
Screening of Korean Natural Products for Anti-Adipogenesis Properties and Isolation of Kaempferol-3-O-rutinoside as a Potent Anti-Adipogenetic Compound from Solidago virgaurea


Abstract
:1. Introduction
2. Results
2.1. List of Natural Extracts That Showed 30% or More Inhibition of Adipogenesis of 3T3-L1 Cells
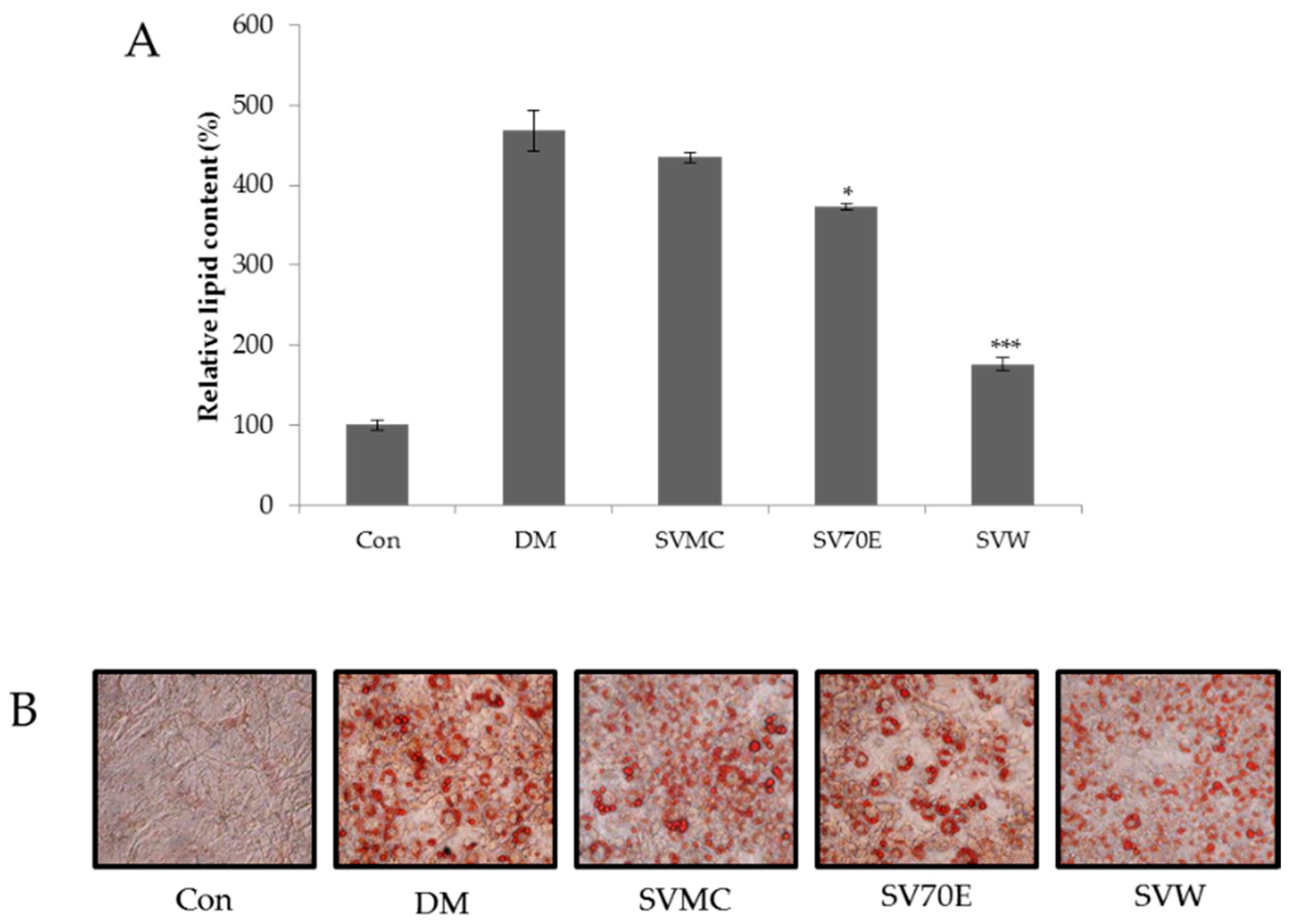
