
Antidepressant Potential of Chlorogenic Acid-Enriched Extract from Eucommia ulmoides Oliver Bark with Neuron Protection and Promotion of Serotonin Release through Enhancing Synapsin I Expression

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
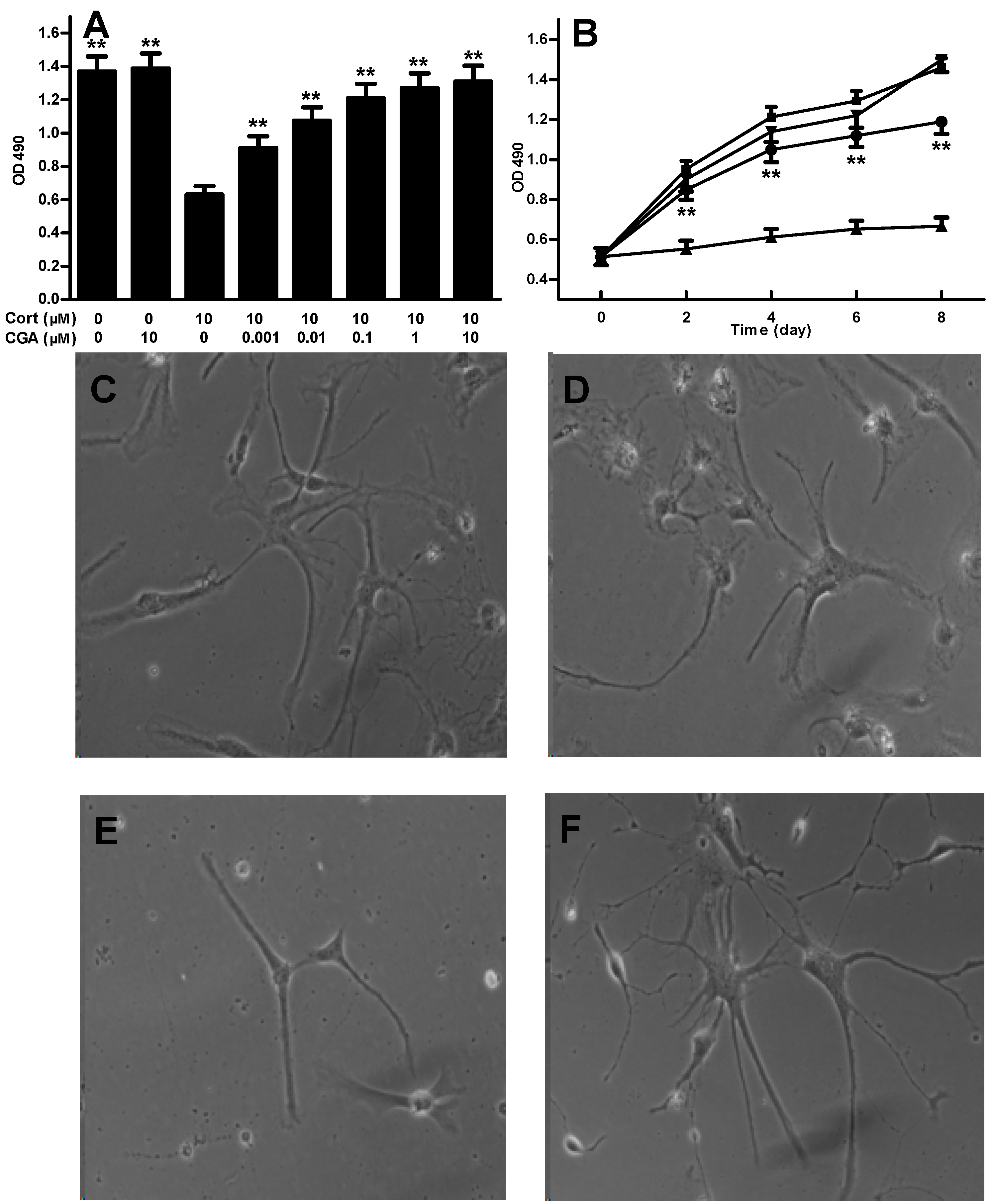
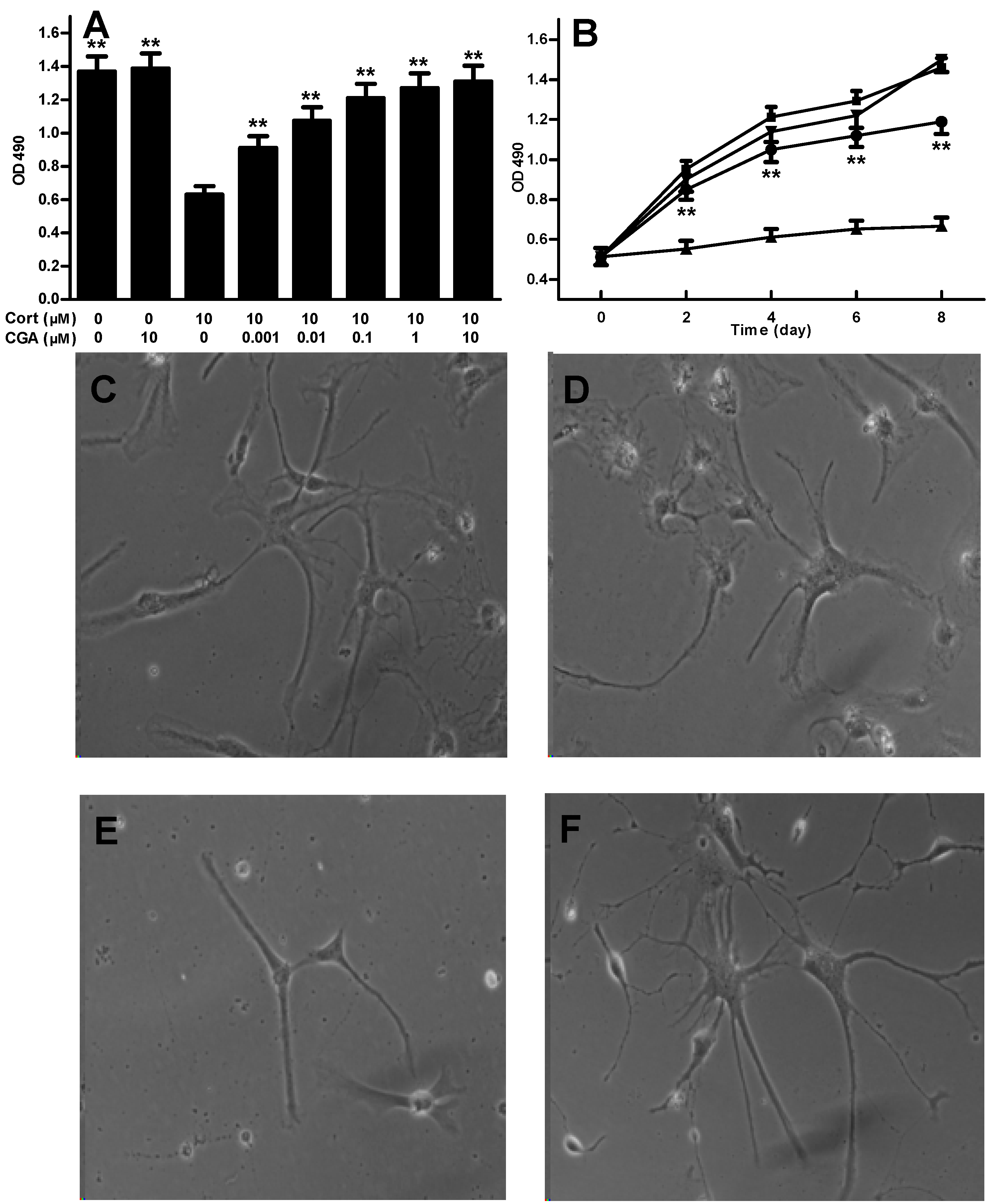
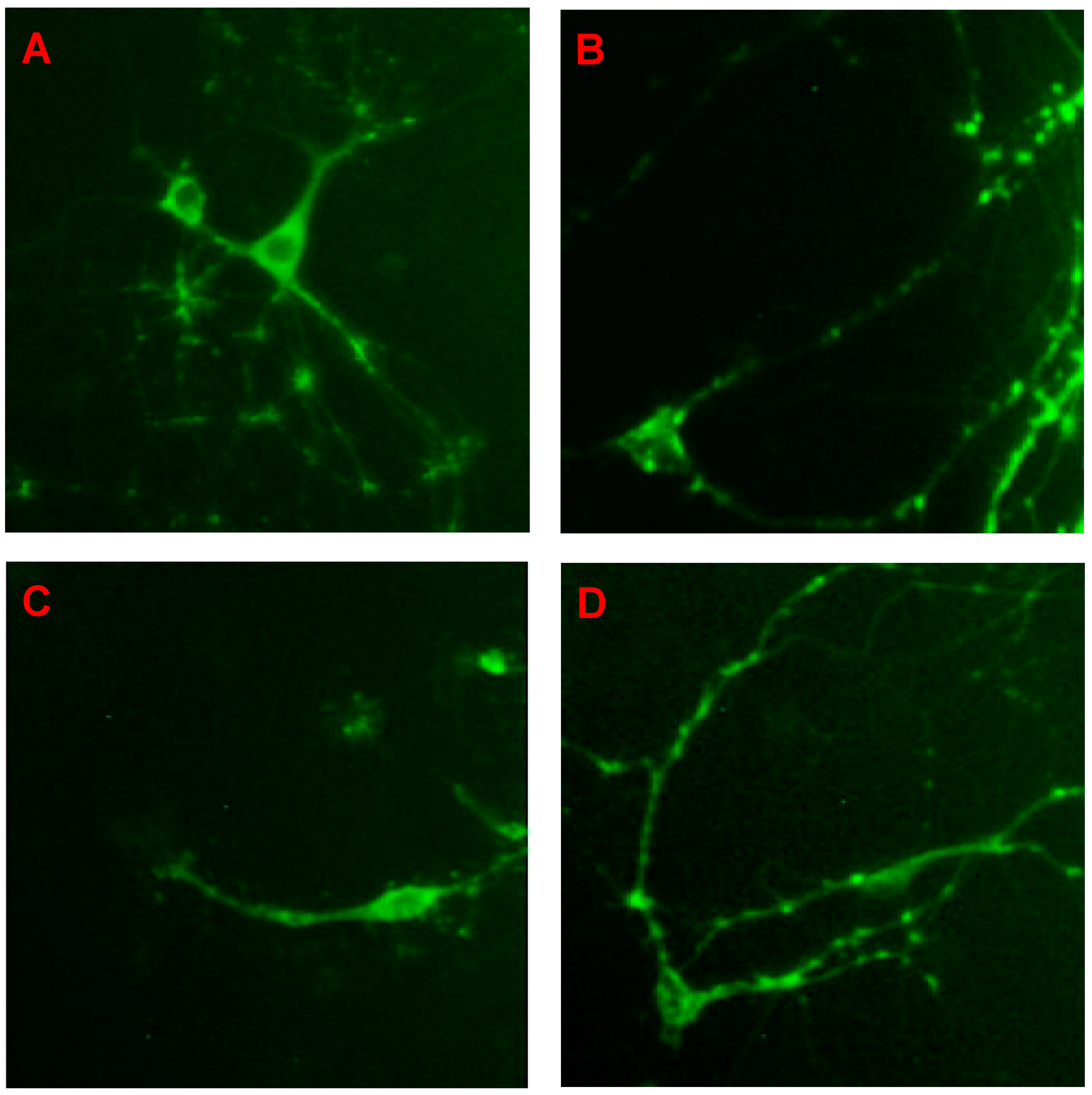
2.1. CGA Promotes the Cell Growth of Fetal Rat Raphe Neurons in Vitro
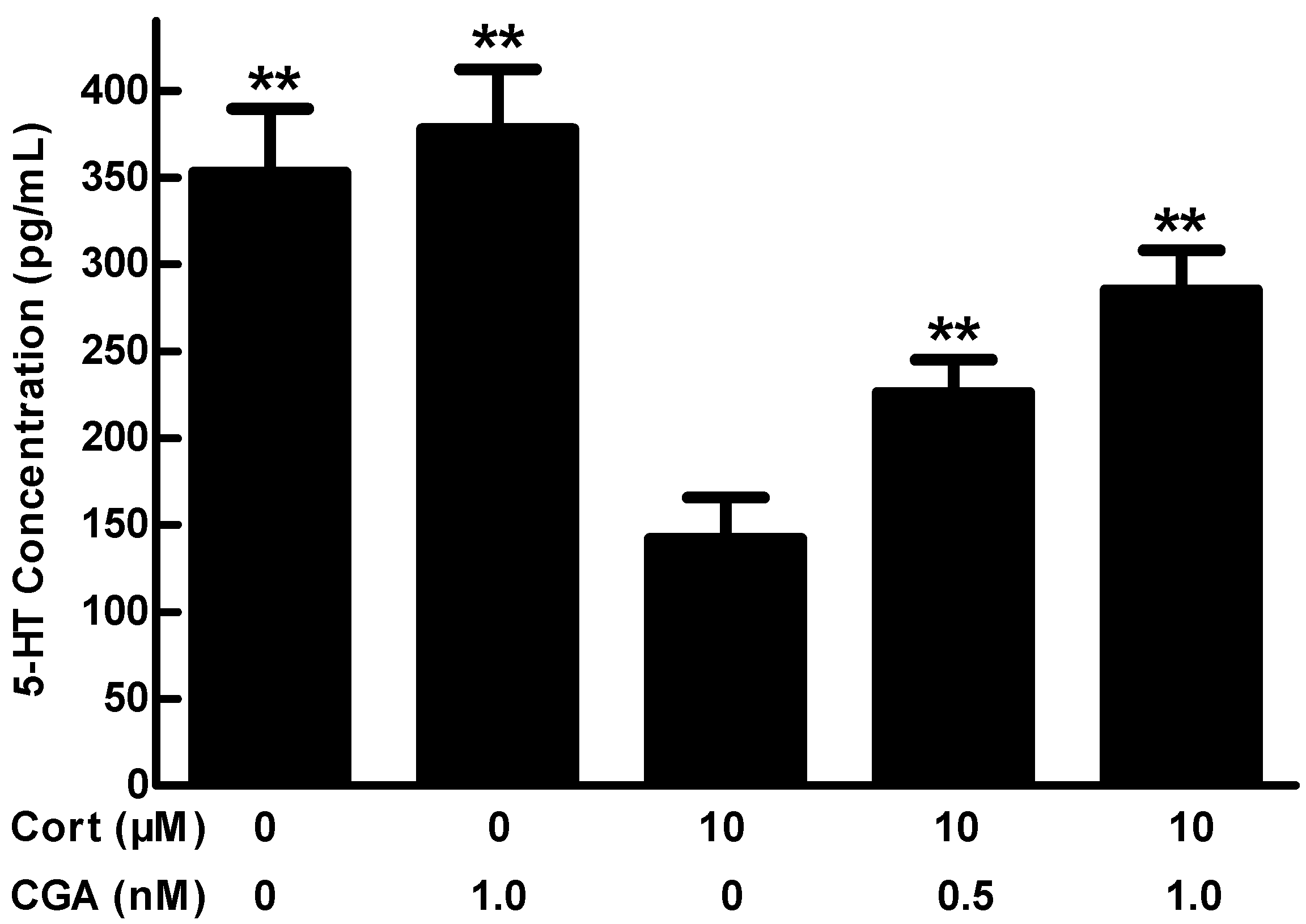
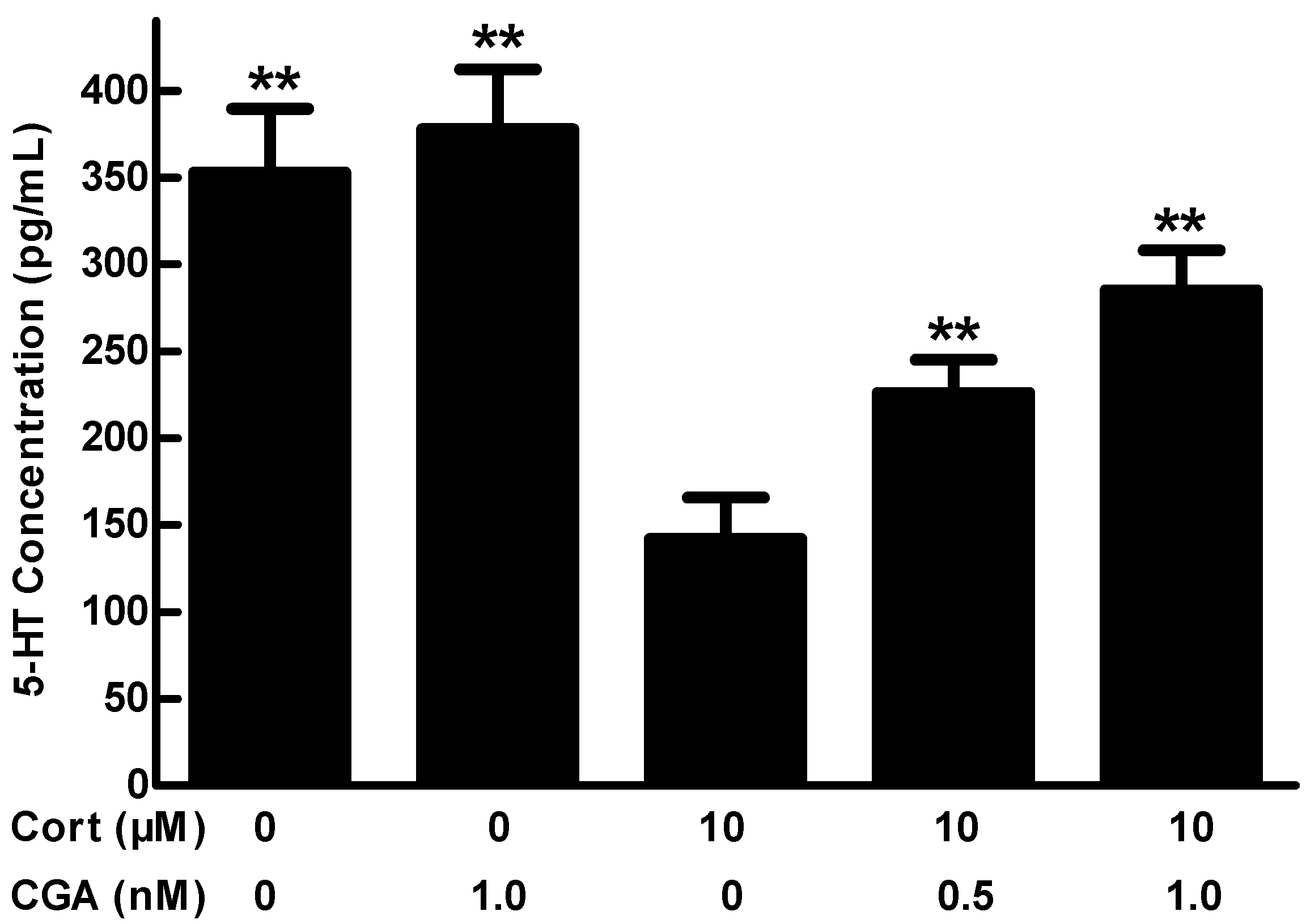
2.2. Effect of CGA on 5-HT Release in the Cells of Fetal Rat Raphe Neurons in Vitro
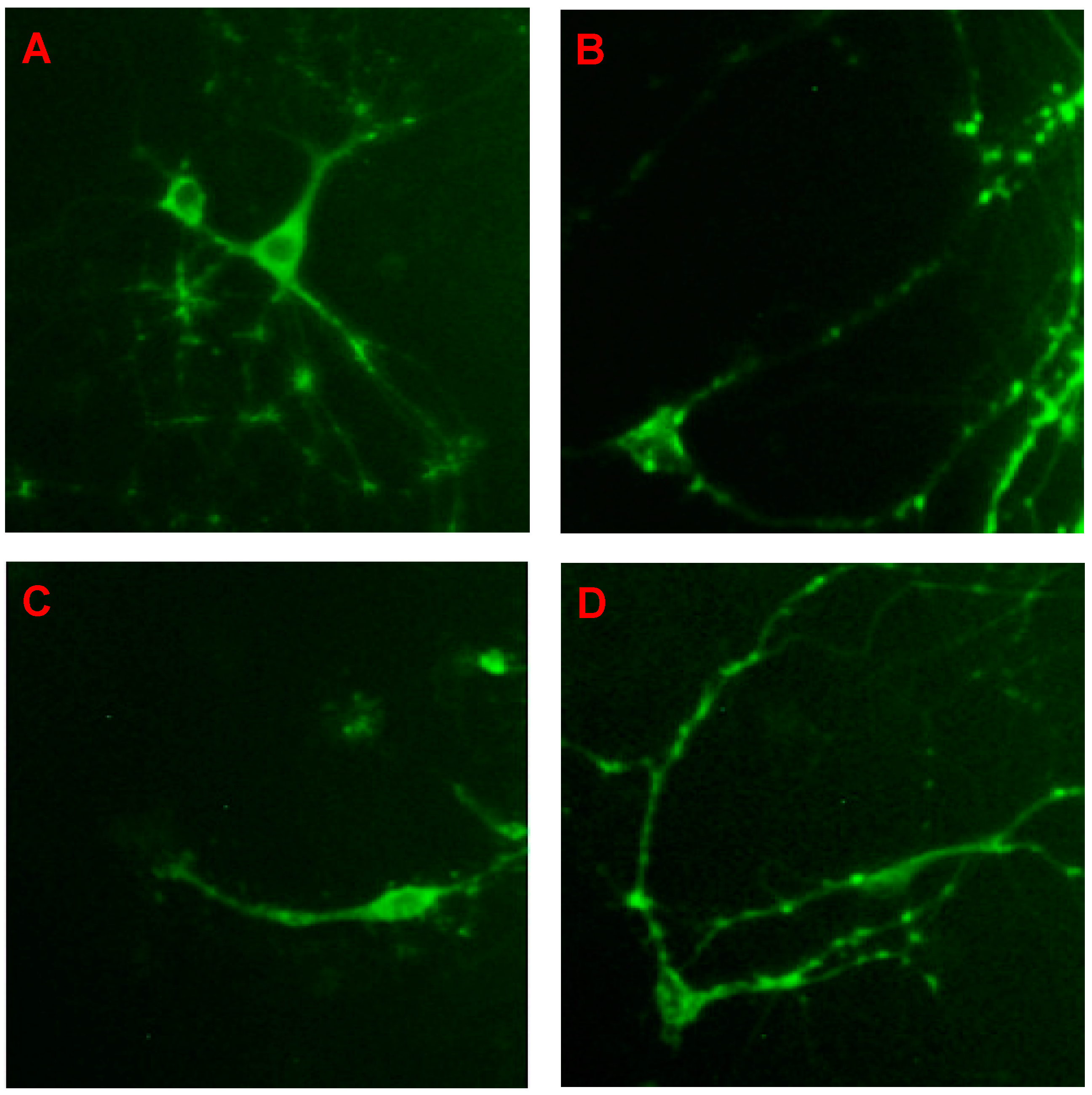
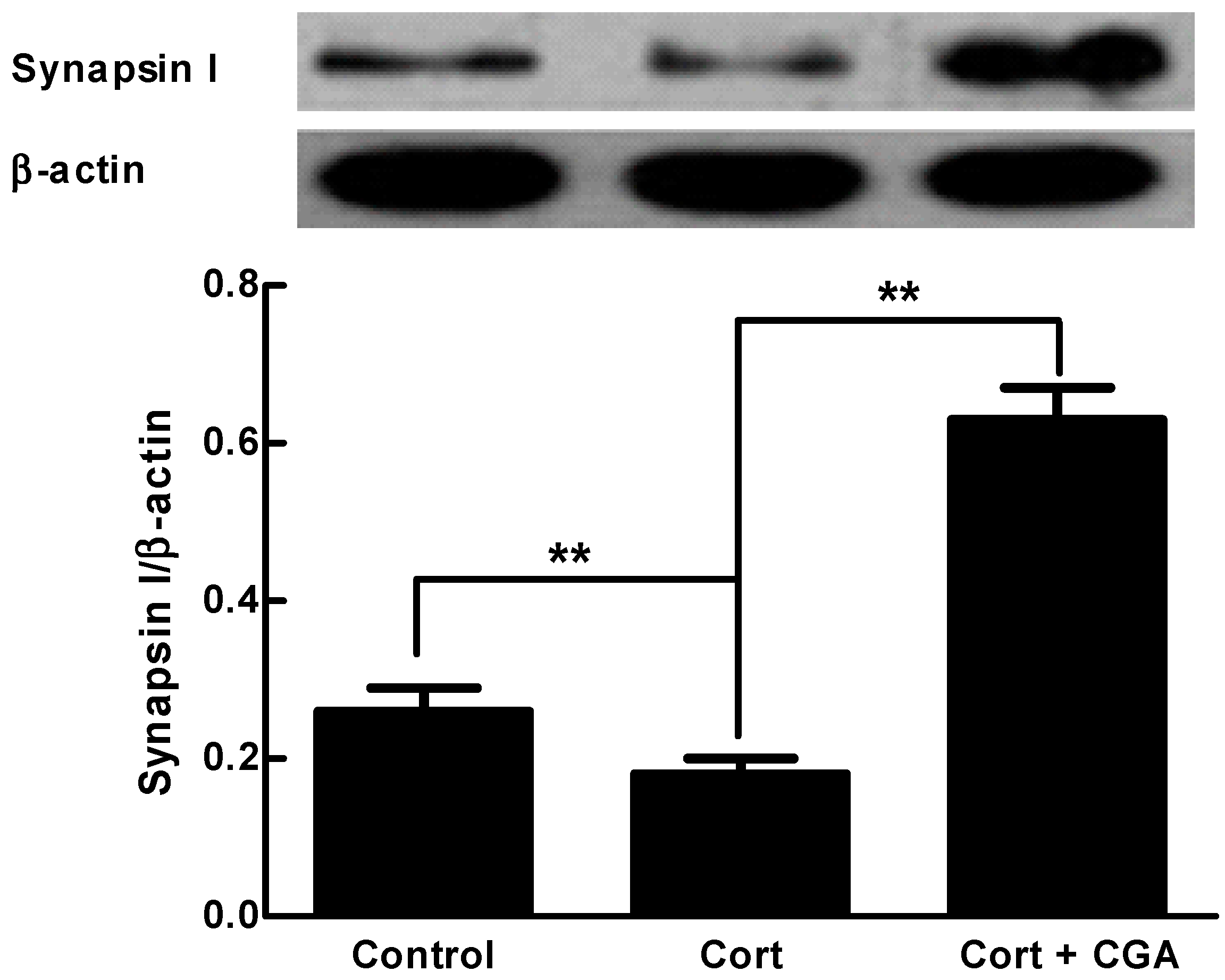
2.3. CGA Enhances the Expression of Synapsin I of the Cells of Fetal Rat Raphe Neurons in Vitro
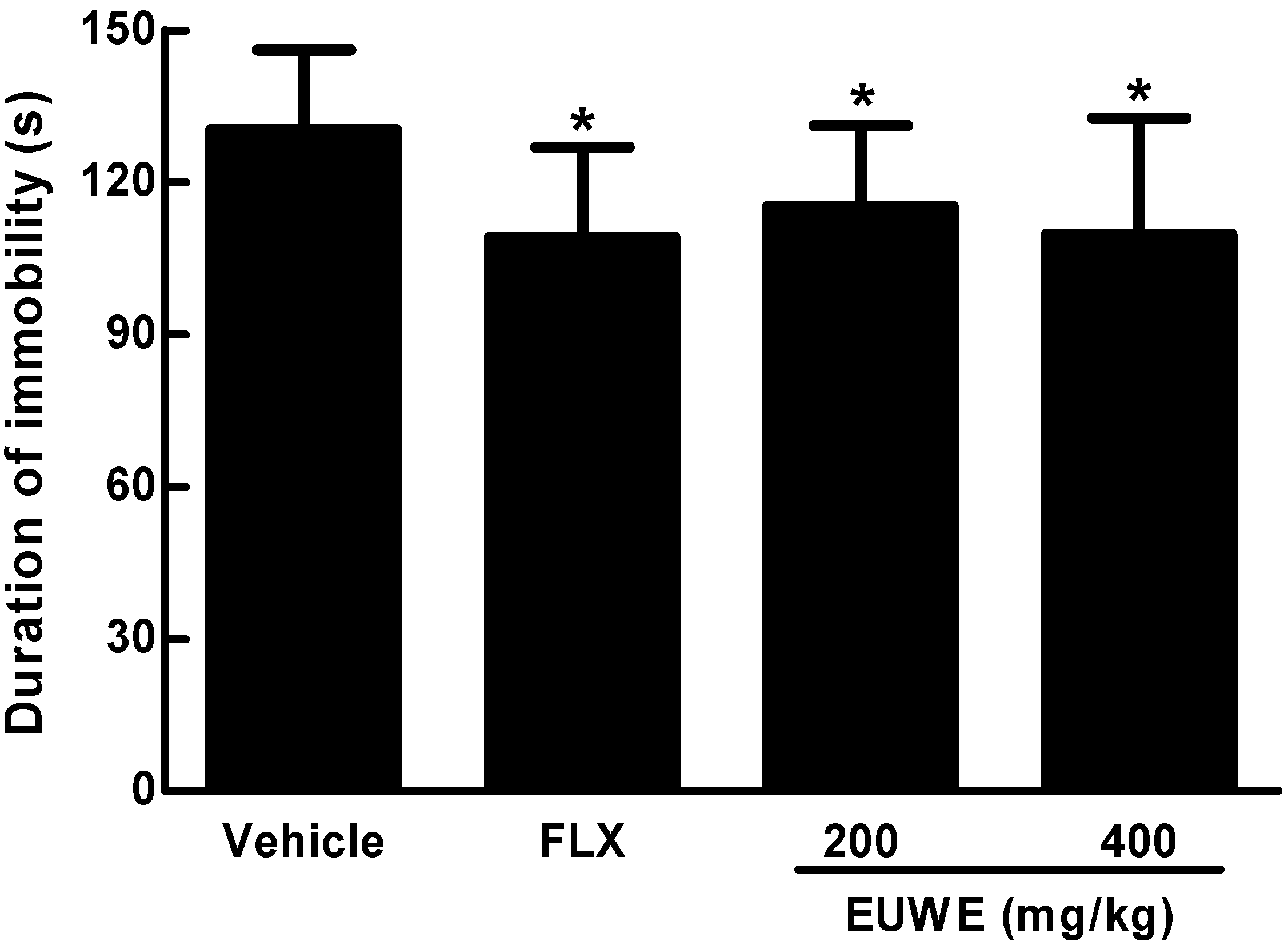
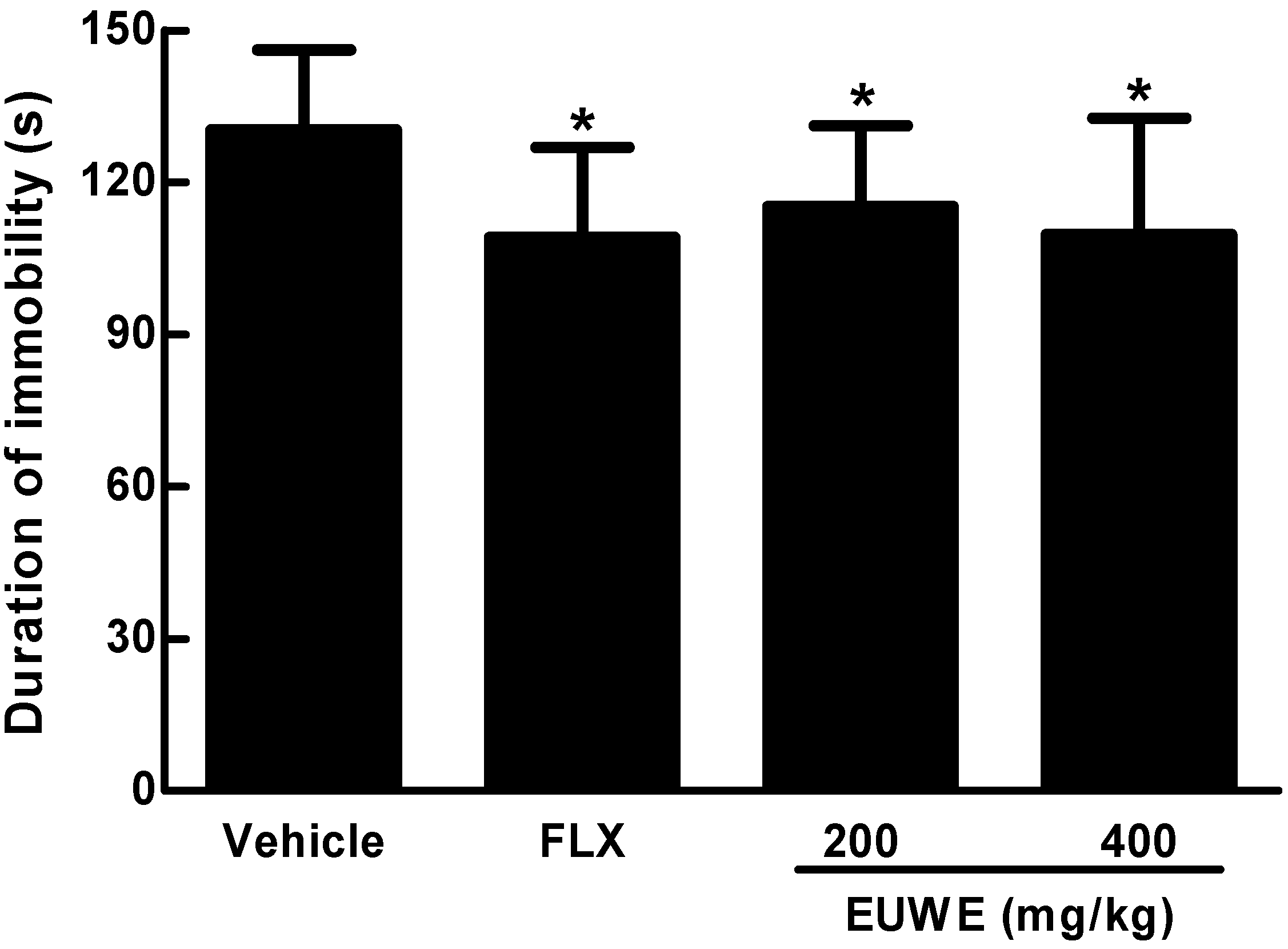
2.4. EUWE Shows Antidepressant-like Effect in the Tail Suspension Test of KM Mice in Vivo
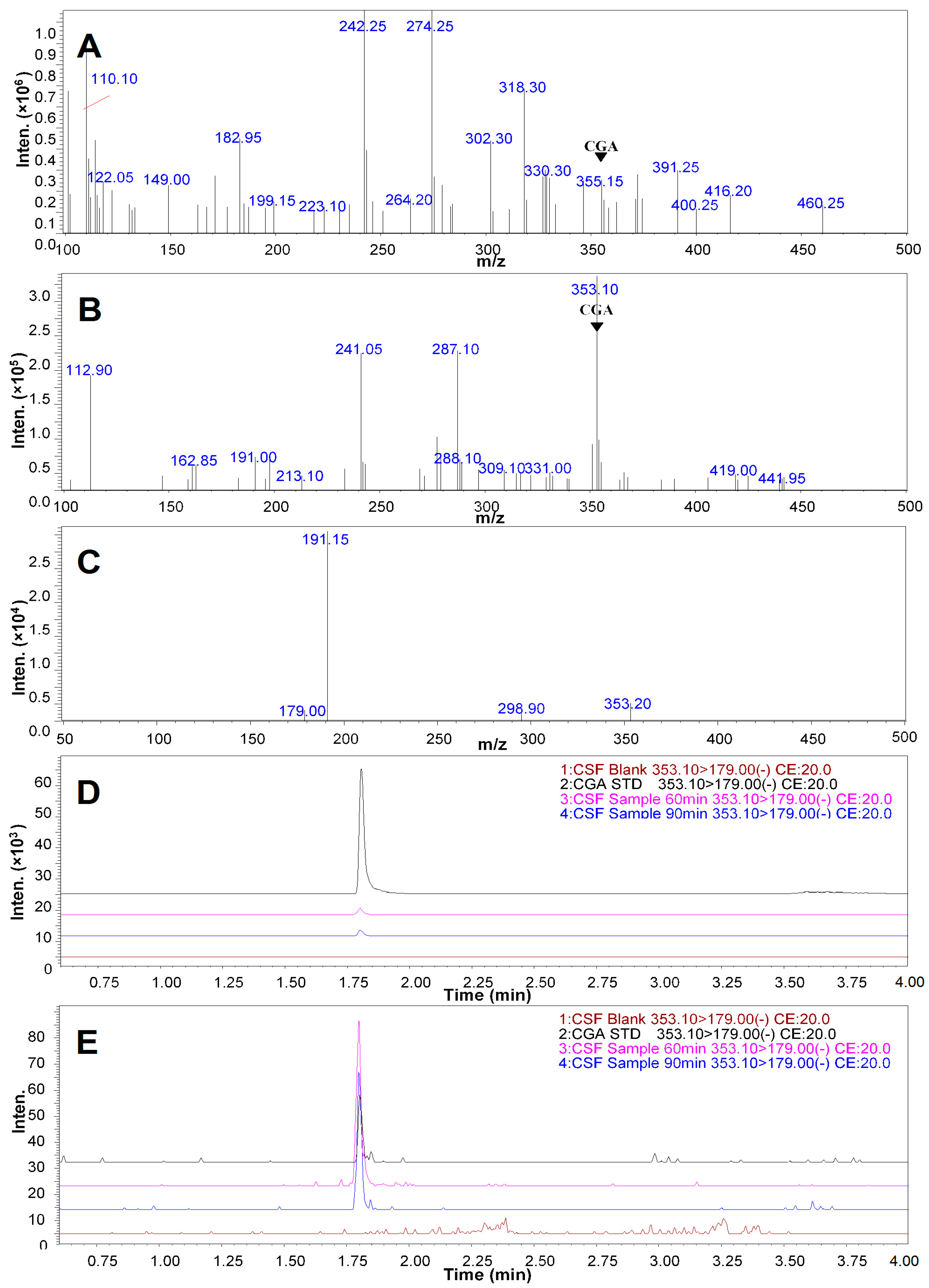
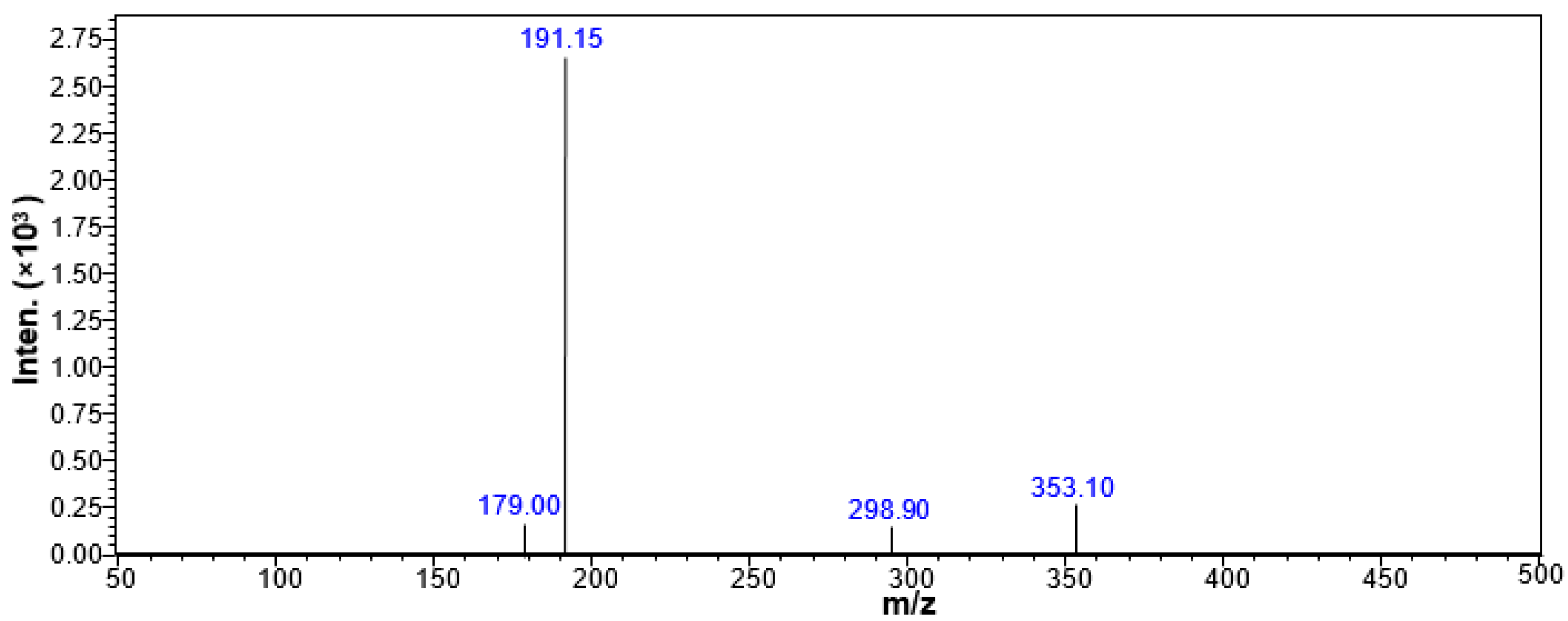
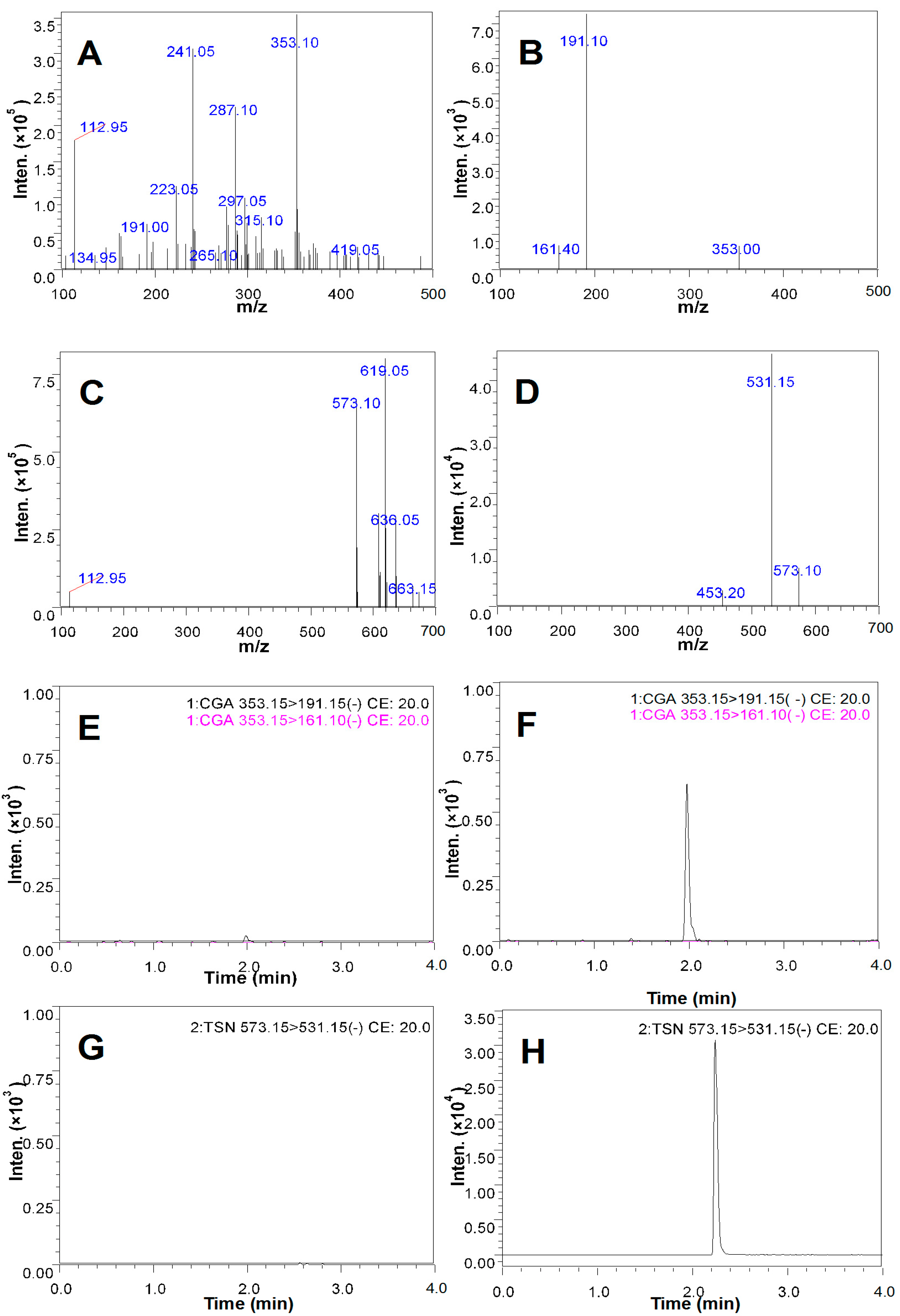
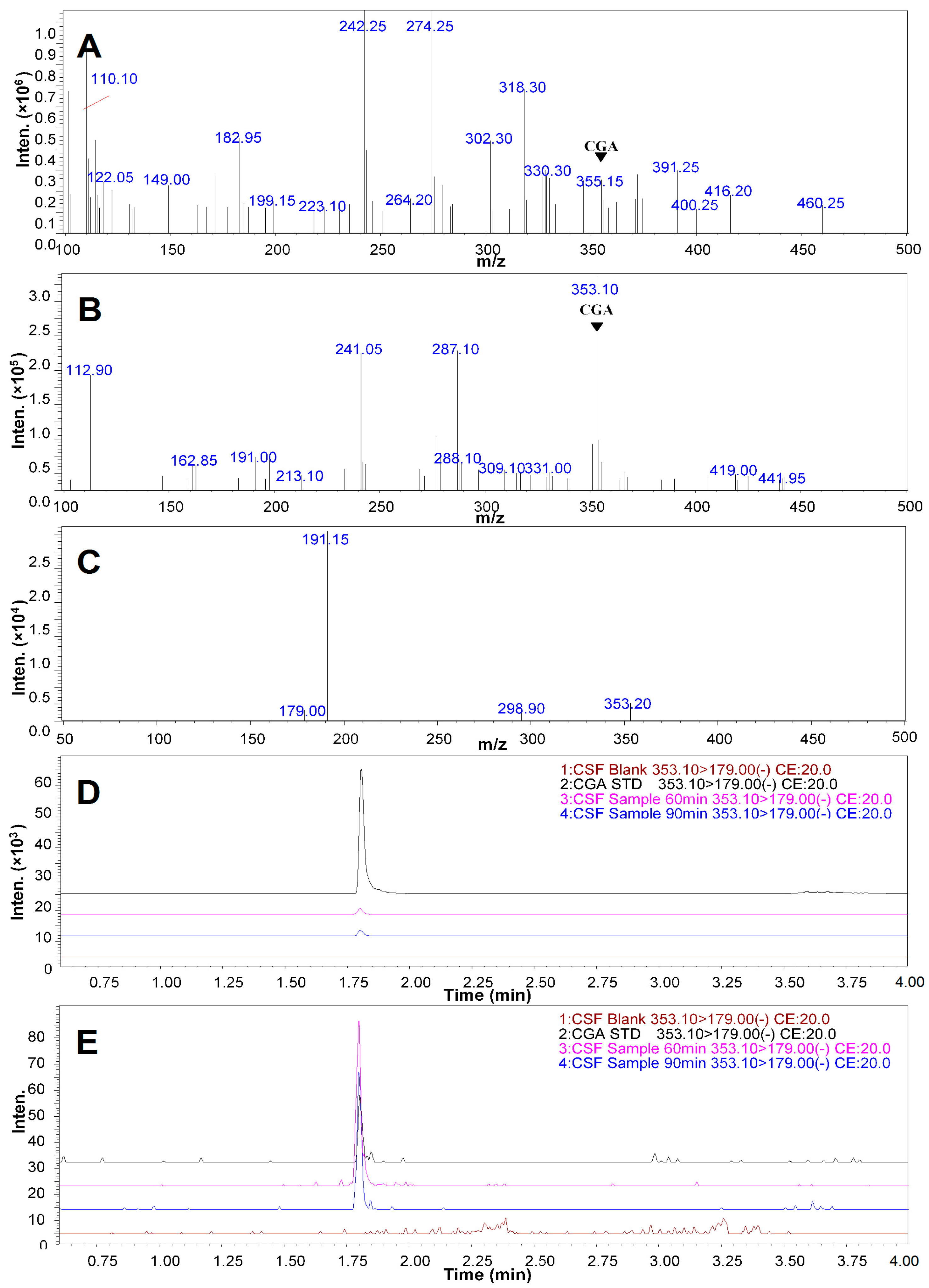
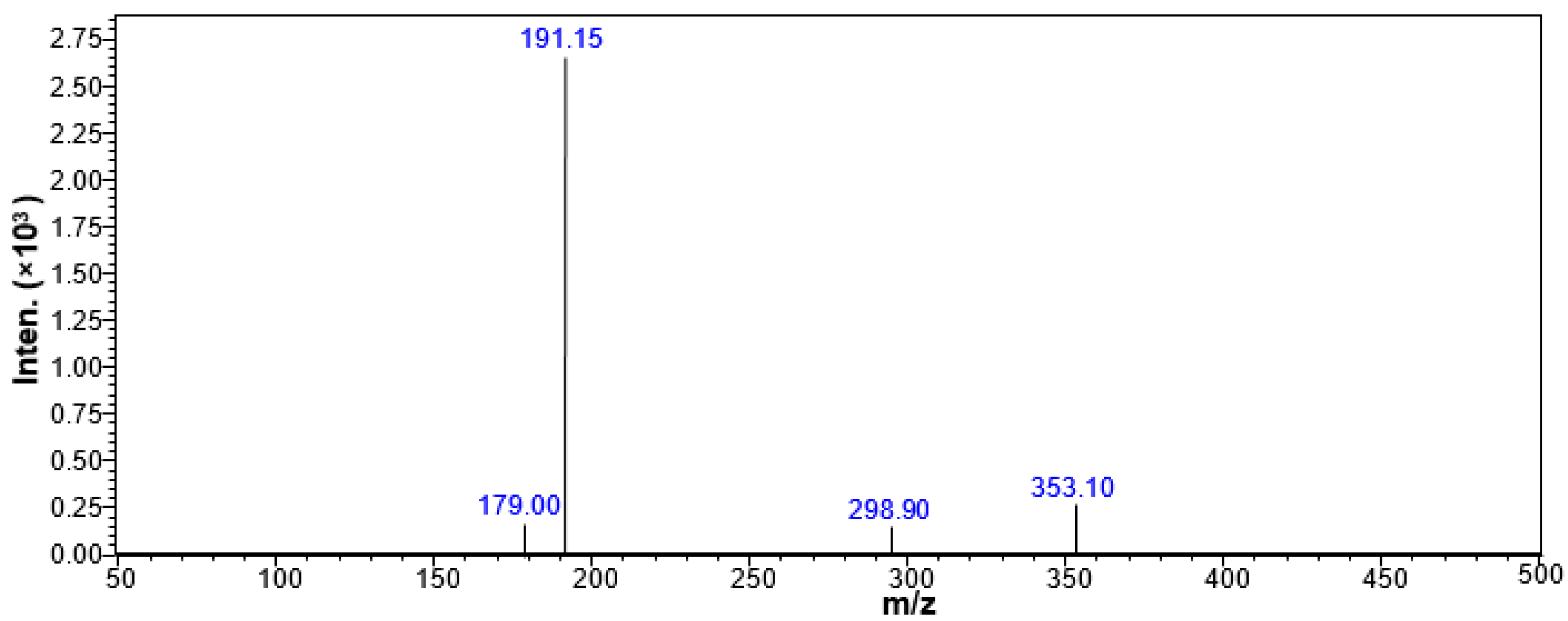
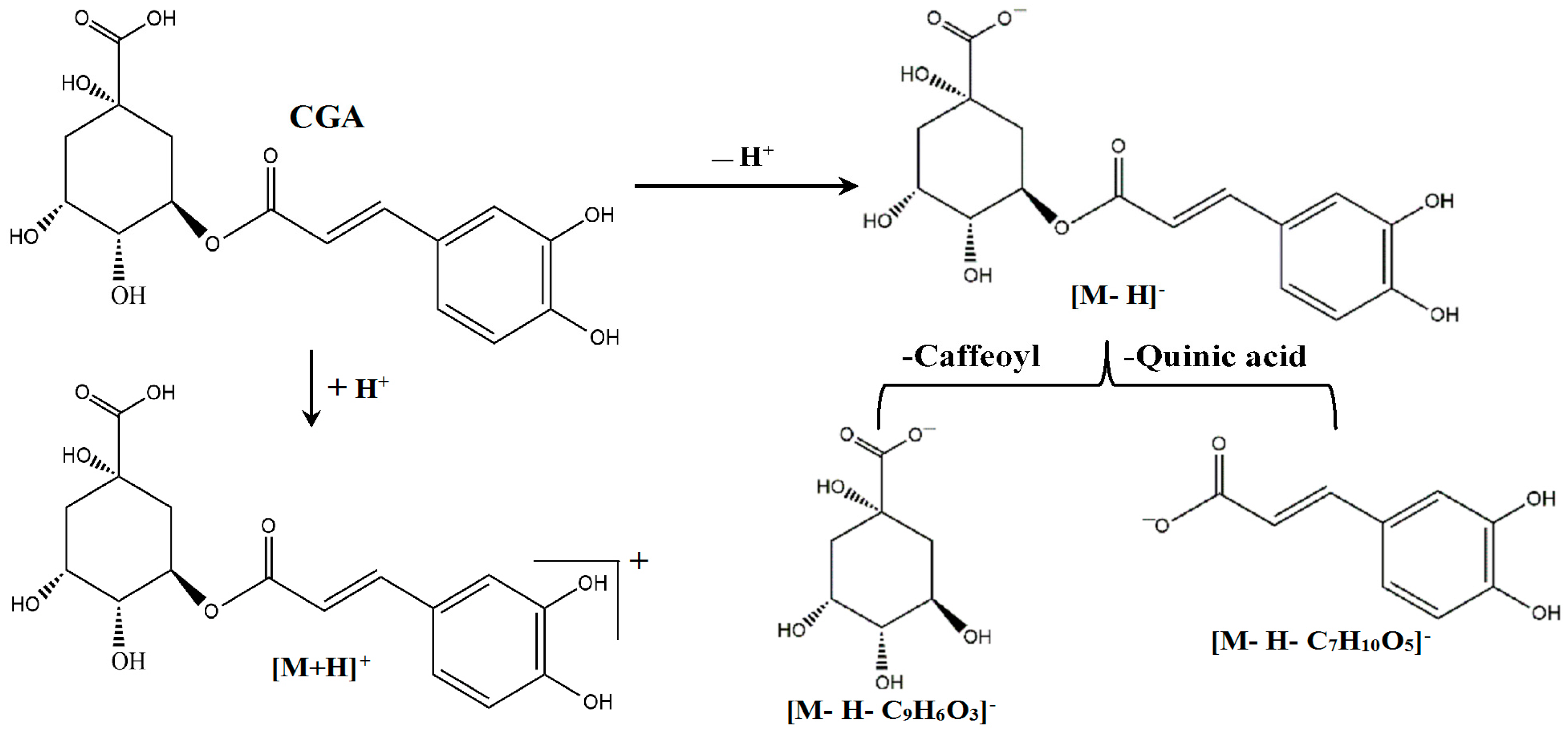
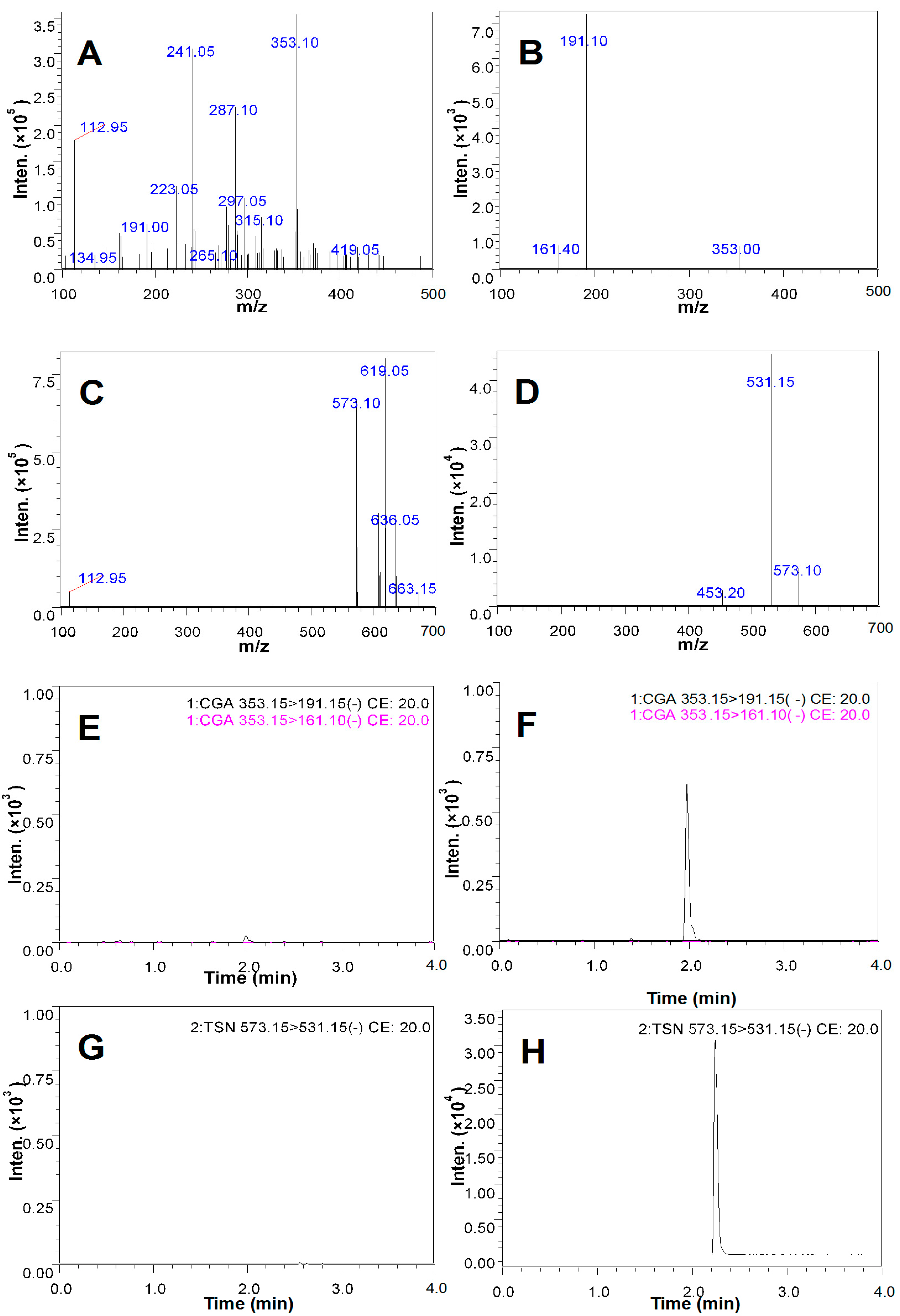
2.5. Qualitative Analysis of CGA in the CSF of the Rats Treated with CGA-Enriched Water Extract of E. ulmoides
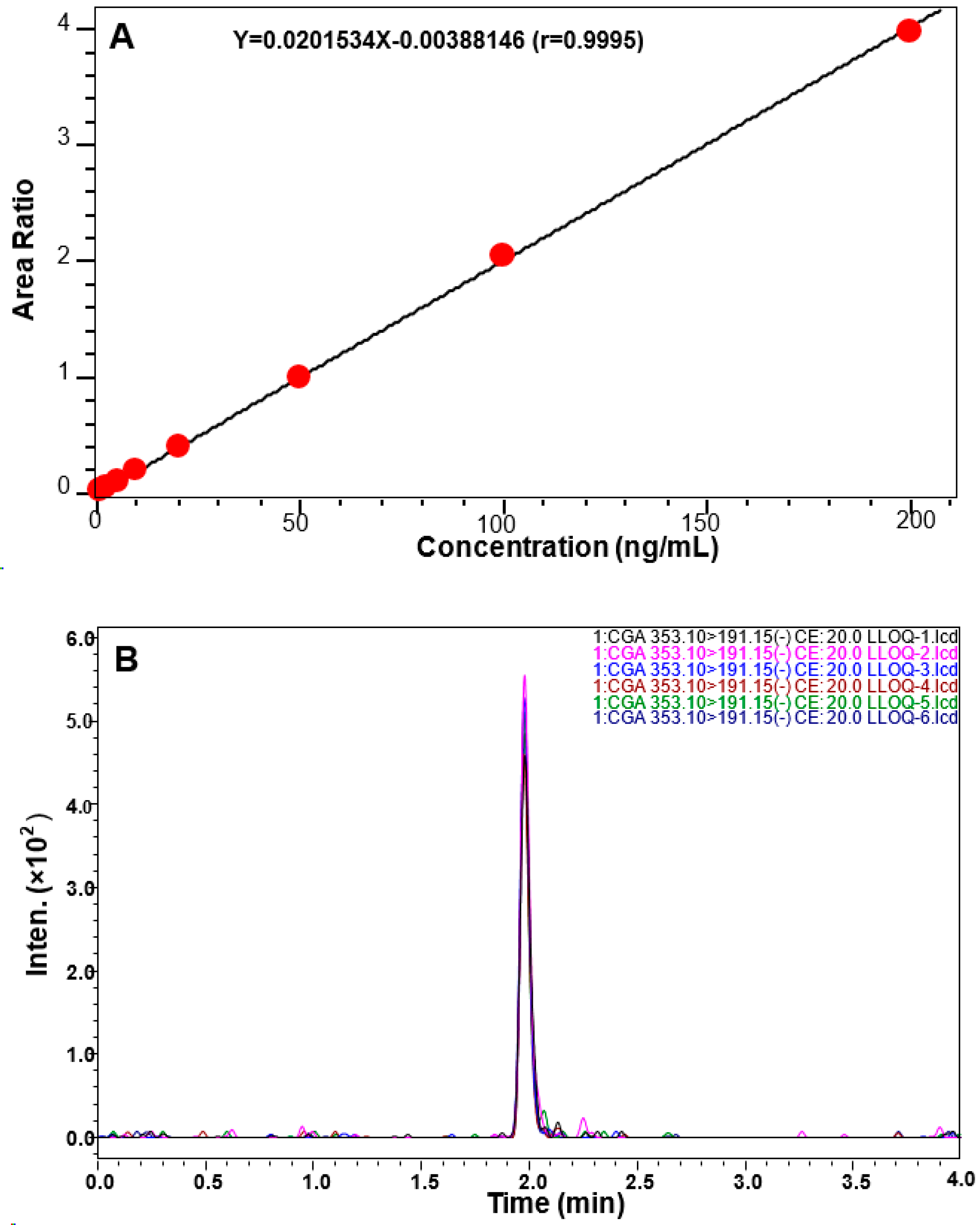
2.6. Quantitative Measurement of CGA in the CSF of Rat
3. Discussion
4. Experimental Section
4.1. Materials
4.2. Plant Material and Extraction
4.3. Animals and Sample Collection
4.4. Cell Culture and Treatment
4.5. Neurotransmitter Detection
4.6. Immunofluorescence Staining
4.7. Western Blot Analysis
4.8. Tail Suspension Test of Mice
4.9. UHPLC-ESI-MS/MS Analysis
4.10. Statistical Analysis
5. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
Abbreviations
| E. ulmoides | Eucommia ulmoides Oliver |
| TCM | Traditional Chinese Medicine |
| CGA | Chlorogenic acid |
| BLB | Blood-cerebrospinal fluid barrier |
