Antibacterial Activity of the Non-Cytotoxic Peptide (p-BthTX-I)2 and Its Serum Degradation Product against Multidrug-Resistant Bacteria

 , , and
, , and
Abstract
:1. Introduction
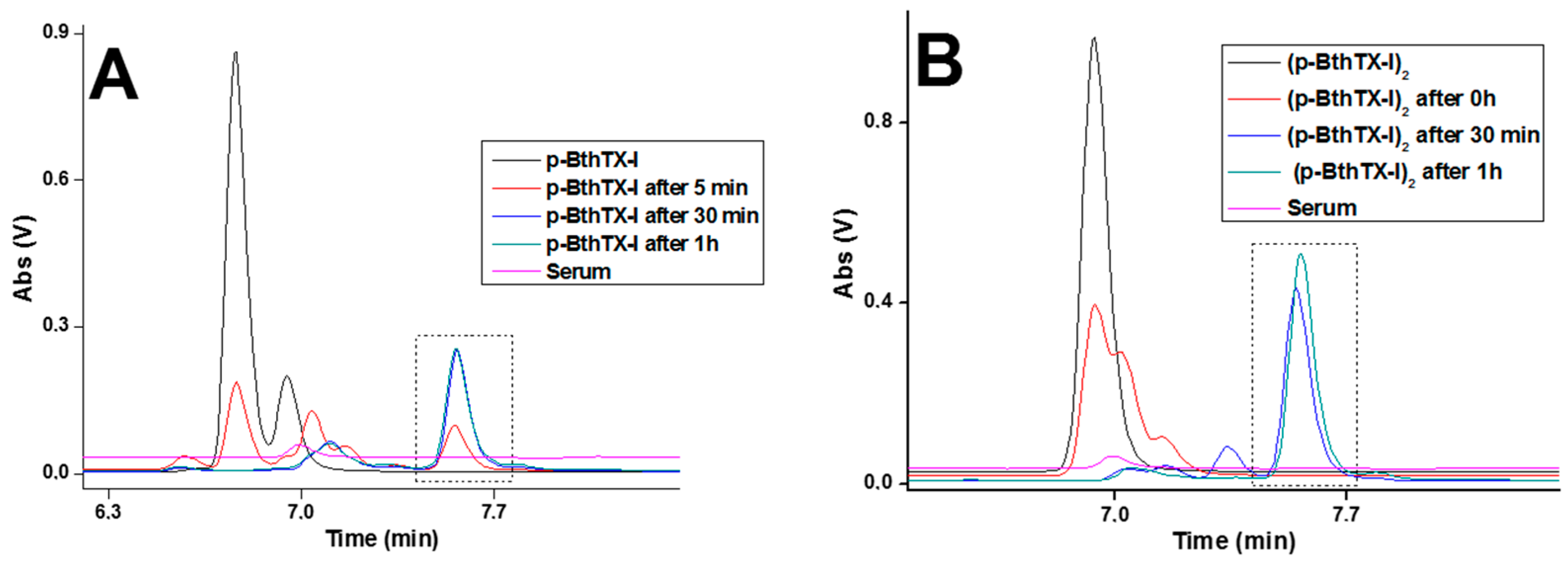
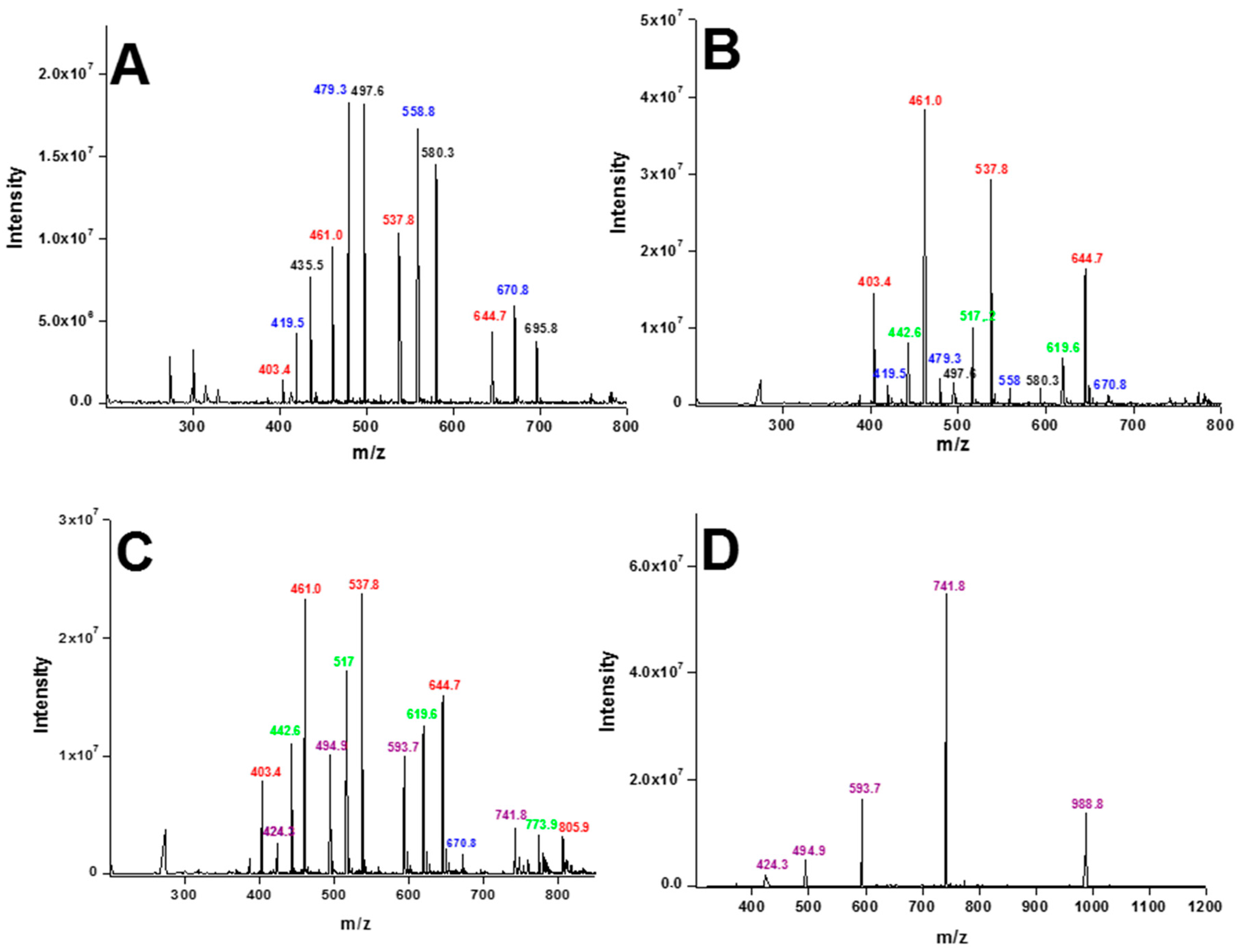
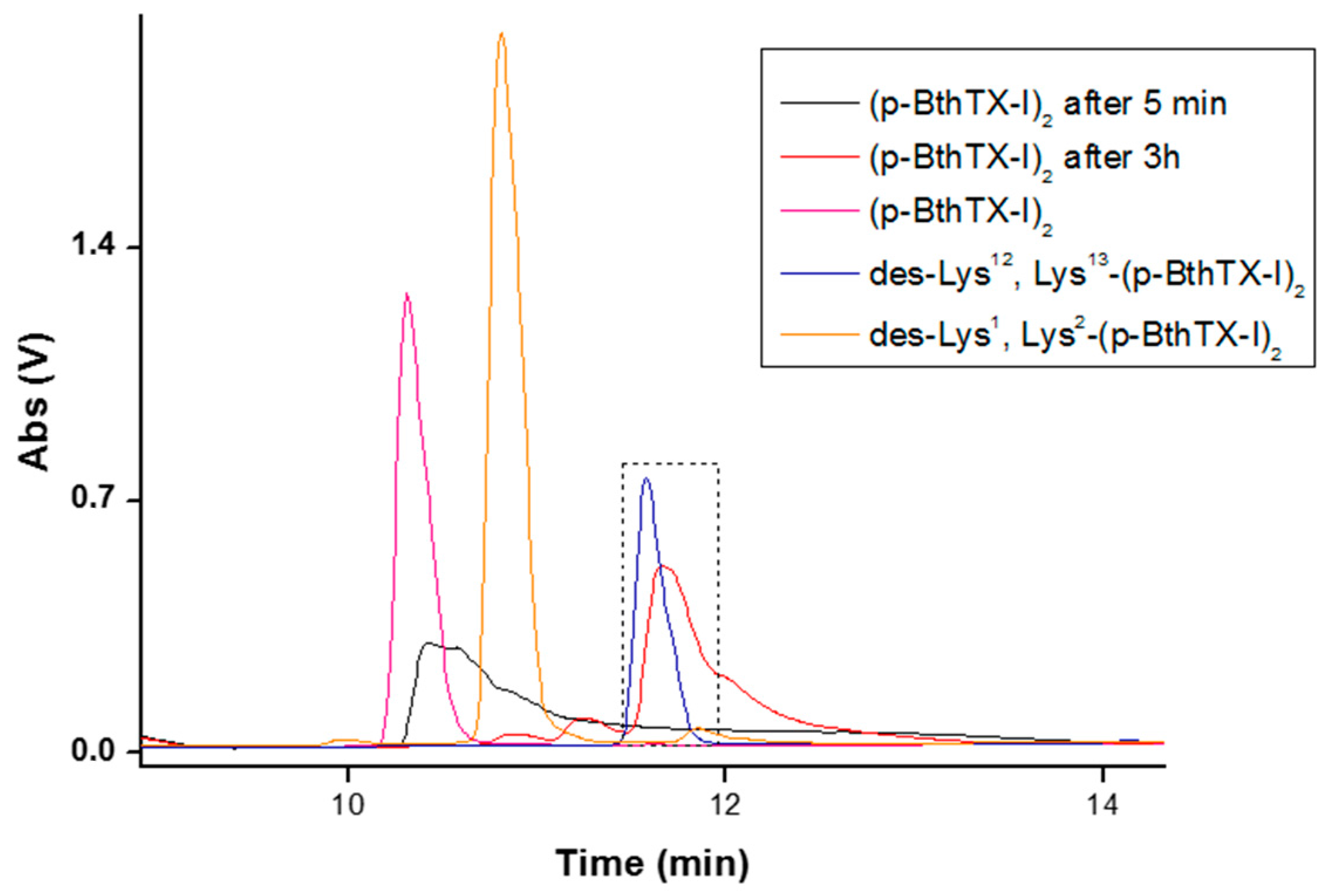
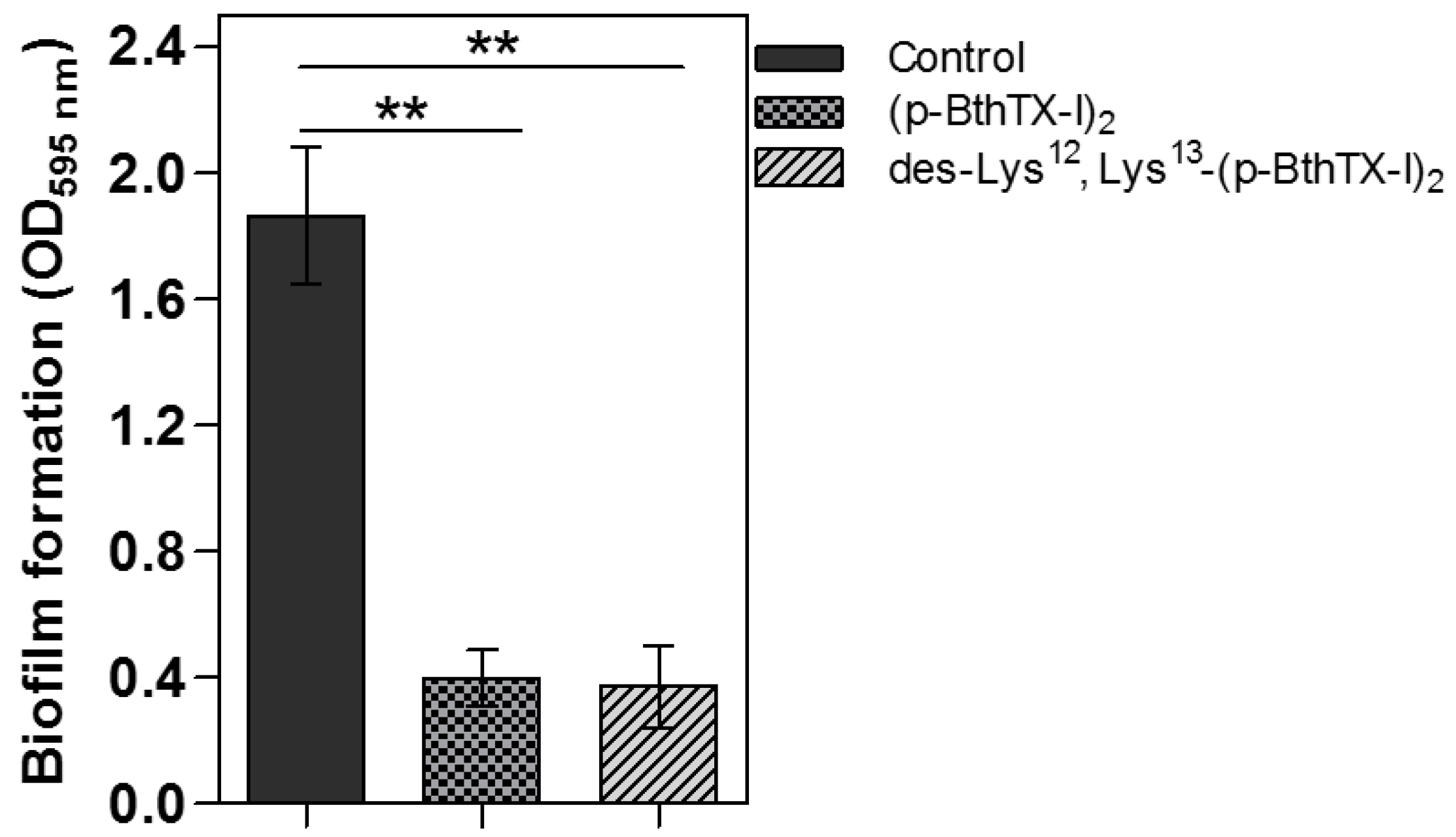
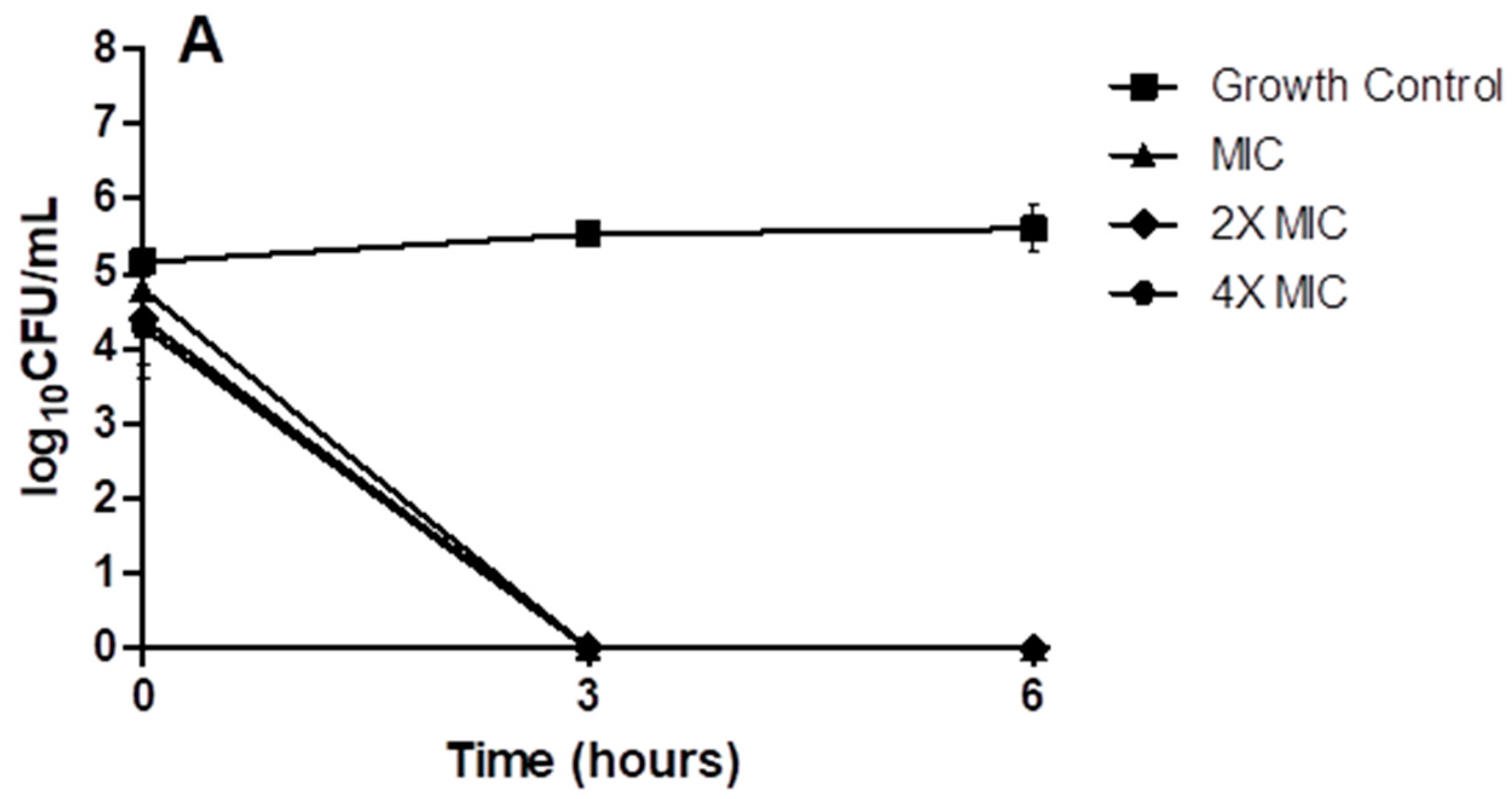
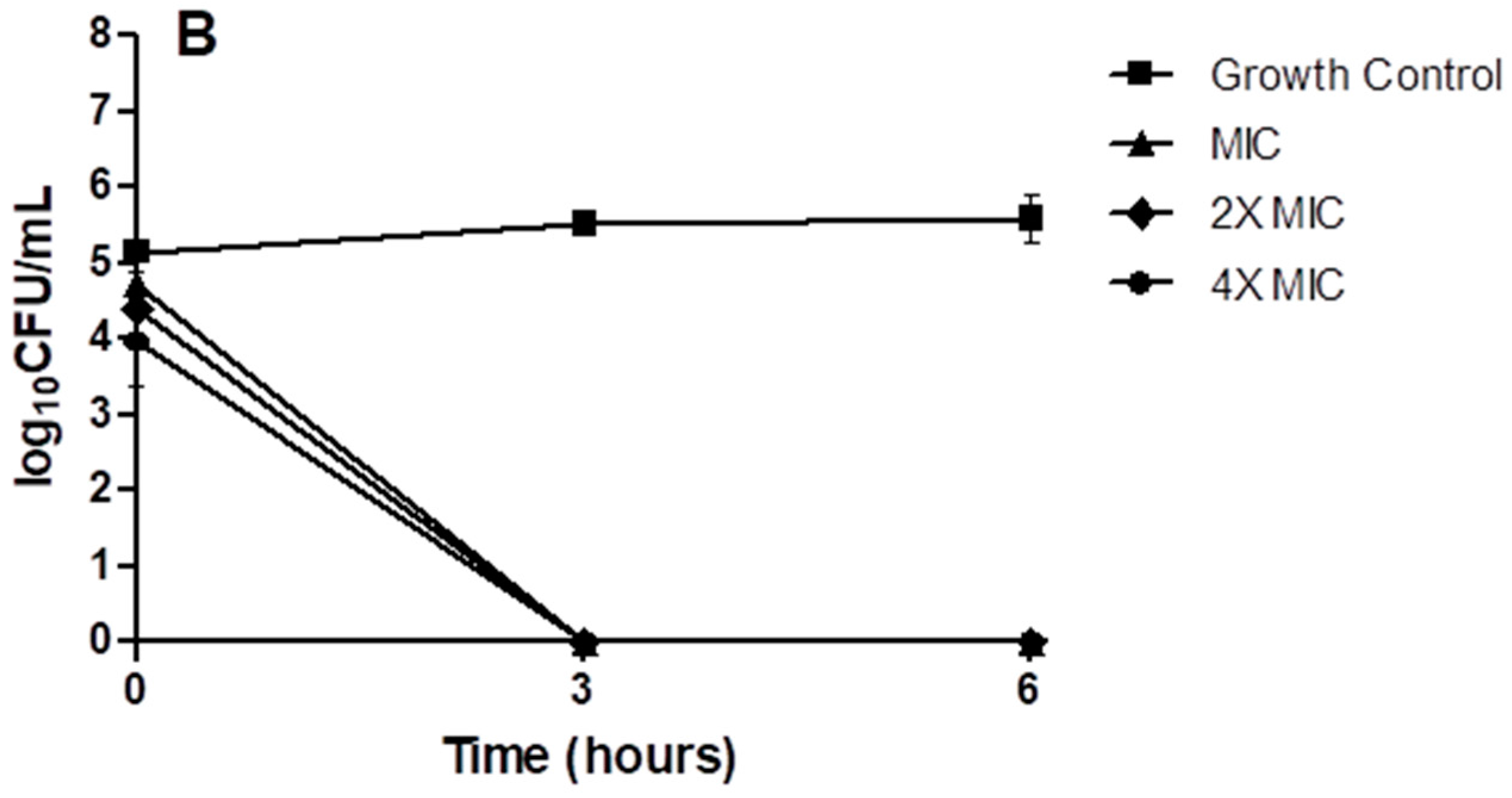
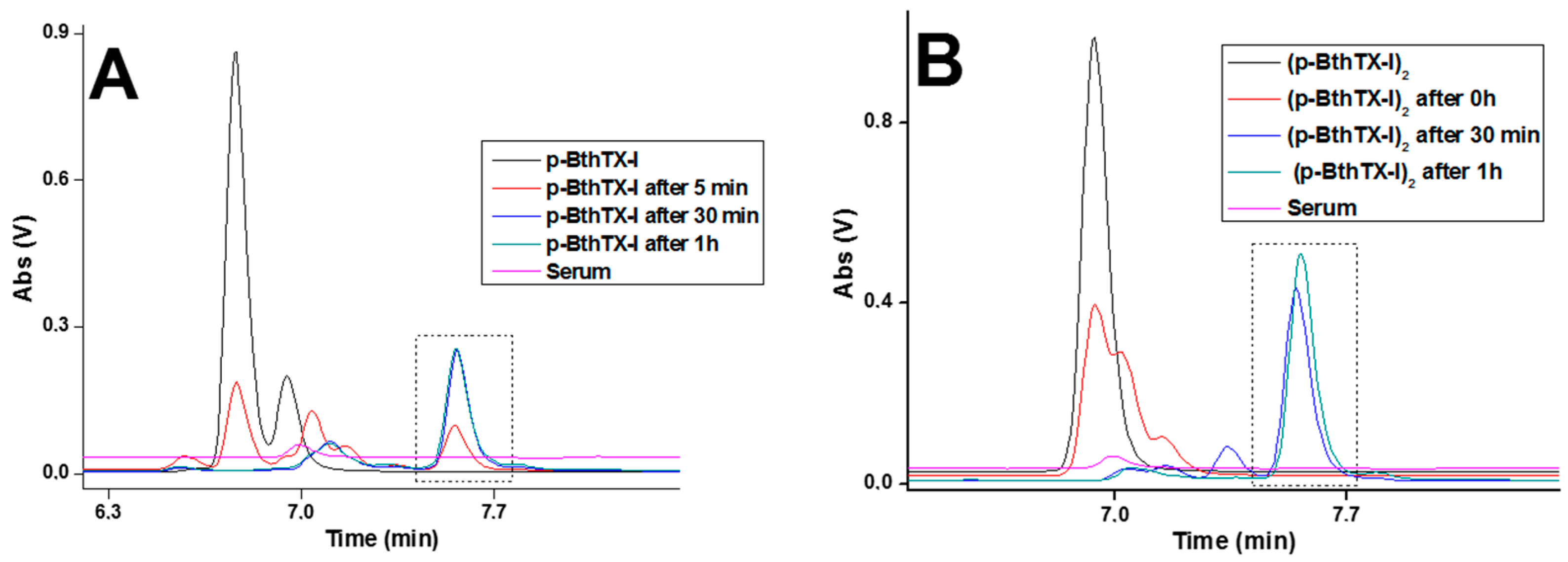
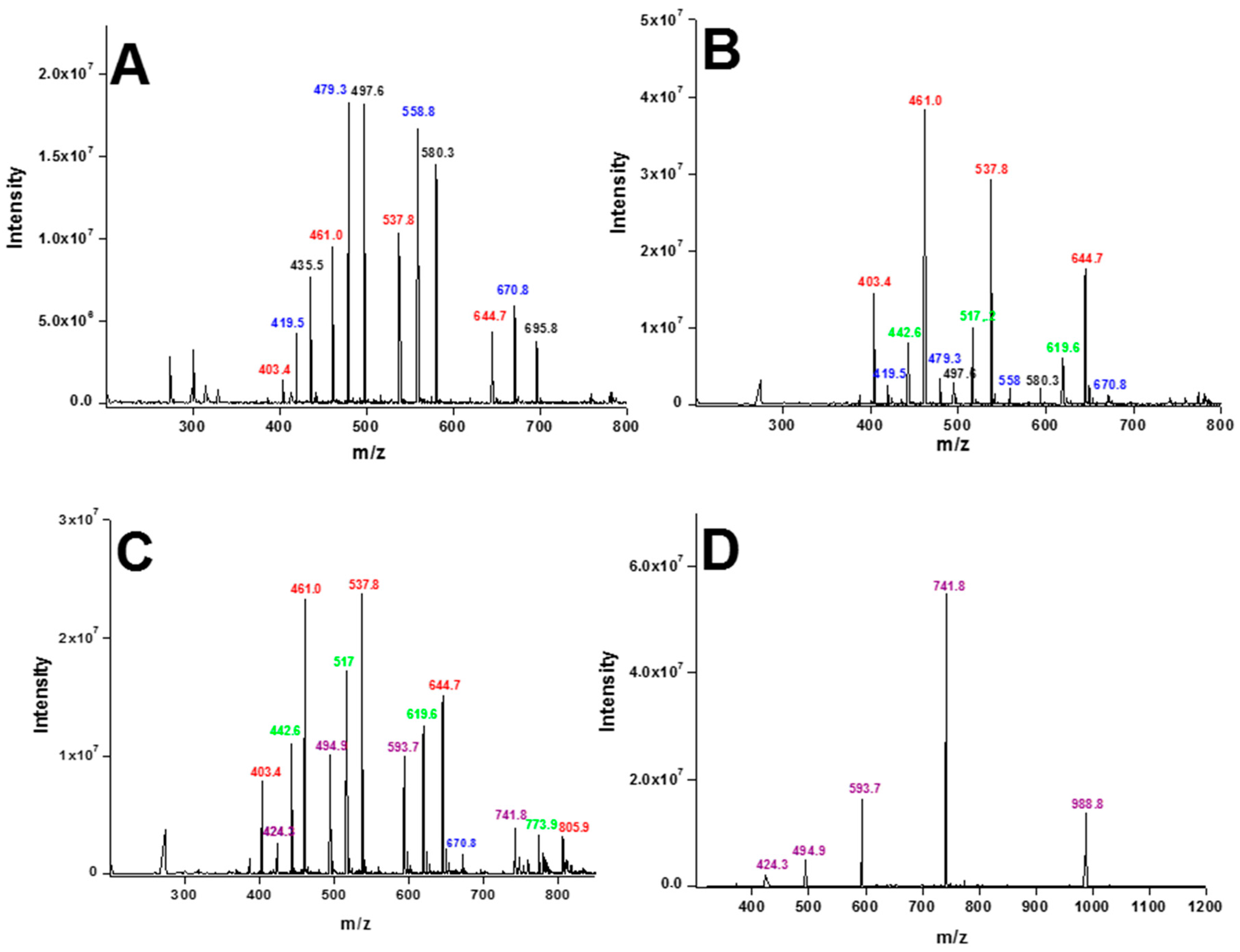
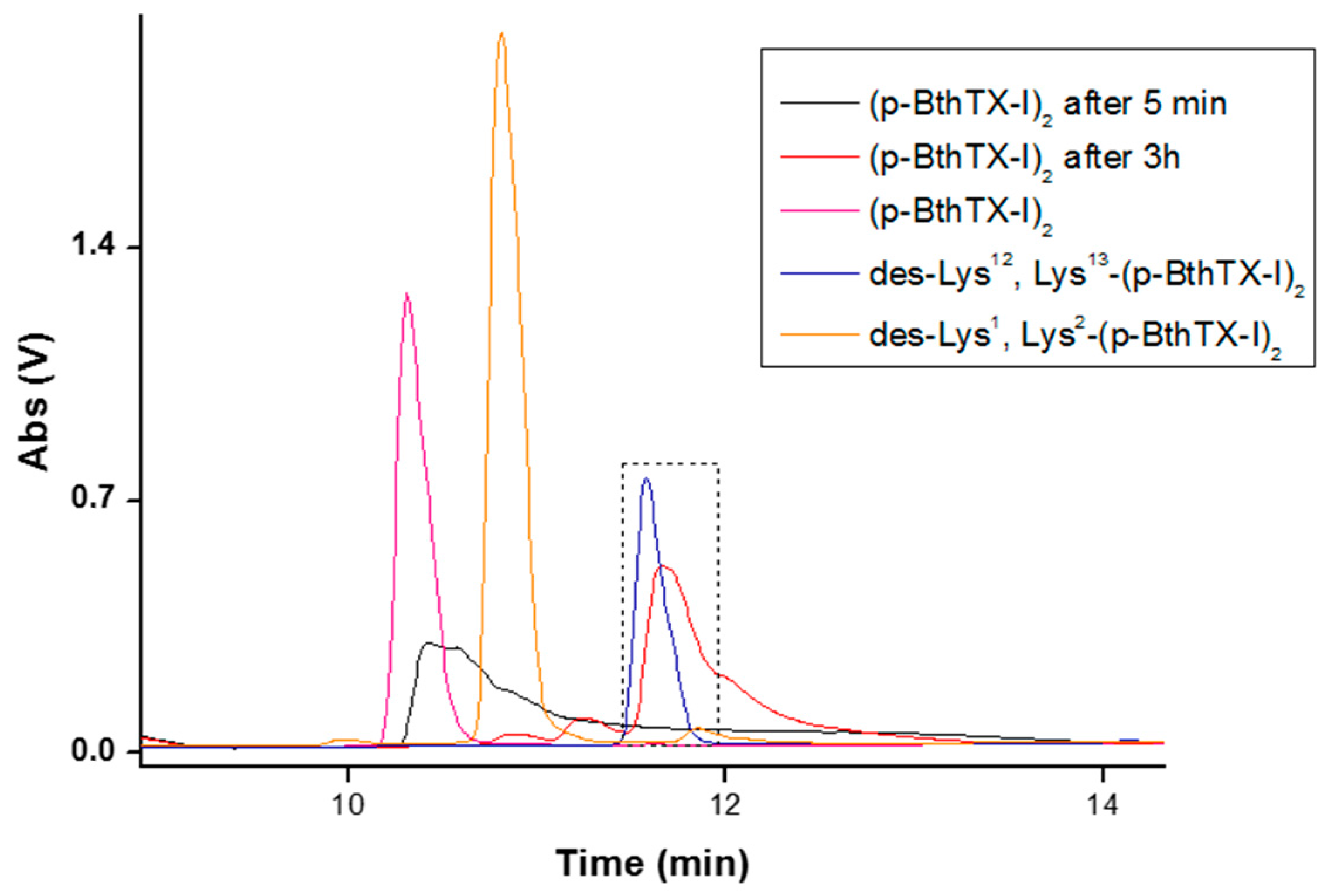
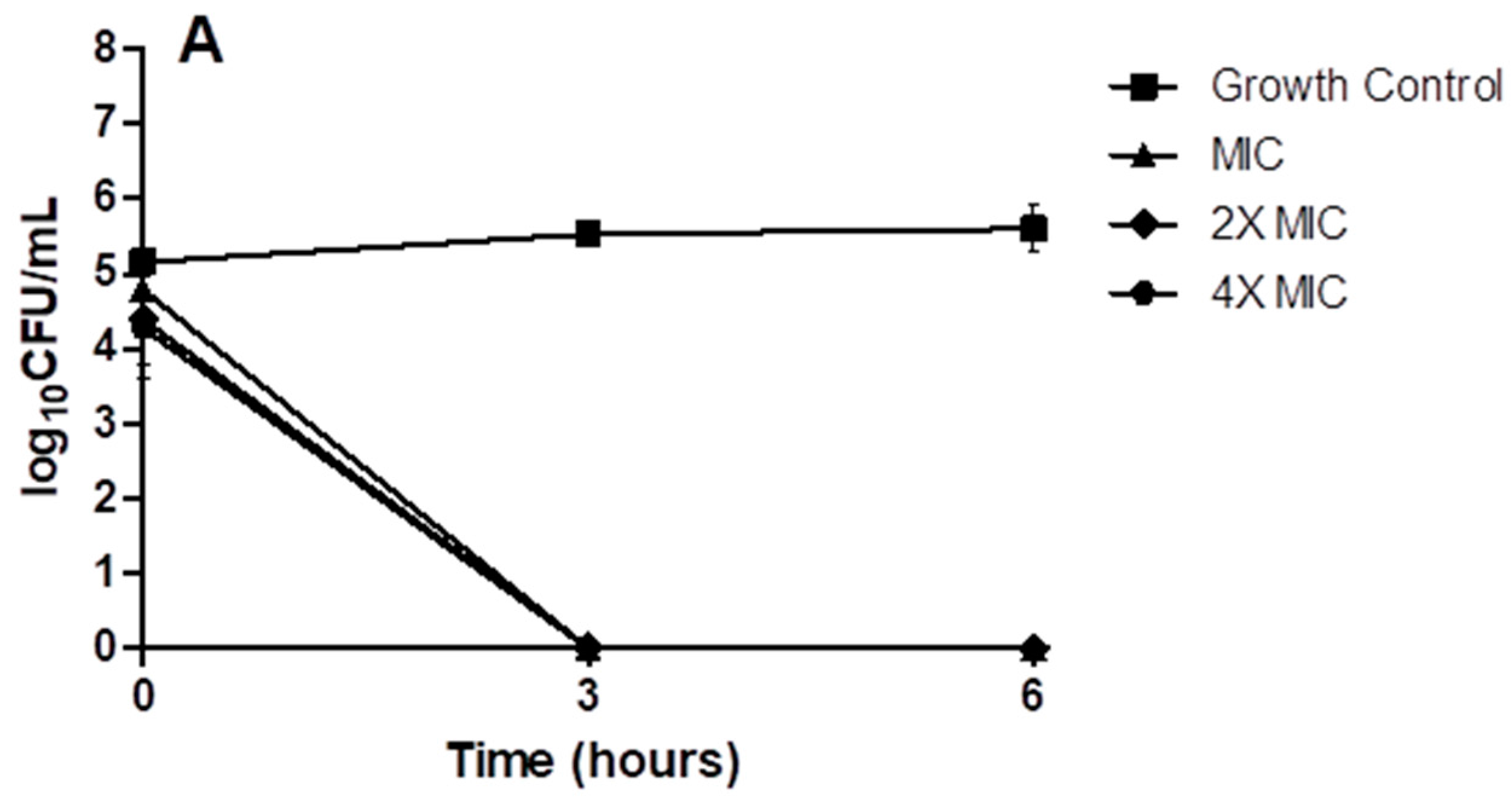
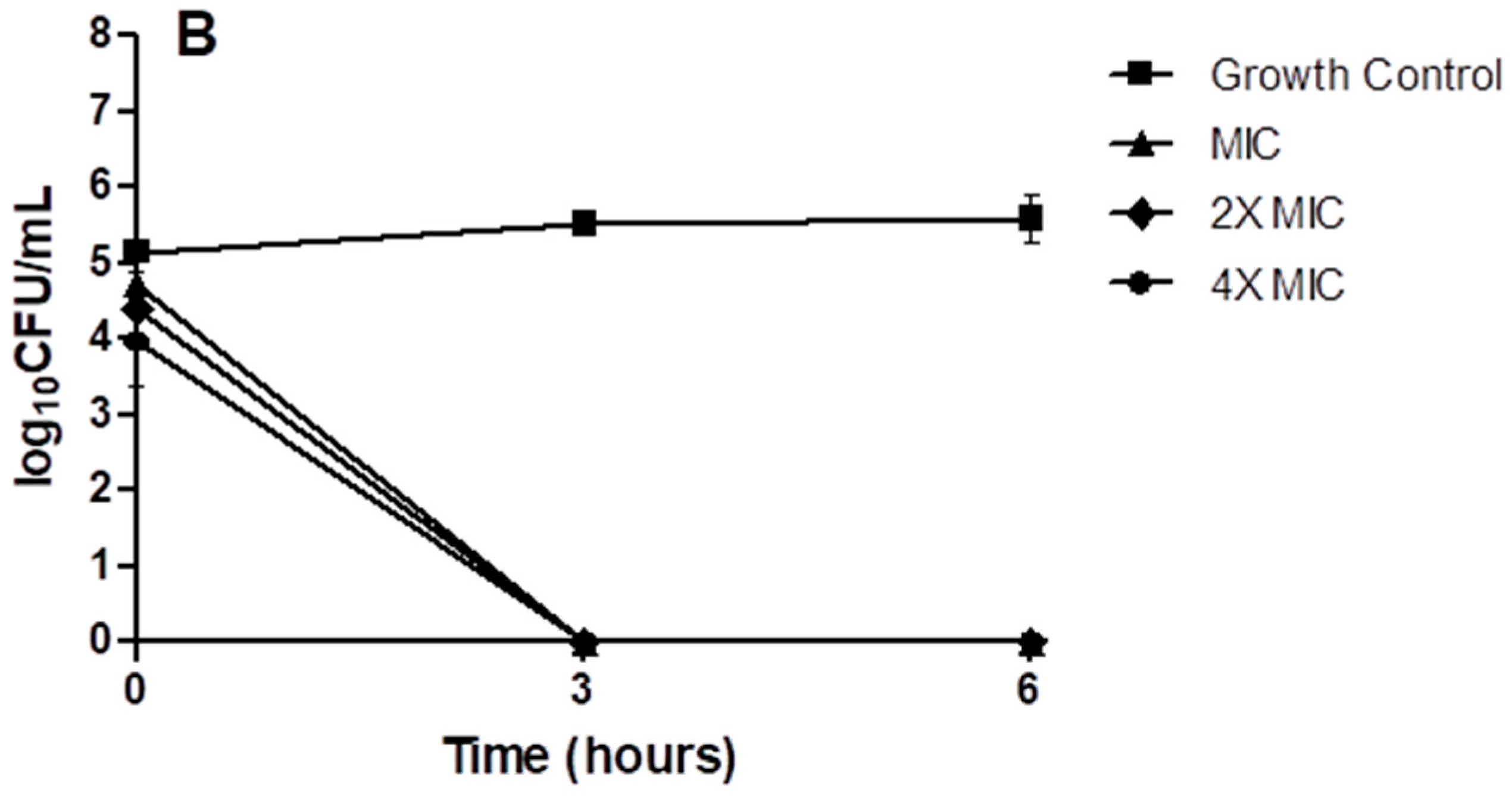
2. Results
3. Discussion
4. Materials and Methods
4.1. Peptide Synthesis
4.2. Electrospray Mass Spectrometry
4.3. Reverse Phase Chromatography
4.4. Serum Stability Assay
4.5. Circular Dichroism Spectroscopy
4.6. Hemolysis Assay
4.7. Fluorescence Spectroscopy
4.8. In Vitro Evaluation of the Antimicrobial Activity
4.9. Biofilm Eradication
4.10. Time-Kill Kinetics
5. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Bush, K. Antibacterial drug discovery in the 21st century. Clin. Microbiol. Infect. 2004, 10 (Suppl. S4), 10–17. [Google Scholar] [CrossRef] [PubMed]
- Lomonte, B.; Angulo, Y.; Moreno, E. Synthetic peptides derived from the C-terminal region of Lys49 phospholipase A2 homologues from Viperidae snake venoms: Biomimetic activities and potential applications. Curr. Pharm. Des. 2010, 16, 3224–3230. [Google Scholar] [CrossRef] [PubMed]
- Gauri, S.S.; Bera, C.K.; Bhattacharyyam, R.; Mandal, S.M. Identification of an antimicrobial peptide from large freshwater snail (Lymnaea stagnalis): Activity against antibiotics resistant Staphylococcus epidermidis. Int. J. Exp. Res. Rev. 2016, 2, 5–9. [Google Scholar]
- Berdy, J. Thoughts and facts about antibiotics: Where we are now and where we are heading. J. Antibiot. 2012, 65, 385–395. [Google Scholar] [CrossRef] [PubMed]
- Da Silva, B.R.; de Freitas, V.A.; Carneiro, V.A.; Arruda, F.V.; Lorenzón, E.N.; de Aguiar, A.S.; Cilli, E.M.; Cavada, B.S.; Teixeira, E.H. Antimicrobial activity of the synthetic peptide Lys-a1 against oral streptococci. Peptides 2013, 42, 78–83. [Google Scholar] [CrossRef] [PubMed]
- Kreling, P.F.; Aida, K.L.; Massunari, L.; Caiaffa, K.S.; Percinoto, C.; Bedran, T.B.; Spolidorio, D.M.; Abuna, G.F.; Cilli, E.M.; Duque, C. Cytotoxicity and the effect of cationic peptide fragments against cariogenic bacteria under planktonic and biofilm conditions. Biofouling 2016, 32, 995–1006. [Google Scholar] [CrossRef] [PubMed]
