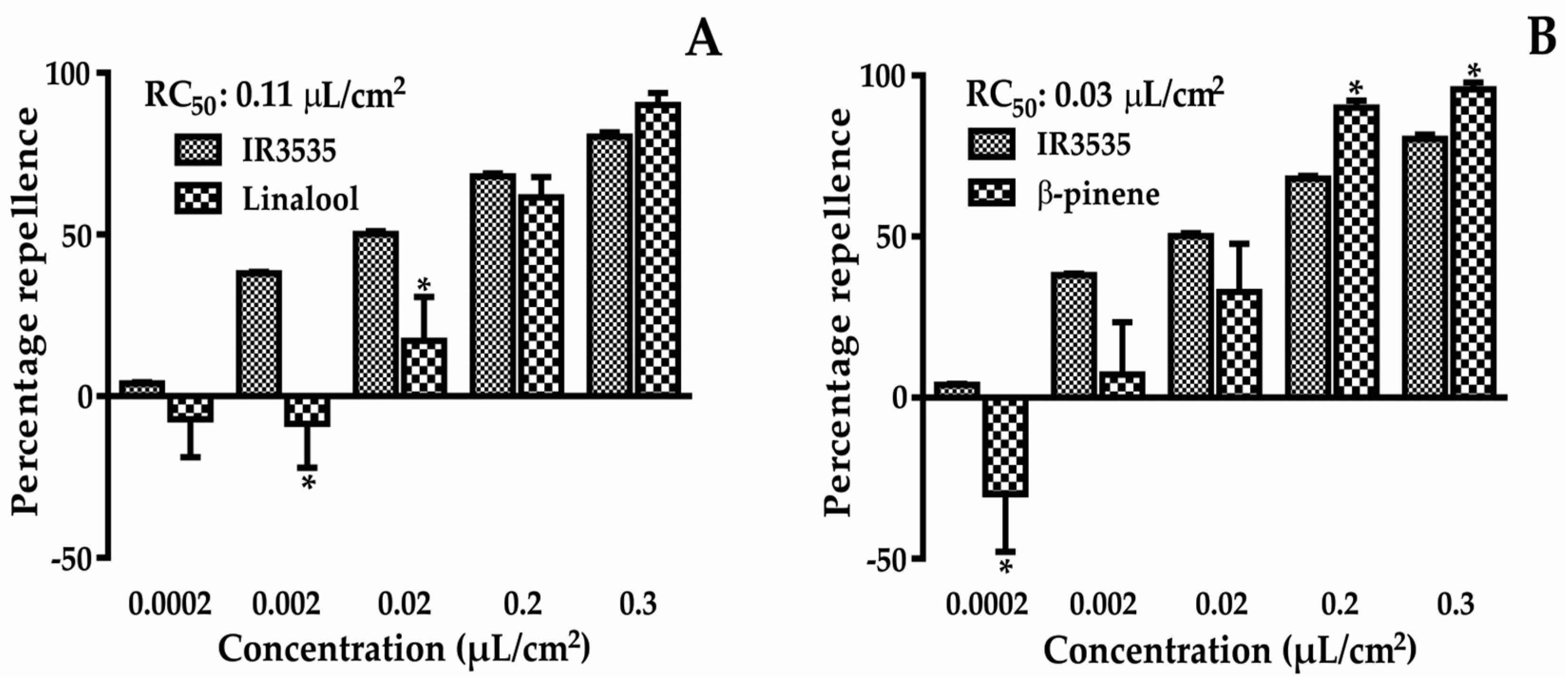
Figure 3.
Relative mRNA expression of selected genes in adults of T. castaneum after 4 h exposure to different concentrations of linalool and β-pinene. Within each gene, relative expression was normalized against the control (Value = 1). * Significant effects compared to control. Rdl: t-value = 1.636; F-value = 0.542; Hr39: t-value = 1.718; F-value = 0.903; GluCl: t-value = 0.450; F-value = 0.579; Grd: t-value = 0.042; F-value = 0.065; Ace1: t-value = 2.559; F-value = 0.924; HisCl2: t-value = 3.157; F-value = 0.715.
Figure 3.
Relative mRNA expression of selected genes in adults of T. castaneum after 4 h exposure to different concentrations of linalool and β-pinene. Within each gene, relative expression was normalized against the control (Value = 1). * Significant effects compared to control. Rdl: t-value = 1.636; F-value = 0.542; Hr39: t-value = 1.718; F-value = 0.903; GluCl: t-value = 0.450; F-value = 0.579; Grd: t-value = 0.042; F-value = 0.065; Ace1: t-value = 2.559; F-value = 0.924; HisCl2: t-value = 3.157; F-value = 0.715.

{kind=link}
{kind=link}
{kind=link}
Table 1.
Results of the validation of homology-modeled proteins from T. castaneum using ProQ.
| Proteins | LGscore | MaxSub |
|---|---|---|
| Carboxylic ester hydrolase | 5.448 | 0.154 |
| Carboxylic ester hydrolase 2 (acetylcholinesterase activity) | 6.207 | 0.459 |
| Gamma-aminobutyric acid-gated anion channel splice variant 3a6a (GABA-RDL) | 1.520 | 0.087 |
| Gamma-aminobutyric acid-gated ion channel (GABA-GRD) | 2.658 | 0.120 |
| Gamma-aminobutyric acid-ligand gated chloride channel 3) (GABA-LCCH3) | 2.611 | 0.129 |
| Glutamate-gated chloride channel | 2.010 | 0.150 |
| Histamine-gated chloride channel 1 | 2.487 | 0.166 |
| Histamine-gated chloride channel 2 | 2.435 | 0.143 |
| Hormone receptor in 39-like protein | 0.947 | 0.432 |
| Nicotinic acetylcholine receptor subunit alpha1 | 1.580 | 0.090 |
| Nicotinic acetylcholine receptor subunit alpha2 | 1.707 | 0.118 |
| pH sensitive chloride channel | 1.740 | 0.139 |
| Putative octopamine/tyramine receptor | 2.150 | 0.100 |
Table 2.