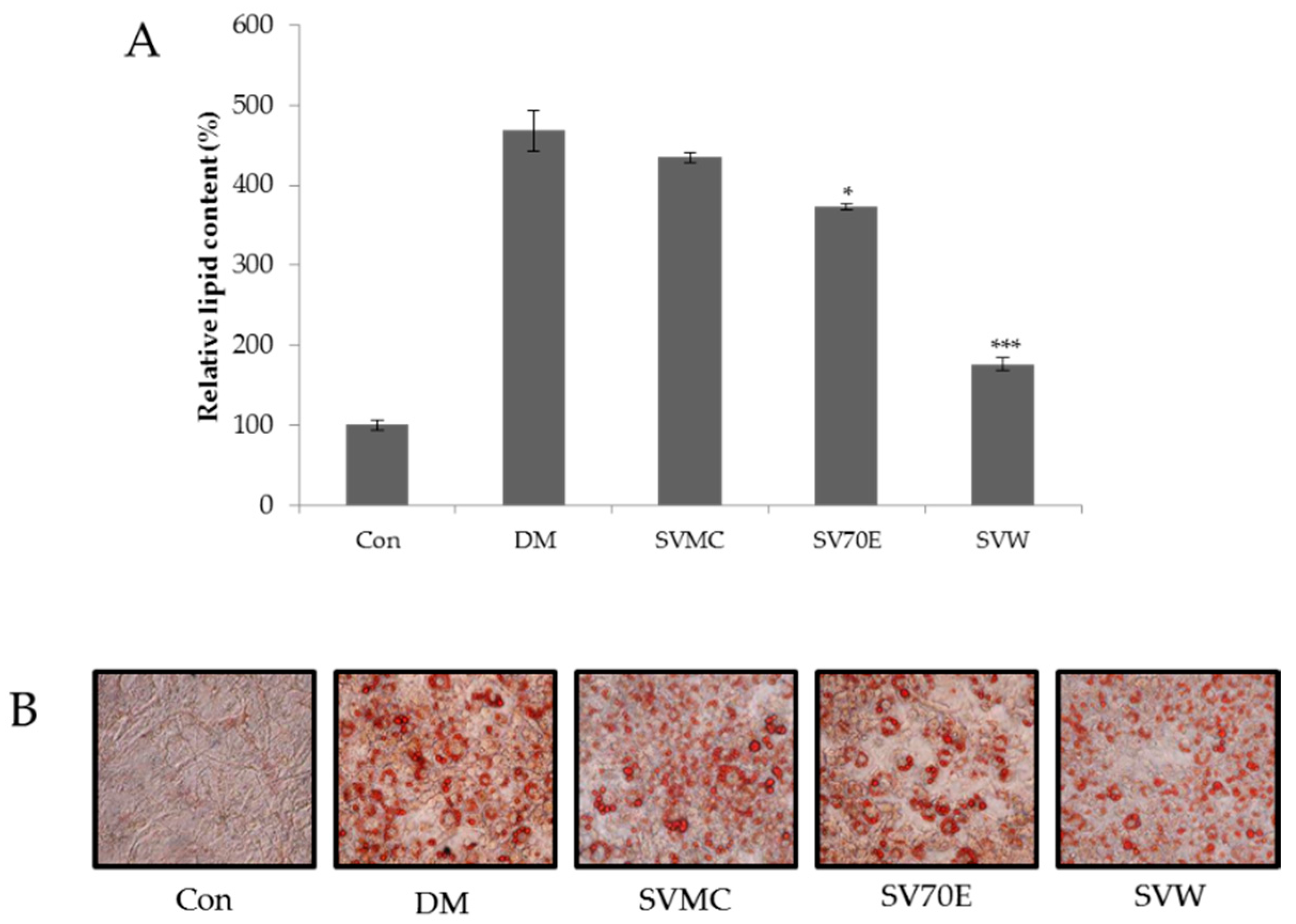
2.2. Effect of the S. virgaurea var. gigantea Extracts on Inhibition of 3T3-L1 Adipocyte Differentiation
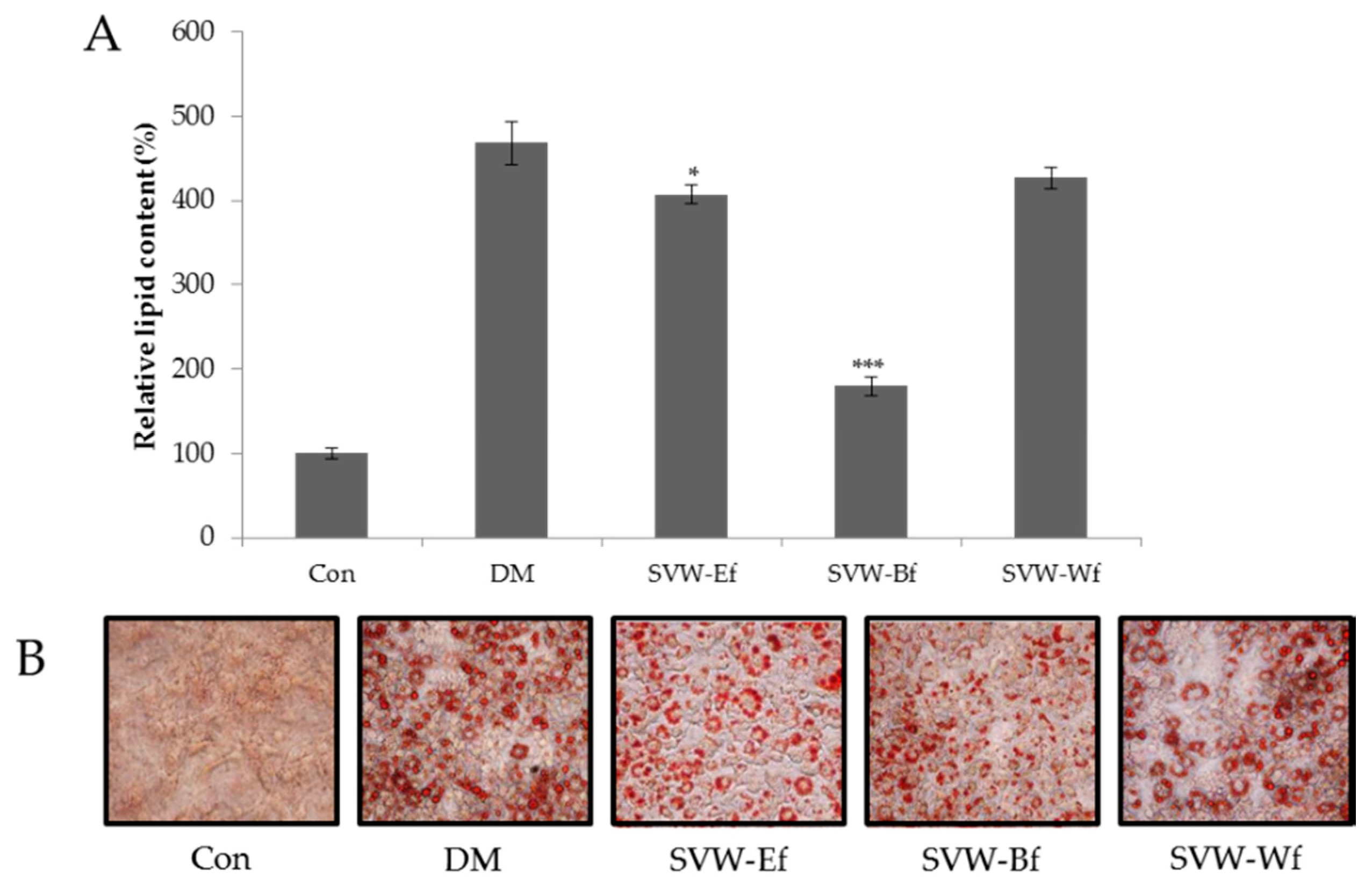