- Lorenzón, E.N.; Santos-Filho, N.A.; Ramos, M.A.; Bauab, T.M.; Camargo, I.L.; Cilli, E.M. C-Terminal lysine-linked magainin 2 with increased activity against multidrug-resistant bacteria. Protein Pept. Lett. 2016, 23, 738–747. [Google Scholar] [CrossRef] [PubMed]
- Cespedes, G.F.; Lorenzón, E.N.; Vicente, E.F.; Mendes-Giannini, M.J.; Fontes, W.; Castro, M.S.; Cilli, E.M. Mechanism of action and relationship between structure and biological activity of Ctx-Ha: A new ceratotoxin-like peptide from Iypsiboas albopunctatus. Protein Pept. Lett. 2012, 19, 596–603. [Google Scholar] [CrossRef] [PubMed]
- Murillo, L.A.; Lan, C.Y.; Agabian, N.M.; Larios, S.; Lomonte, B. Fungicidal activity of a phospholipase A2-derived synthetic peptide variant against Candida albicans. Rev. Esp. Quimioter. 2007, 20, 330–333. [Google Scholar] [PubMed]
- Wang, G.; Li, X.; Wang, Z. Apd3: The antimicrobial peptide database as a tool for research and education. Nucleic Acids Res. 2016, 44, D1087–D1093. [Google Scholar] [CrossRef] [PubMed]
- Hancock, R.E.; Sahl, H.G. Antimicrobial and host-defense peptides as new anti-infective therapeutic strategies. Nat. Biotechnol. 2006, 24, 1551–1557. [Google Scholar] [CrossRef] [PubMed]
- Shlyapnikov, Y.M.; Andreev, Y.A.; Kozlov, S.A.; Vassilevski, A.A.; Grishin, E.V. Bacterial production of latarcin 2a, a potent antimicrobial peptide from spider venom. Protein Expr. Purif. 2008, 60, 89–95. [Google Scholar] [CrossRef] [PubMed]
- Verdonck, F.; Bosteels, S.; Desmet, J.; Moerman, L.; Noppe, W.; Willems, J.; Tytgat, J.; Van Der Walt, J. A novel class of pore-forming peptides in the venom of Parabuthus schlechteri Purcell (scorpions: Buthidae). Cimbebasia 2000, 16, 247–260. [Google Scholar]
- Gomes, V.M.; Carvalho, A.O.; Da Cunha, M.; Keller, M.N.; Bloch, C.; Deolindo, P.; Alves, E.W. Purification and characterization of a novel peptide with antifungal activity from Bothrops jararaca venom. Toxicon 2005, 45, 817–827. [Google Scholar] [CrossRef] [PubMed]
- Páramo, L.; Lomonte, B.; Pizarro-Cerdá, J.; Bengoechea, J.A.; Gorvel, J.P.; Moreno, E. Bactericidal activity of Lys49 and Asp49 myotoxic phospholipases A2 from Bothrops asper snake venom. Eur. J. Biochem. 1998, 253, 452–461. [Google Scholar] [CrossRef] [PubMed]