| BBB | Blood-brain barrier |
| Cort | Corticosterone |
| 5-HT | 5-hydroxytryptamine or serotonin |
| ELISA | Enzyme-linked immunosorbent assay |
| EUWE | Water extract of E. ulmoides |
| FLX | Fluoxetine |
| TSN | Toosendanin |
| RSD | Relative standard deviation |
| ERK | Extracellular signal-regulated kinase |
| MAPK | Mitogen-associated protein kinase |
| PKA | Protein kinase A |
| CaMK | Calmodulin-dependent protein kinase |
| UHPLC | Ultrahigh performance liquid chromatography |
| DMEM | Dulbecco’s modified Eagle’s medium |
| FBS | Fetal bovine serum |
References
- He, X.; Wang, J.; Li, M.; Hao, D.; Yang, Y.; Zhang, C.; He, R.; Tao, R. Eucommia ulmoides Oliv.: Ethnopharmacology, phytochemistry and pharmacology of an important traditional Chinese medicine. J. Ethnopharmacol. 2014, 151, 78–92. [Google Scholar] [CrossRef] [PubMed]
- Xie, G.P.; Jiang, N.; Wang, S.N.; Qi, R.Z.; Wang, L.; Zhao, P.R.; Liang, L.; Yu, B. Eucommia ulmoides Oliv. bark aqueous extract inhibits osteoarthritis in a rat model of osteoarthritis. J. Ethnopharmacol. 2015, 162, 148–154. [Google Scholar] [CrossRef] [PubMed]
- Kwon, S.H.; Ma, S.X.; Hong, S.I.; Kim, S.Y.; Lee, S.Y.; Jang, C.G. Eucommia ulmoides Oliv. bark. attenuates 6-hydroxydopamine-induced neuronal cell death through inhibition of oxidative stress in SH-SY5Y cells. J. Ethnopharmacol. 2014, 152, 173–182. [Google Scholar] [CrossRef] [PubMed]
- Guo, H.; Shi, F.; Li, M.; Liu, Q.; Yu, B.; Hu, L. Neuroprotective effects of Eucommia ulmoides Oliv. and its bioactive constituent work via ameliorating the ubiquitin-proteasome system. BMC Complement. Altern. Med. 2015, 15, 151. [Google Scholar] [CrossRef] [PubMed]
- Zhang, R.; Pan, Y.L.; Hu, S.J.; Kong, X.H.; Juan, W.; Mei, Q.B. Effects of total lignans from Eucommia ulmoides barks prevent bone loss in vivo and in vitro. J. Ethnopharmacol. 2014, 155, 104–112. [Google Scholar] [CrossRef] [PubMed]