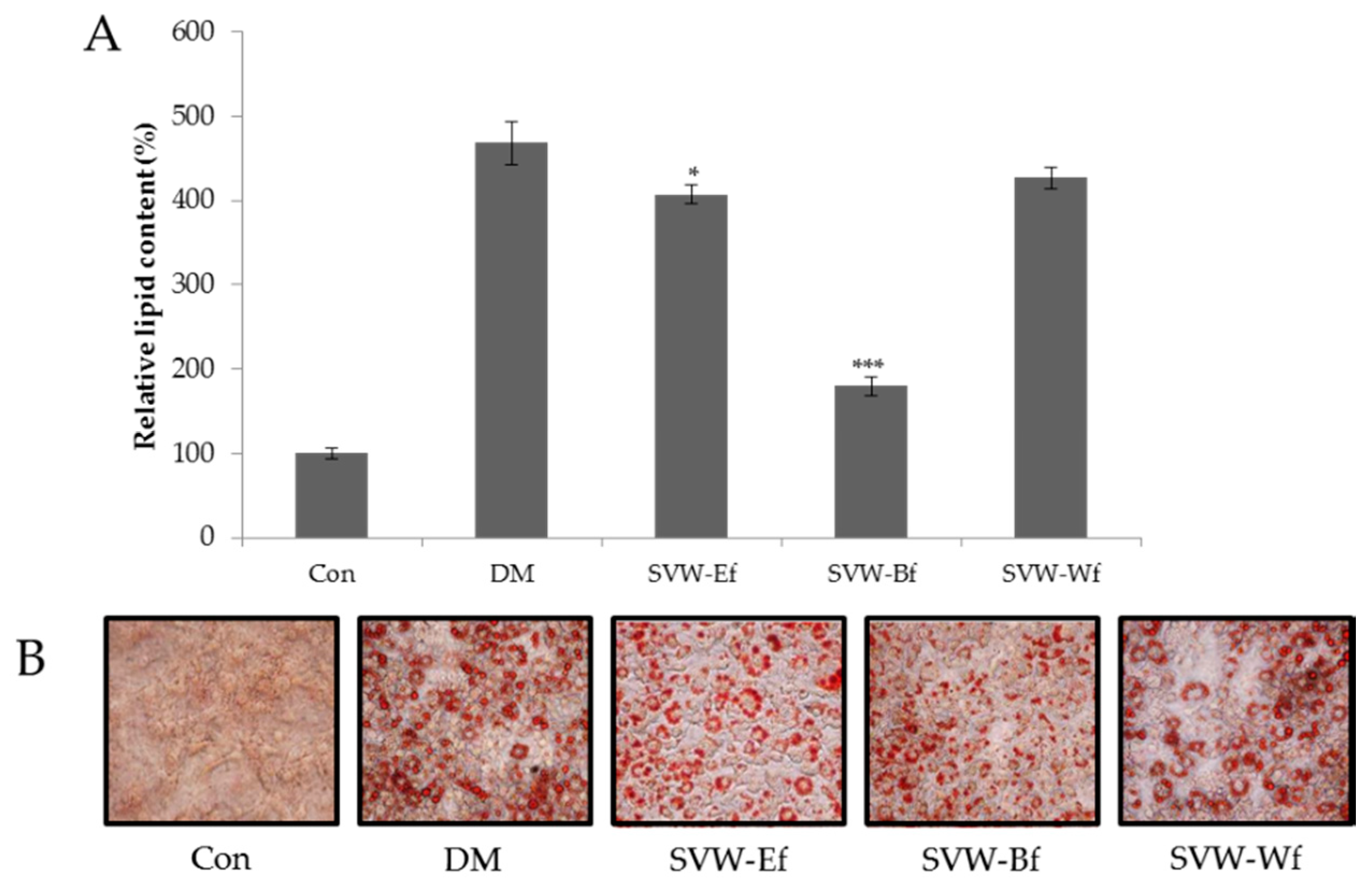
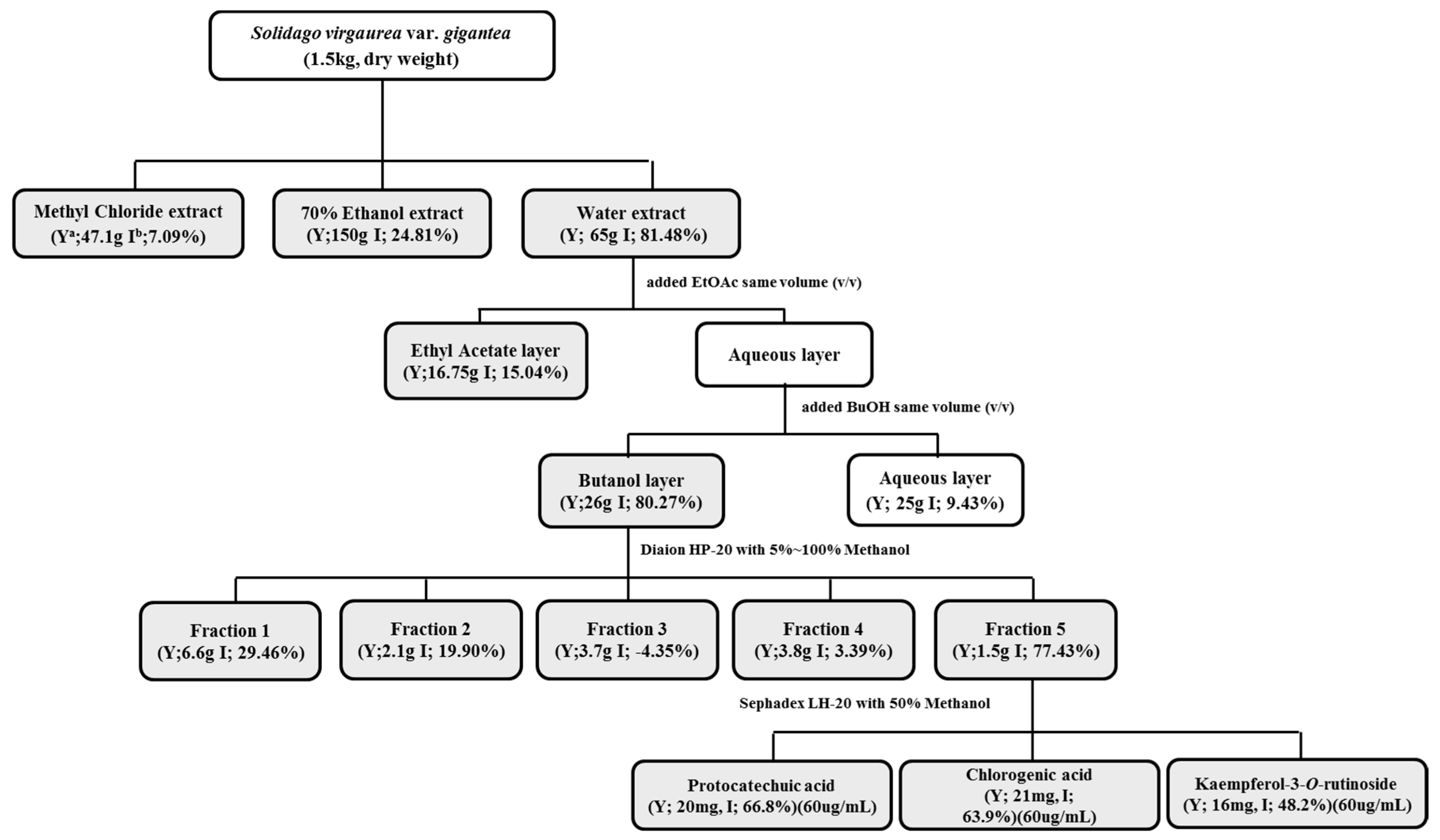
2.3. Effect of Solvent Fractions of SVW on Inhibition of 3T3-L1 Adipocyte Differentiation