- Costa, T.R.; Menaldo, D.L.; Oliveira, C.Z.; Santos-Filho, N.A.; Teixeira, S.S.; Nomizo, A.; Fuly, A.L.; Monteiro, M.C.; de Souza, B.M.; Palma, M.S.; et al. Myotoxic phospholipases A2 isolated from Bothrops brazili snake venom and synthetic peptides derived from their C-terminal region: Cytotoxic effect on microorganism and tumor cells. Peptides 2008, 29, 1645–1656. [Google Scholar] [CrossRef] [PubMed]
- Homsi-Brandeburgo, M.I.; Queiroz, L.S.; Santo-Neto, H.; Rodrigues-Simioni, L.; Giglio, J.R. Fractionation of Bothrops jararacussu snake venom: Partial chemical characterization and biological activity of bothropstoxin. Toxicon 1988, 26, 615–627. [Google Scholar] [CrossRef]
- Aragão, E.A.; Chioato, L.; Ward, R.J. Permeabilization of E. coli K12 inner and outer membranes by bothropstoxin-I, a Lys49 phospholipase A2 from Bothrops jararacussu. Toxicon 2008, 51, 538–546. [Google Scholar] [CrossRef] [PubMed]
- Santos-Filho, N.A.; Lorenzon, E.N.; Ramos, M.A.; Santos, C.T.; Piccoli, J.P.; Bauab, T.M.; Fusco-Almeida, A.M.; Cilli, E.M. Synthesis and characterization of an antibacterial and non-toxic dimeric peptide derived from the C-terminal region of bothropstoxin-I. Toxicon 2015, 103, 160–168. [Google Scholar] [CrossRef] [PubMed]
- Dempsey, C.E.; Ueno, S.; Avison, M.B. Enhanced membrane permeabilization and antibacterial activity of a disulfide-dimerized magainin analogue. Biochemistry 2003, 42, 402–409. [Google Scholar] [CrossRef] [PubMed]
- Zhu, W.L.; Shin, S.Y. Effects of dimerization of the cell-penetrating peptide Tat analog on antimicrobial activity and mechanism of bactericidal action. J. Pept. Sci. 2009, 15, 345–352. [Google Scholar] [CrossRef] [PubMed]
- Lorenzón, E.N.; Sanches, P.R.; Nogueira, L.G.; Bauab, T.M.; Cilli, E.M. Dimerization of aurein 1.2: Effects in structure, antimicrobial activity and aggregation of Candida albicans cells. Amino Acids 2013, 44, 1521–1528. [Google Scholar] [CrossRef] [PubMed]
- Lorenzón, E.N.; Cespedes, G.F.; Vicente, E.F.; Nogueira, L.G.; Bauab, T.M.; Castro, M.S.; Cilli, E.M. Effects of dimerization on the structure and biological activity of antimicrobial peptide Ctx-Ha. Antimicrob. Agents Chemother. 2012, 56, 3004–3010. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, L.T.; Chau, J.K.; Perry, N.A.; de Boer, L.; Zaat, S.A.; Vogel, H.J. Serum stabilities of short tryptophan- and arginine-rich antimicrobial peptide analogs. PLoS ONE 2010, 5, e12684. [Google Scholar] [CrossRef] [PubMed]