- Kwon, S.H.; Lee, H.K.; Kim, J.A.; Hong, S.I.; Kim, S.Y.; Jo, T.H.; Park, Y.I.; Lee, C.K.; Kim, Y.B.; Lee, S.Y.; et al. Neuroprotective effects of Eucommia ulmoides Oliv. Bark on amyloid beta-induced learning and memory impairments in mice. Neurosci Lett. 2011, 487, 123–127. [Google Scholar] [CrossRef] [PubMed]
- Kwon, S.H.; Ma, S.X.; Joo, H.J.; Lee, S.Y.; Jang, C.G. Inhibitory Effects of Eucommia ulmoides Oliv. Bark on Scopolamine-Induced Learning and Memory Deficits in Mice. Biomol. Ther. 2013, 21, 462–469. [Google Scholar] [CrossRef] [PubMed]
- Jin, X.; Amitani, K.; Zamami, Y.; Takatori, S.; Hobara, N.; Kawamura, N.; Hirata, T.; Wada, A.; Kitamura, Y.; Kawasaki, H. Ameliorative effect of Eucommia ulmoides Oliv. leaves extract (ELE) on insulin resistance and abnormal perivascular innervation in fructose-drinking rats. J. Ethnopharmacol. 2010, 128, 672–678. [Google Scholar] [CrossRef] [PubMed]
- Yongsheng, L.; Shumei, L.; Guodong, W. Studies on resin purification process optimization of Eucommia ulmoides Oliver and its antihypertensive effect mechanism. Afr. J. Tradit. Complement Altern. Med. 2014, 11, 475–480. [Google Scholar] [CrossRef] [PubMed]
- Peng, W.; Ge, S.; Li, D.; Mo, B.; Daochun, Q.; Ohkoshi, M. Report: Molecular basis of antibacterial activities in extracts of Eucommia ulmoides wood. Pak. J. Pharm. Sci. 2014, 27, 2133–2138. [Google Scholar] [PubMed]
- Hao, S.; Xiao, Y.; Lin, Y.; Mo, Z.; Chen, Y.; Peng, X.; Xiang, C.; Li, Y.; Li, W. Chlorogenic acid-enriched extract from Eucommia ulmoides leaves inhibits hepatic lipid accumulation through regulation of cholesterol metabolism in HepG2 cells. Pharm. Biol. 2016, 54, 251–259. [Google Scholar] [CrossRef] [PubMed]
- Fujikawa, T.; Hirata, T.; Hosoo, S.; Nakajima, K.; Wada, A.; Yurugi, Y.; Soya, H.; Matsui, T.; Yamaguchi, A.; Ogata, M.; et al. Asperuloside stimulates metabolic function in rats across several organs under high-fat diet conditions, acting like the major ingredient of Eucommia leaves with anti-obesity activity. J. Nutr. Sci. 2012, 1, e10. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.; Lee, S.; Shim, J.; Kim, H.W.; Kim, J.; Jang, Y.J.; Yang, H.; Park, J.; Choi, S.H.; Yoon, J.H.; et al. Caffeinated coffee, decaffeinated coffee, and the phenolic phytochemical chlorogenic acid up-regulate NQO1 expression and prevent H2O2-induced apoptosis in primary cortical neurons. Neurochem. Int. 2012, 60, 466–474. [Google Scholar] [CrossRef] [PubMed]
- Shen, W.; Qi, R.; Zhang, J.; Wang, Z.; Wang, H.; Hu, C.; Zhao, Y.; Bie, M.; Wang, Y.; Fu, Y.; et al. Chlorogenic acid inhibits LPS-induced microglial activation and improves survival of dopaminergic neurons. Brain Res. Bull. 2012, 88, 487–494. [Google Scholar] [CrossRef] [PubMed]
- Stefanello, N.; Schmatz, R.; Pereira, L.B.; Rubin, M.A.; da Rocha, J.B.; Facco, G.; Pereira, M.E.; Mazzanti, C.M.; Passamonti, S.; Rodrigues, M.V.; et al. Effects of chlorogenic acid, caffeine, and coffee on behavioral and biochemical parameters of diabetic rats. Mol. Cell. Biochem. 2014, 388, 277–286. [Google Scholar] [CrossRef] [PubMed]
- Kwon, S.H.; Lee, H.K.; Kim, J.A.; Hong, S.I.; Kim, H.C.; Jo, T.H.; Park, Y.I.; Lee, C.K.; Kim, Y.B.; Lee, S.Y.; et al. Neuroprotective effects of chlorogenic acid on scopolamine-induced amnesia via anti-acetylcholinesterase and anti-oxidative activities in mice. Eur. J. Pharmacol. 2010, 649, 210–217. [Google Scholar] [CrossRef] [PubMed]
- Oboh, G.; Agunloye, O.M.; Akinyemi, A.J.; Ademiluyi, A.O.; Adefegha, S.A. Comparative study on the inhibitory effect of caffeic and chlorogenic acids on key enzymes linked to Alzheimer’s disease and some pro-oxidant induced oxidative stress in rats’ brain-in vitro. Neurochem. Res. 2013, 38, 413–419. [Google Scholar] [CrossRef] [PubMed]
- Uchida, M.; Katoh, T.; Mori, M.; Maeno, T.; Ohtake, K.; Kobayashi, J.; Morimoto, Y.; Natsume, H. Intranasal administration of milnacipran in rats: Evaluation of the transport of drugs to the systemic circulation and central nervous system and the pharmacological effect. Biol. Pharm. Bull. 2011, 34, 740–747. [Google Scholar] [CrossRef] [PubMed]