AutoDock vina-calculated affinities (Kcal/mol) obtained for docking linalool and β-pinene on some proteins.
Table 2.
AutoDock vina-calculated affinities (Kcal/mol) obtained for docking linalool and β-pinene on some proteins.
| Proteins | Uniprot Code | Linalool | β-Pinene | Inhibitors |
|---|---|---|---|---|
| Carboxylic ester hydrolase | E7DN61 | −5.3 ± 0.1 | −5.8 ± 0.0 | |
| Carboxylic ester hydrolase 2 (acetylcholinesterase activity) | E7DN62 | −5.5 ± 0.2 | −6.1 ± 0.1 | |
| Gamma-aminobutyric acid-gated anion channel splice variant 3a6a | A8DMU1 | −4.9 ± 0.1 | −6.1 ± 0.0 | −9.8 ± 0.0 * |
| Gamma-aminobutyric acid-gated ion channel | A8DMU2 | −5.1 ± 0.1 | −6.0 ± 0.0 | −9.4 ± 0.0 * |
| Gamma-aminobutyric acid-ligand gated chloride channel 3 | A8DMU3 | −6.7 ± 0.2 | −7.2 ± 0.0 | −9.0 ± 0.0 * |
| Glutamate-gated chloride channel | A8DMU5 | −4.9 ± 0.2 | −6.6 ± 0.3 | |
| Histamine-gated chloride channel 1 | A8DMU7 | −5.3 ± 0.2 | −5.6 ± 0.0 | |
| Histamine-gated chloride channel 2 | A8DMU8 | −5.0 ± 0.2 | −5.7 ± 0.0 | |
| Hormone receptor in 39-like protein | D2A6H1 | −5.2 ± 0.2 | −6.0 ± 0.0 | |
| Nicotinic acetylcholine receptor subunit alpha1 | A8DIP3 | −7.1 ± 0.2 | −7.4 ± 0.1 | −8.8 ± 0.0 + |
| Nicotinic acetylcholine receptor subunit alpha2 | A8DIQ7 | −6.8 ± 0.3 | −7.6 ± 0.0 | −8.5 ± 0.0 + |
| pH sensitive chloride channel | A8DMU9 | −5.4 ± 0.2 | −5.0 ± 0.0 | |
| Putative octopamine/tyramine receptor | D6WB14 | −7.2 ± 0.2 | −7.7±0.0 | −8.2 ± 0.0 ‡ |
Table 3.
Primer sequences of the genes used for real-rime polymerase chain reaction (RT-PCR).
| Gene Name | Gene Symbol | Entrez Gene ID | Forward (5′-3′) | Reverse (5′-3′) | Amplicon Size |
|---|---|---|---|---|---|
| Genes Evaluated | |||||
| Acetylcholinesterase | Ace1 | HQ260968.1 | CCGTTCGTCCCAGTCATTG | AGTAGTAGCCTTCTTCTGTGTTAG | 121 |
| GABA-gated anion channel splice variant 3a6a | Rdl | NM_001114292.1 | ACTTGGGCGACGTCAACATA | ACGTGAAATCCATCTGGACC | 159 |
| GABA-gated ion channel | Grd | NM_001114300.1 | GGTCTCCTTCTGGCTGAACC | TGGACCACAGCGAACTGAAT | 198 |
| Glutamate-gated chloride channel | Glucl | NM_001114304.1 | TGAATGGCACAGATGGTCCC | CCAGACTCGACTGGCTTCAG | 194 |
| Histamine-gated chloride channel 2 | Hiscl2 | NM_001109951.1 | TGGATGTCCAGTTGTTCGGT | TGTGGCTGAATAGGCAAGTCAT | 176 |
| Hormone receptor in 39-like protein | Hr39 | XR_043083.1 | CGACCGTCGACTGTACAAAA | AGTCGACATGGAACGGAAAC | 145 |
| Housekeeping Gene | |||||
| Ribosomal protein 49 | Rps49 | XM_964471.2 | TGGCAAACTCAAACGCAACT | AGCGCCTACGAACCCTGTT | 62 |
| Ribosomal protein 18 | Rps18 | XM_968539.2 | CGAAGAGGTCGAGAAAATCG | CGTGGTCTTGGTGTGTTGAC | 235 |
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Pajaro-Castro, N.; Caballero-Gallardo, K.; Olivero-Verbel, J. Neurotoxic Effects of Linalool and β-Pinene on Tribolium castaneum Herbst. Molecules 2017, 22, 2052. https://doi.org/10.3390/molecules22122052
AMA Style
Pajaro-Castro N, Caballero-Gallardo K, Olivero-Verbel J. Neurotoxic Effects of Linalool and β-Pinene on Tribolium castaneum Herbst. Molecules. 2017; 22(12):2052. https://doi.org/10.3390/molecules22122052
Chicago/Turabian StylePajaro-Castro, Nerlis, Karina Caballero-Gallardo, and Jesus Olivero-Verbel. 2017. "Neurotoxic Effects of Linalool and β-Pinene on Tribolium castaneum Herbst" Molecules 22, no. 12: 2052. https://doi.org/10.3390/molecules22122052