- Cudic, M.; Condie, B.A.; Weiner, D.J.; Lysenko, E.S.; Xiang, Z.Q.; Insug, O.; Bulet, P.; Otvos, L. Development of novel antibacterial peptides that kill resistant isolates. Peptides 2002, 23, 2071–2083. [Google Scholar] [CrossRef]
- Falciani, C.; Lozzi, L.; Pini, A.; Corti, F.; Fabbrini, M.; Bernini, A.; Lelli, B.; Niccolai, N.; Bracci, L. Molecular basis of branched peptides resistance to enzyme proteolysis. Chem. Biol. Drug Des. 2007, 69, 216–221. [Google Scholar] [CrossRef] [PubMed]
- Jambunathan, K.; Galande, A.K. Design of a serum stability tag for bioactive peptides. Protein Pept. Lett. 2014, 21, 32–38. [Google Scholar] [CrossRef] [PubMed]
- Yi, J.; Liu, Z.; Craft, D.; O’Mullan, P.; Ju, G.; Gelfand, C.A. Intrinsic peptidase activity causes a sequential multi-step reaction (SMSR) in digestion of human plasma peptides. J. Proteom. Res. 2008, 7, 5112–5118. [Google Scholar] [CrossRef] [PubMed]
- Zhu, P.; Bowden, P.; Zhang, D.; Marshall, J.G. Mass spectrometry of peptides and proteins from human blood. Mass Spectrom. Rev. 2011, 30, 685–732. [Google Scholar] [CrossRef] [PubMed]
- Walker, J.R.; Altman, R.K.; Warren, J.W.; Altman, E. Using protein-based motifs to stabilize peptides. J. Pept. Res. 2003, 62, 214–226. [Google Scholar] [CrossRef] [PubMed]
- Perryman, M.B.; Knell, J.D.; Roberts, R. Carboxypeptidase-catalyzed hydrolysis of C-terminal lysine: Mechanism for in vivo production of multiple forms of creatine kinase in plasma. Clin. Chem. 1984, 30, 662–664. [Google Scholar] [PubMed]
- Turner, J.; Cho, Y.; Dinh, N.N.; Waring, A.J.; Lehrer, R.I. Activities of LL-37, a cathelin-associated antimicrobial peptide of human neutrophils. Antimicrob. Agents Chemother. 1998, 42, 2206–2214. [Google Scholar] [PubMed]
- Yu, H.Y.; Tu, C.H.; Yip, B.S.; Chen, H.L.; Cheng, H.T.; Huang, K.C.; Lo, H.J.; Cheng, J.W. Easy strategy to increase salt resistance of antimicrobial peptides. Antimicrob. Agents Chemother. 2011, 55, 4918–4921. [Google Scholar] [CrossRef] [PubMed]
- Chu, H.L.; Yu, H.Y.; Yip, B.S.; Chih, Y.H.; Liang, C.W.; Cheng, H.T.; Cheng, J.W. Boosting salt resistance of short antimicrobial peptides. Antimicrob. Agents Chemother. 2013, 57, 4050–4052. [Google Scholar] [CrossRef] [PubMed]
- Mohanram, H.; Bhattacharjya, S. Salt-resistant short antimicrobial peptides. Biopolymers 2016, 106, 345–356. [Google Scholar] [CrossRef] [PubMed]
- Mohamed, M.F.; Abdelkhalek, A.; Seleem, M.N. Evaluation of short synthetic antimicrobial peptides for treatment of drug-resistant and intracellular Staphylococcus aureus. Sci. Rep. 2016, 6, 29707. [Google Scholar] [CrossRef] [PubMed]