- Chao, J.; Wang, H.; Zhao, W.; Zhang, M.; Zhang, L. Investigation of the inclusion behavior of chlorogenic acid with hydroxypropyl-ß-cyclodextrin. Int. J. Biol. Macromol. 2012, 50, 277–282. [Google Scholar] [CrossRef] [PubMed]
- Park, S.H.; Sim, Y.B.; Han, P.L.; Lee, J.K.; Suh, H.W. Antidepressant-like effect of chlorogenic acid isolated from Artemisia capillaris Thunb. Anim. Cells Syst. 2010, 4, 253–259. [Google Scholar] [CrossRef]
- Czéh, B.; Michaelis, T.; Watanabe, T.; Frahm, J.; de Biurrun, G.; van Kampen, M.; Bartolomucci, A.; Fuchs, E. Stress-induced changes in cerebral metabolites, hippocampal volume, and cell proliferation are prevented by antidepressant treatment with tianeptine. Proc. Natl. Acad. Sci. USA 2001, 98, 12796–12801. [Google Scholar] [CrossRef] [PubMed]
- Pariante, C.M. The role of multi-drug resistance p-glycoprotein in glucocorticoid function: studies in animals and relevance in humans. Eur. J. Pharmacol. 2008, 583, 263–271. [Google Scholar] [CrossRef] [PubMed]
- Orchinik, M.; Weiland, N.G.; McEwen, B.S. Chronic exposure to stress levels of corticosterone alters GABAA receptor subunit mRNA levels in rat hippocampus. Brain Res. Mol. Brain Res. 1995, 34, 29–37. [Google Scholar] [CrossRef]
- Belmaker, R.H.; Agam, G. Major depressive disorder. N. Engl. J. Med. 2008, 358, 55–68. [Google Scholar] [CrossRef] [PubMed]
- Ashimi, S.S. An in Vitro Characterization of the Raphe Nucleus and the Effects of SSRIs on Synaptic Neurotransmission. Ph.D. Thesis, The University of Texas Southwestern Medical Center, Dallas, TX, USA, 2010. [Google Scholar]
- Smith, T.D.; Kuczenski, R.; George-Friedman, K.; Malley, J.D.; Foote, S.L. In vivo microdialysis assessment of extracellular serotonin and dopamine levels in awake monkeys during sustained fluoxetine administration. Synapse 2000, 38, 460–470. [Google Scholar] [CrossRef]