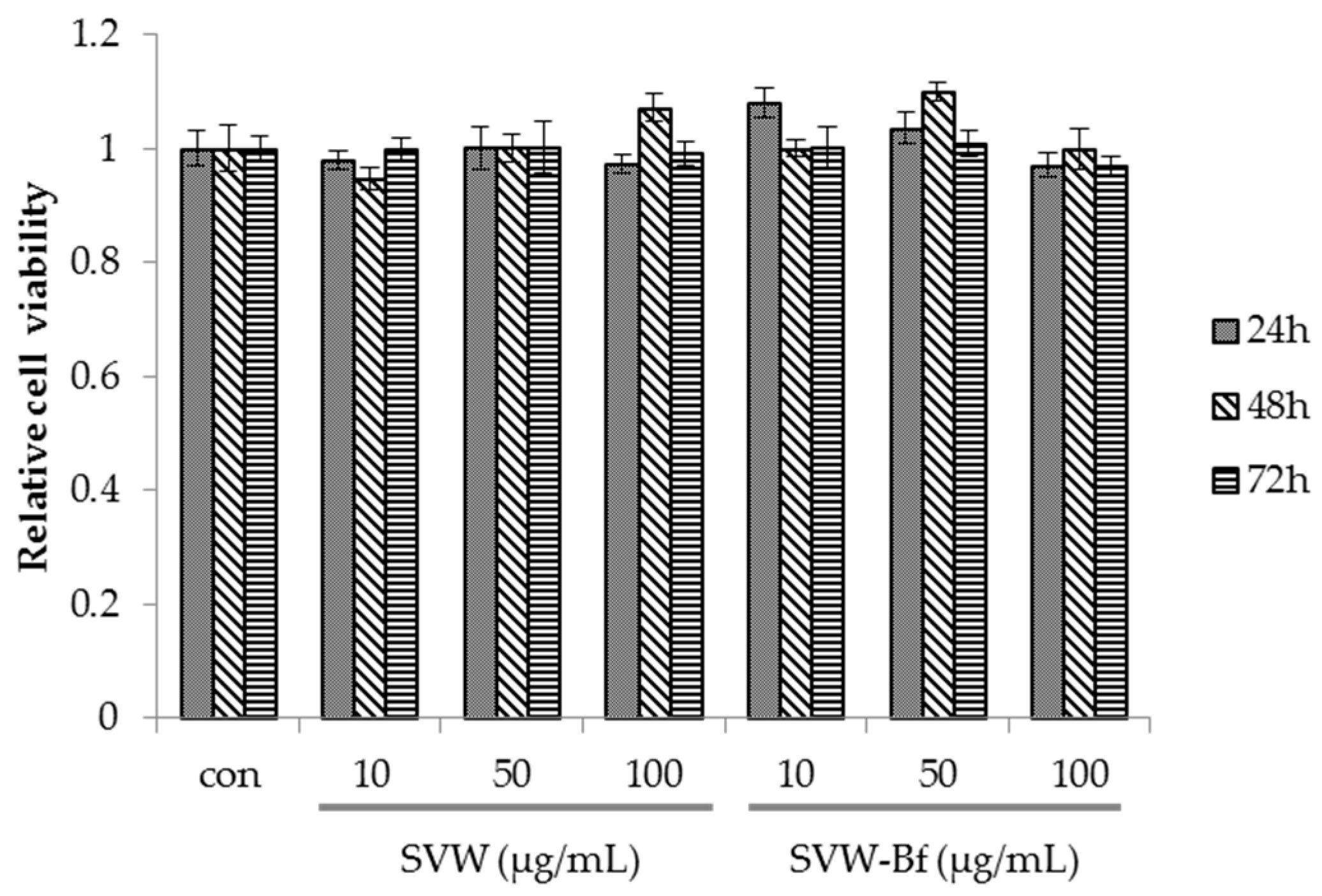
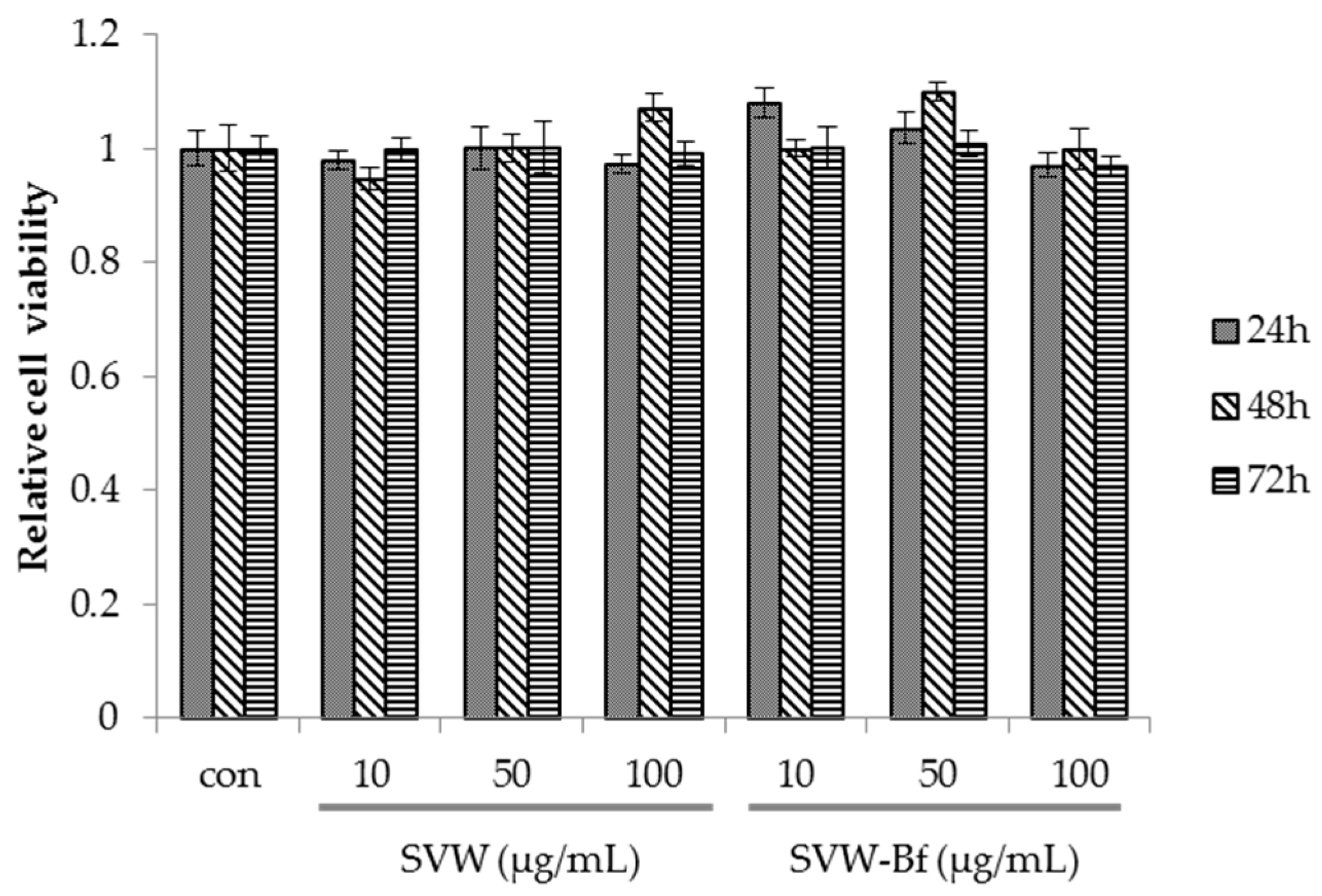
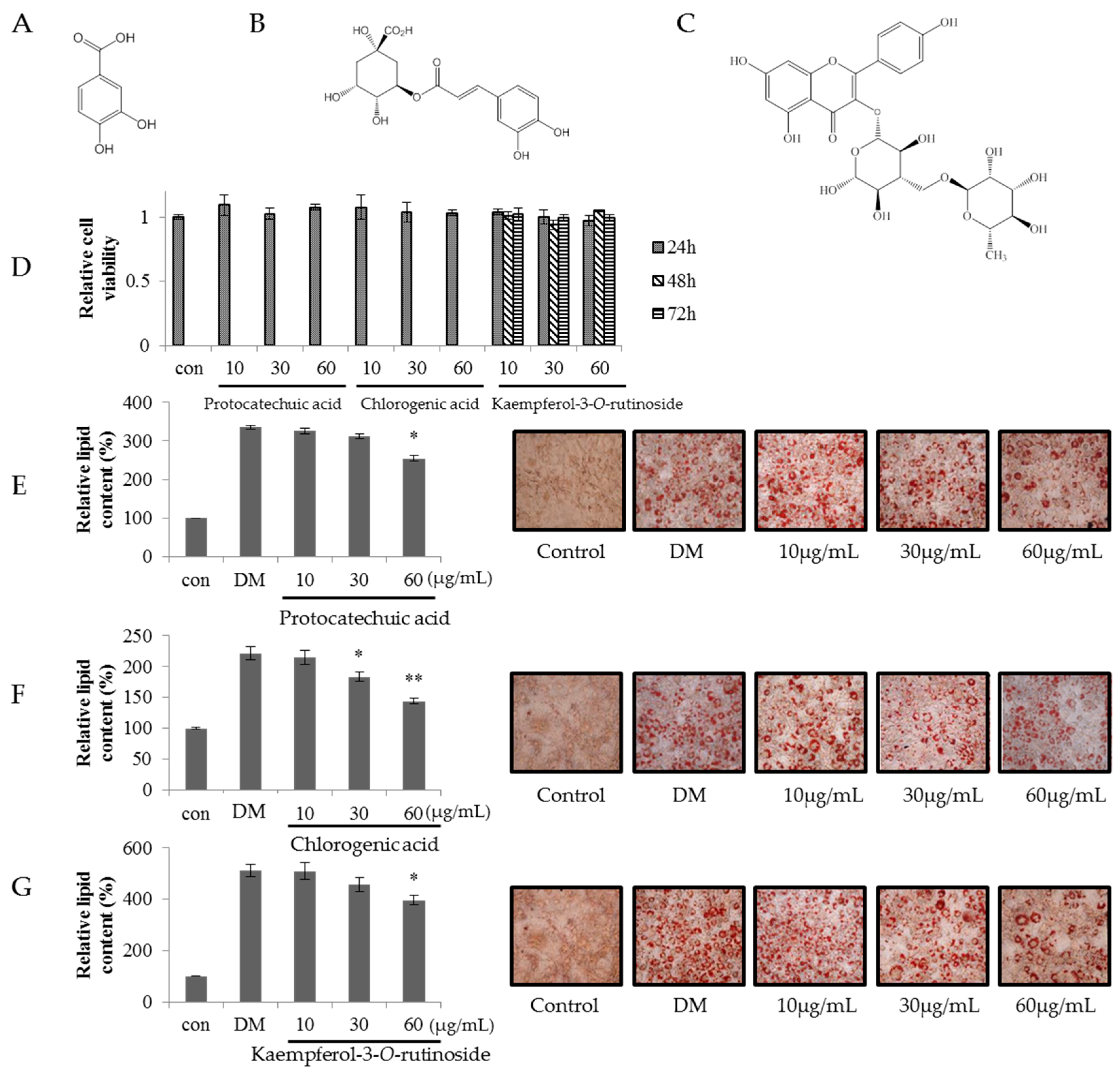
2.4. Effect of SVW and SVW-Bf on Preadipocyte Viability