- Dabul, A.N.; Kos, V.N.; Gilmore, M.S.; Camargo, I.L. Draft genome sequence of methicillin-resistant Staphylococcus aureus strain SA16, representative of an endemic clone from a Brazilian hospital. Genome Announc. 2013, 1, e00754-13. [Google Scholar] [CrossRef] [PubMed]
- Oliveira, G.A.; Faria, J.B.; Levy, C.E.; Mamizuka, E.M. Characterization of the brazilian endemic clone of methicillin-resistant Staphylococcus aureus (MRSA) from hospitals throughout Brazil. Braz. J. Infect. Dis. 2001, 5, 163–170. [Google Scholar] [CrossRef] [PubMed]
- Abreu, A.C.; Tavares, R.R.; Borges, A.; Mergulhão, F.; Simões, M. Current and emergent strategies for disinfection of hospital environments. J. Antimicrob. Chemother. 2013, 68, 2718–2732. [Google Scholar] [CrossRef] [PubMed]
- Saidel-Odes, L.; Borer, A. Limiting and controlling carbapenem-resistant Klebsiella pneumoniae. Infect. Drug Resist. 2013, 7, 9–14. [Google Scholar] [PubMed]
- Pendleton, J.N.; Gorman, S.P.; Gilmore, B.F. Clinical relevance of the ESKAPE pathogens. Expert Rev. Anti-Infect. Ther. 2013, 11, 297–308. [Google Scholar] [CrossRef] [PubMed]
- Gong, P.; Cheng, M.; Li, X.; Jiang, H.; Yu, C.; Kahaer, N.; Li, J.; Zhang, L.; Xia, F.; Hu, L.; et al. Characterization of Enterococcus faecium bacteriophage IME-EFm5 and its endolysin LysEFm5. Virology 2016, 492, 11–20. [Google Scholar] [CrossRef] [PubMed]
- Egido, S.H.; Ruiz, M.S.; Inés Revuelta, S.; García, I.G.; Bellido, J.L. Enterococcus faecium small colony variant endocarditis in an immunocompetent patient. New Microbes New Infect. 2016, 9, 47–49. [Google Scholar] [CrossRef] [PubMed]
- Merlo, T.P.; Dabul, A.N.G.; Camargo, I.L.B.C. Different vana elements in E. faecalis and in E. faecium suggest at least two origins of Tn1546 among VRE in a Brazilian hospital. Microb. Drug Resist. 2015, 21, 320–328. [Google Scholar] [CrossRef] [PubMed]
- Bispo, P.J.; Hofling-Lima, A.L.; Pignatari, A.C. Characterization of ocular methicillin-resistant Staphylococcus epidermidis isolates belonging predominantly to clonal complex 2 subcluster II. J. Clin. Microbiol. 2014, 52, 1412–1417. [Google Scholar] [CrossRef] [PubMed]