- Valtorta, F.; Iezzi, N.; Benfenati, F.; Lu, B.; Poo, M.M.; Greengard, P. Accelerated structural maturation induced by synapsin I at developing neuromuscular synapses of Xenopus laevis. Eur. J. Neurosci. 1995, 7, 261–270. [Google Scholar] [CrossRef] [PubMed]
- Takei, Y.; Harada, A.; Takeda, S.; Kobayashi, K.; Terada, S.; Noda, T.; Takahashi, T.; Hirokawa, N. Synapsin I deficiency results in the structural change in the presynaptic terminals in the murine nervous system. J. Cell Biol. 1995, 131, 1789–1800. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chin, L.S.; Li, L.; Ferreira, A.; Kosik, K.S.; Greengard, P. Impairment of axonal development and of synaptogenesis in hippocampal neurons of synapsin I-deficient mice. Proc. Natl. Acad. Sci. USA 1995, 92, 9230–9234. [Google Scholar] [CrossRef] [PubMed]
- Petrik, D.; Lagace, D.C.; Eisch, A.J. The neurogenesis hypothesis of affective and anxiety disorders: Are we mistaking the scaffolding for the building? Neuropharmacol. 2012, 62, 21–34. [Google Scholar] [CrossRef] [PubMed]
- Eisch, A.J.; Petrik, D. Depression and hippocampal neurogenesis: a road to remission? Science. 2012, 338, 72–75. [Google Scholar] [CrossRef] [PubMed]
- Wu, L.M.; Han, H.; Wang, Q.N.; Hou, H.L.; Tong, H.; Yan, X.B.; Zhou, J.N. Mifepristone repairs region-dependent alteration of synapsin I in hippocampus in rat model of depression. Neuropsychopharmacology 2007, 32, 2500–2510. [Google Scholar] [CrossRef] [PubMed]
- Zhu, C.S.; Zhang, B.; Lin, Z.J.; Wang, X.J.; Zhou, Y.; Sun, X.X.; Xiao, M.L. Relationship between high-performance liquid chromatography fingerprints and uric acid-lowering activities of Cichorium intybus L. Molecules 2015, 20, 9455–9467. [Google Scholar] [CrossRef] [PubMed]
- Lopes-Lutz, D.; Dettmann, J.; Nimalaratne, C.; Schieber, A. Characterization and quantification of polyphenols in Amazon grape (Pourouma cecropiifolia Martius). Molecules 2010, 15, 8543–8552. [Google Scholar] [CrossRef] [PubMed]