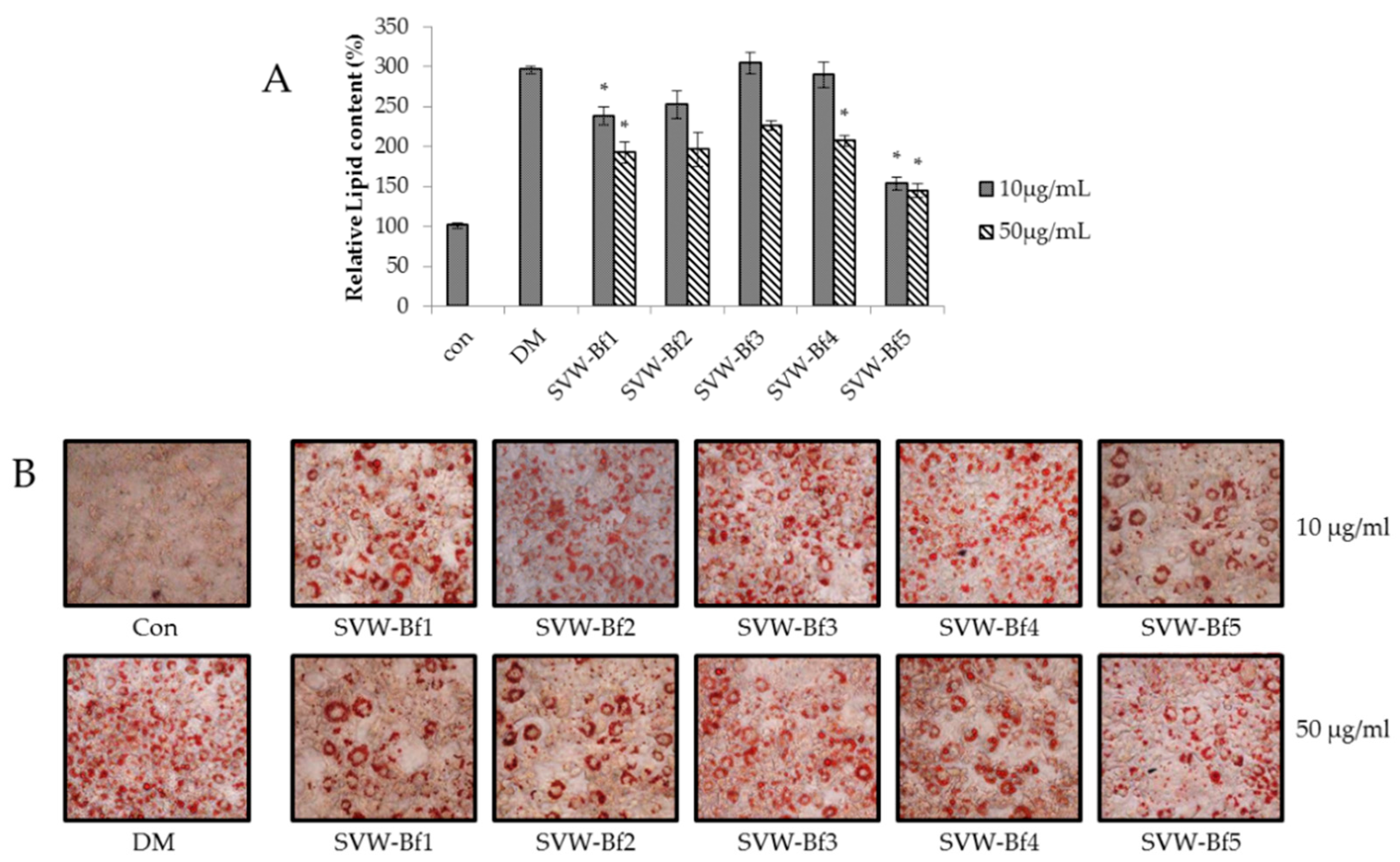
2.5. Effect of Sub-Fractions of SVW-Bf on Inhibition of 3T3-L1 Adipocyte Differentiation
2.6. Effect of Kaempferol-3-O-rutinoside, Chlorogenic Acid and