- Soroush, S.; Jabalameli, F.; Taherikalani, M.; Amirmozafari, N.; Imani Fooladi, A.A.; Asadollahi, K.; Beigverdi, R.; Emaneini, M. Investigation of biofilm formation ability, antimicrobial resistance and the staphylococcal cassette chromosome mec patterns of methicillin resistant Staphylococcus epidermidis with different sequence types isolated from children. Microb. Pathog. 2016, 93, 126–130. [Google Scholar] [CrossRef] [PubMed]
- O’Gara, J.P.; Humphreys, H. Staphylococcus epidermidis biofilms: Importance and implications. J. Med. Microb. 2001, 50, 582–587. [Google Scholar] [CrossRef] [PubMed]
- Fey, P.D. Modality of bacterial growth presents unique targets: How do we treat biofilm-mediated infections? Curr. Opin. Microb. 2010, 13, 610–615. [Google Scholar] [CrossRef] [PubMed]
- Thurlow, L.R.; Hanke, M.L.; Fritz, T.; Angle, A.; Aldrich, A.; Williams, S.H.; Engebretsen, I.L.; Bayles, K.W.; Horswill, A.R.; Kielian, T. Staphylococcus aureus biofilms prevent macrophage phagocytosis and attenuate inflammation in vivo. J. Immunol. 2011, 186, 6585–6596. [Google Scholar] [CrossRef] [PubMed]
- Otto, M. Staphylococcal biofilms. Curr. Top. Microb. Immunol. 2008, 322, 207–228. [Google Scholar]
- Chessa, D.; Ganau, G.; Spiga, L.; Bulla, A.; Mazzarello, V.; Campus, G.V.; Rubino, S. Staphylococcus aureus and Staphylococcus epidermidis virulence strains as causative agents of persistent infections in breast implants. PLoS ONE 2016, 11, e0146668. [Google Scholar] [CrossRef] [PubMed]
- Rubino, C.; Brongo, S.; Pagliara, D.; Cuomo, R.; Abbinante, G.; Campitiello, N.; Santanelli, F.; Chessa, D. Infections in breast implants: A review with a focus on developing countries. J. Infect. Dev. Ctries 2014, 8, 1089–1095. [Google Scholar] [CrossRef] [PubMed]
- Beenken, K.E.; Spencer, H.; Griffin, L.M.; Smeltzer, M.S. Impact of extracellular nuclease production on the biofilm phenotype of Staphylococcus aureus under in vitro and in vivo conditions. Infect. Immun. 2012, 80, 1634–1638. [Google Scholar] [CrossRef] [PubMed]
- Solis, N.; Cain, J.A.; Cordwell, S.J. Comparative analysis of Staphylococcus epidermidis strains utilizing quantitative and cell surface shaving proteomics. J. Proteom. 2016, 130, 190–199. [Google Scholar] [CrossRef] [PubMed]
- Gill, S.R.; Fouts, D.E.; Archer, G.L.; Mongodin, E.F.; Deboy, R.T.; Ravel, J.; Paulsen, I.T.; Kolonay, J.F.; Brinkac, L.; Beanan, M.; et al. Insights on evolution of virulence and resistance from the complete genome analysis of an early methicillin-resistant Staphylococcus aureus strain and a biofilm-producing methicillin-resistant Staphylococcus epidermidis strain. J. Bacteriol. 2005, 187, 2426–2438. [Google Scholar] [CrossRef] [PubMed]