- Lech, K.; Witkoś, K.; Jarosz, M. HPLC-UV-ESI MS/MS identification of the color constituents of sawwort (Serratula tinctoria L.). Anal. Bioanal. Chem. 2014, 406, 3703–3708. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Q.; Zhagn, J.Y.; Sui, C.L.; Shi, X.Y.; Qiao, Y.J.; Lu, J.Q. Regularity of changes in chlorogenic acids in Lonicera japonica extracts by HPLC-DAD-ESI-MS/MS. Zhongguo Zhong Yao Za Zhi (In Chinese) 2012, 37, 3564–3568. [Google Scholar] [PubMed]
- Pieribone, V.A.; Shupliakov, O.; Brodin, L.; Hilfiker-Rothenfluh, S.; Czernik, A.J.; Greengard, P. Distinct pools of synaptic vesicles in neurotransmitter release. Nature 1995, 375, 493–497. [Google Scholar] [CrossRef] [PubMed]
- Greengard, P.; Valtorta, F.; Czernik, A.J.; Benfenati, F. Synaptic vesicle phosphoproteins and regulation of synaptic function. Science. 1993, 259, 780–785. [Google Scholar] [CrossRef] [PubMed]
- Kushner, S.A.; Elgersma, Y.; Murphy, G.G.; Jaarsma, D.; van Woerden, G.M.; Hojjati, M.R.; Cui, Y.; LeBoutillier, J.C.; Marrone, D.F.; Choi, E.S.; et al. Modulation of presynaptic plasticity and learning by the H-ras/extracellular signal-regulated kinase/synapsin I signaling pathway. J. Neurosci. 2005, 25, 9721–9734. [Google Scholar] [CrossRef] [PubMed]
- Menegon, A.; Bonanomi, D.; Albertinazzi, C.; Lotti, F.; Ferrari, G.; Kao, H.T.; Benfenati, F.; Baldelli, P.; Valtorta, F. Protein kinase A-mediated synapsin I phosphorylation is a central modulator of Ca2+-dependent synaptic activity. J. Neurosci. 2006, 26, 11670–11681. [Google Scholar] [CrossRef] [PubMed]
- Fiumara, F.; Onofri, F.; Benfenati, F.; Montarolo, P.G.; Ghirardi, M. Intracellular injection of synapsin I induces neurotransmitter release in C1 neurons of Helix pomatia contacting a wrong target. Neuroscience 2001, 104, 271–280. [Google Scholar] [CrossRef]
- Pittaluga, A.; Raiteri, L.; Longordo, F.; Luccini, E.; Barbiero, V.S.; Racagni, G.; Popoli, M.; Raiteri, M. Antidepressant treatments and function of glutamate ionotropic receptors mediating amine release in hippocampus. Neuropharmacology 2007, 53, 27–36. [Google Scholar] [CrossRef] [PubMed]
- Millan, M.J. The role of monoamines in the actions of established and “novel” antidepressant agents: A critical review. Eur. J. Pharmacol. 2004, 500, 371–384. [Google Scholar] [CrossRef] [PubMed]
- Kobayashi, K.; Ikeda, Y.; Haneda, E.; Suzuki, H. Chronic fluoxetine bidirectionally modulates potentiating effects of serotonin on the hippocampal mossy fiber synaptic transmission. J. Neurosci. 2008, 28, 6272–6280. [Google Scholar] [CrossRef] [PubMed]
- Svetlov, S.I.; Prima, V.; Kirk, D.R.; Gutierrez, H.; Curley, K.C.; Hayes, R.L.; Wang, K.K. Morphologic and biochemical characterization of brain injury in a model of controlled blast overpressure exposure. J. Trauma 2010, 69, 795–804. [Google Scholar] [CrossRef] [PubMed]
- Sample Availability: Samples of the chlorogenic acid and toosendanin are available from the authors.
© 2016 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons by Attribution (CC-BY) license ( http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wu, J.; Chen, H.; Li, H.; Tang, Y.; Yang, L.; Cao, S.; Qin, D. Antidepressant Potential of Chlorogenic Acid-Enriched Extract from Eucommia ulmoides Oliver Bark with Neuron Protection and Promotion of Serotonin Release through Enhancing Synapsin I Expression. Molecules 2016, 21, 260. https://doi.org/10.3390/molecules21030260
Wu J, Chen H, Li H, Tang Y, Yang L, Cao S, Qin D. Antidepressant Potential of Chlorogenic Acid-Enriched Extract from Eucommia ulmoides Oliver Bark with Neuron Protection and Promotion of Serotonin Release through Enhancing Synapsin I Expression. Molecules. 2016; 21(3):260. https://doi.org/10.3390/molecules21030260
Chicago/Turabian StyleWu, Jianming, Haixia Chen, Hua Li, Yong Tang, Le Yang, Shousong Cao, and Dalian Qin. 2016. "Antidepressant Potential of Chlorogenic Acid-Enriched Extract from Eucommia ulmoides Oliver Bark with Neuron Protection and Promotion of Serotonin Release through Enhancing Synapsin I Expression" Molecules 21, no. 3: 260. https://doi.org/10.3390/molecules21030260