- Merrifield, R.B. Solid phase peptide synthesis. I. The synthesis of a tetrapeptide. J. Am. Chem. Soc. 1963, 85, 2149–2154. [Google Scholar] [CrossRef]
- Kaiser, E.; Colescott, R.L.; Bossinger, C.D.; Cook, P.I. Color test for detection of free terminal amino groups in the solid-phase synthesis of peptides. Anal. Biochem. 1970, 34, 595–598. [Google Scholar] [CrossRef]
- Castro, M.S.; Ferreira, T.C.; Cilli, E.M.; Crusca, E.; Mendes-Giannini, M.J.; Sebben, A.; Ricart, C.A.; Sousa, M.V.; Fontes, W. Hylin a1, the first cytolytic peptide isolated from the arboreal South American frog Hypsiboas albopunctatus (“spotted treefrog”). Peptides 2009, 30, 291–296. [Google Scholar] [CrossRef] [PubMed]
- Clinical and Laboratory Standards Institute (CLSI). Performance Standards for Antimicrobial Susceptibility Testing; 33th Informational Supplement; CLSI Document m100-s24; Clinical and Laboratory Standards Institute: Wayne, PA, USA, 2014; Volume 34. [Google Scholar]
- Pankey, G.; Sabath, L. Clinical relevance of bacteriostatic versus bactericidal mechanisms of action in the treatment of gram-positive bacterial infections. Clin. Infect. Dis. 2004, 38, 864–870. [Google Scholar] [CrossRef] [PubMed]
- Qin, N.; Tan, X.; Jiao, Y.; Liu, L.; Zhao, W.; Yang, S.; Jia, A. RNA-seq-based transcriptome analysis of methicillin-resistant Staphylococcus aureus biofilm inhibition by ursolic acid and resveratrol. Sci. Rep. 2014, 4, 5467. [Google Scholar] [CrossRef] [PubMed]
- Woolfrey, B.F.; Lally, R.T.; Ederer, M.N. Evaluation of oxacillin tolerance in Staphylococcus aureus by a novel method. Antimicrob. Agents Chemother. 1985, 28, 381–388. [Google Scholar] [CrossRef] [PubMed]
Sample Availability: Samples of the compounds used in this paper are available from the authors. |

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Peptide | Sequence | RT (min) a | Mass | ||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| (p-BthTX-I)2 | K | K | Y | R | Y | H | L | K | P | F | C | K | K | 9.18 | 3476.2 |
| des-Lys13-(p-BthTX-I)2 | K | K | Y | R | Y | H | L | K | P | F | C | K | - | 10.91 | 3219.9 |
| des-Lys12, Lys13-(p-BthTX-I)2 | K | K | Y | R | Y | H | L | K | P | F | C | - | - | 11.35 | 2963.6 |
| des-Lys1, Lys2-(p-BthTX-I)2 | - | - | Y | R | Y | H | L | K | P | F | C | K | K | 10.87 | 3219.9 |
| des-Lys1-(p-BthTX-I)2 | - | K | Y | R | Y | H | L | K | P | F | C | K | K | 10.69 | 2963.6 |
| Strain | MIC (µM) | ||||
|---|---|---|---|---|---|
| (p-BthTX-I)2 | des-Lys13-(p-BthTX-I)2 | des-Lys12, Lys13-(p-BthTX-I)2 | des-Lys1, Lys2-(p-BthTX-I)2 | des-Lys1-(p-BthTX-I)2 | |
| E. coli | 4 | 32 | 16 | 64 | 16 |
| S. aureus | 64 | 128 | 128 | 64 | 128 |
| C. albicans | >128 | >128 | >128 | >128 | >128 |
| HC50 | >128 | >128 | >128 | >128 | >128 |
| Bacterial Strains | p-BthTX-I | (p-BthTX-I)2 | des-Lys12/Lys13-(p-BthTX-I)2 | |||
|---|---|---|---|---|---|---|
| MIC (µM) | MBC (µM) | MIC (µM) | MBC (µM) | MIC (µM) | MBC (µM) | |
| S. epidermidis ATCC35984 | 128 | 512 | 16 | 64 | 32 | 32 |
| S. aureus ATCC25923 | >512 | N.D. | 512 | 512 | 128 | 256 |
| S. aureus SA16 | 512 | 512 | >512/256′ | 512 | 128 | 128 |
| S. aureus SA33 | 512/>512′ | >512 | 256/512′ | 512 | 128 | 256 |
| S. aureus SA88 | 512 | >512 | 256 | 256 | 256 | 512 |
| S. aureus SA90 | 512/>512′ | 512 | 128 | 256 | 128/512′ | 512/512′ |
| E. faecium VRE16 | 128 | >512 | 32 | >512 | 16 | 64 |
| E. faecium HSJRP8 | 32 | >512 | 8 | >512 | 8 | 32 |
| K. pneumoniae ATCC700603 | >512 | N.D. | 64 | 64 | 128/64 | 256/128 |
| K. pneumoniae ATCCBAA1705 | >512 | N.D. | 256 | 512 | 32 | 64 |
| K. pneumoniae NDM-1 | >512/512′ | >512 | 256 | 512 | 256 | 512 |
| E. coli ATCC35218 | 512 | >512 | 512 | 512 | 64 | 256 |
| E. coli CA4 | >512 | N.D. | 256 | 512 | 64 | 128 |
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Santos-Filho, N.A.; Fernandes, R.S.; Sgardioli, B.F.; Ramos, M.A.S.; Piccoli, J.P.; Camargo, I.L.B.C.; Bauab, T.M.; Cilli, E.M. Antibacterial Activity of the Non-Cytotoxic Peptide (p-BthTX-I)2 and Its Serum Degradation Product against Multidrug-Resistant Bacteria. Molecules 2017, 22, 1898. https://doi.org/10.3390/molecules22111898
Santos-Filho NA, Fernandes RS, Sgardioli BF, Ramos MAS, Piccoli JP, Camargo ILBC, Bauab TM, Cilli EM. Antibacterial Activity of the Non-Cytotoxic Peptide (p-BthTX-I)2 and Its Serum Degradation Product against Multidrug-Resistant Bacteria. Molecules. 2017; 22(11):1898. https://doi.org/10.3390/molecules22111898
Chicago/Turabian StyleSantos-Filho, Norival A., Rafaela S. Fernandes, Bruna F. Sgardioli, Matheus A. S. Ramos, Julia P. Piccoli, Ilana L. B. C. Camargo, Tais M. Bauab, and Eduardo M. Cilli. 2017. "Antibacterial Activity of the Non-Cytotoxic Peptide (p-BthTX-I)2 and Its Serum Degradation Product against Multidrug-Resistant Bacteria" Molecules 22, no. 11: 1898. https://doi.org/10.3390/molecules22111898