Protocatechuic Acid of SVW-Bf5 on Preadipocyte Viability and Inhibition of 3T3-L1 Adipocyte Differentiation
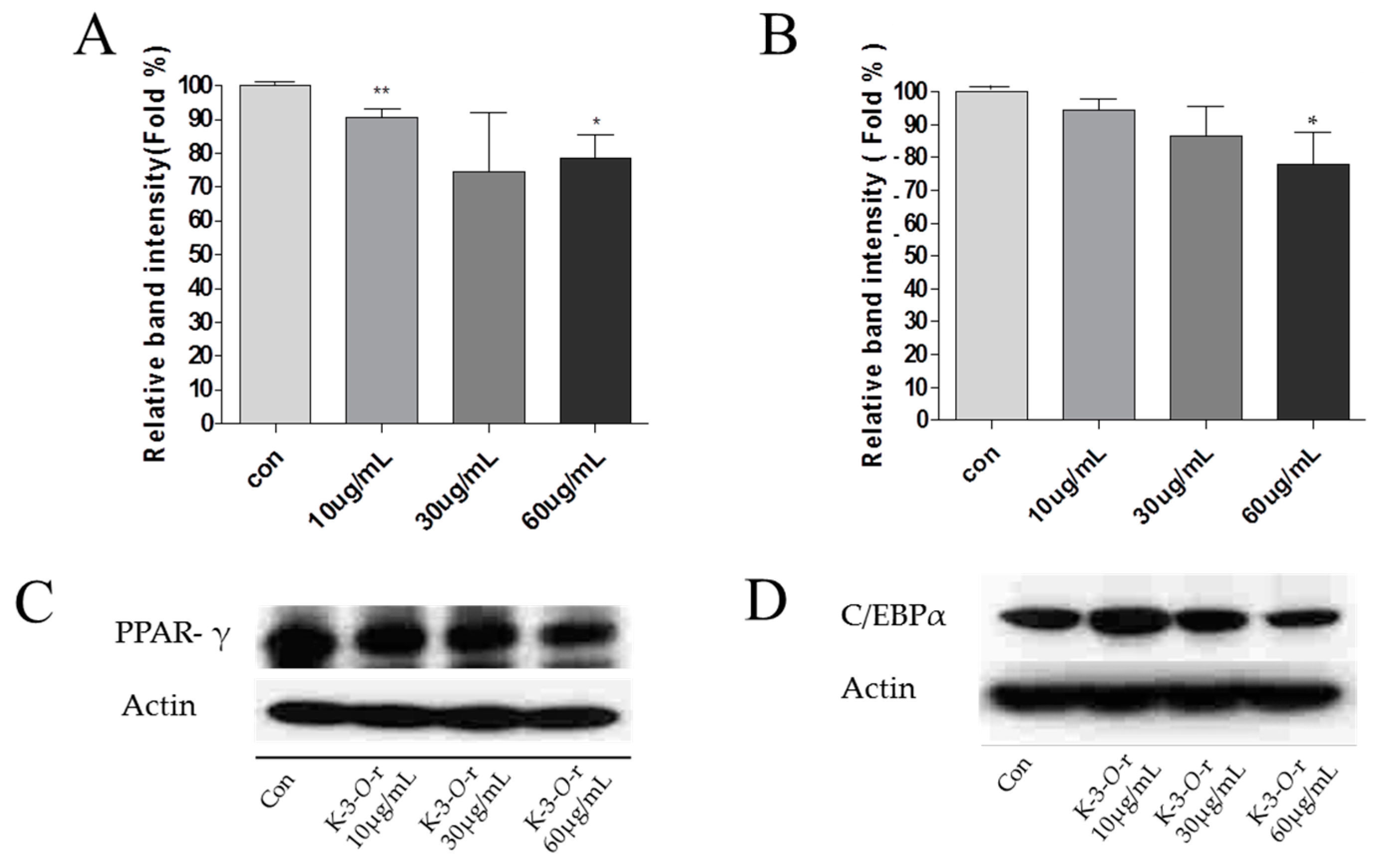
2.7. Effect of Kaempferol-3-O-rutinoside on PPAR-γ and C/EBP-α Protein Expression in 3T3-L1 Cells
3. Discussion
4. Materials and Methods
4.1. General Information
4.2. Plant Materials
4.3. Preparation of Natural Extracts
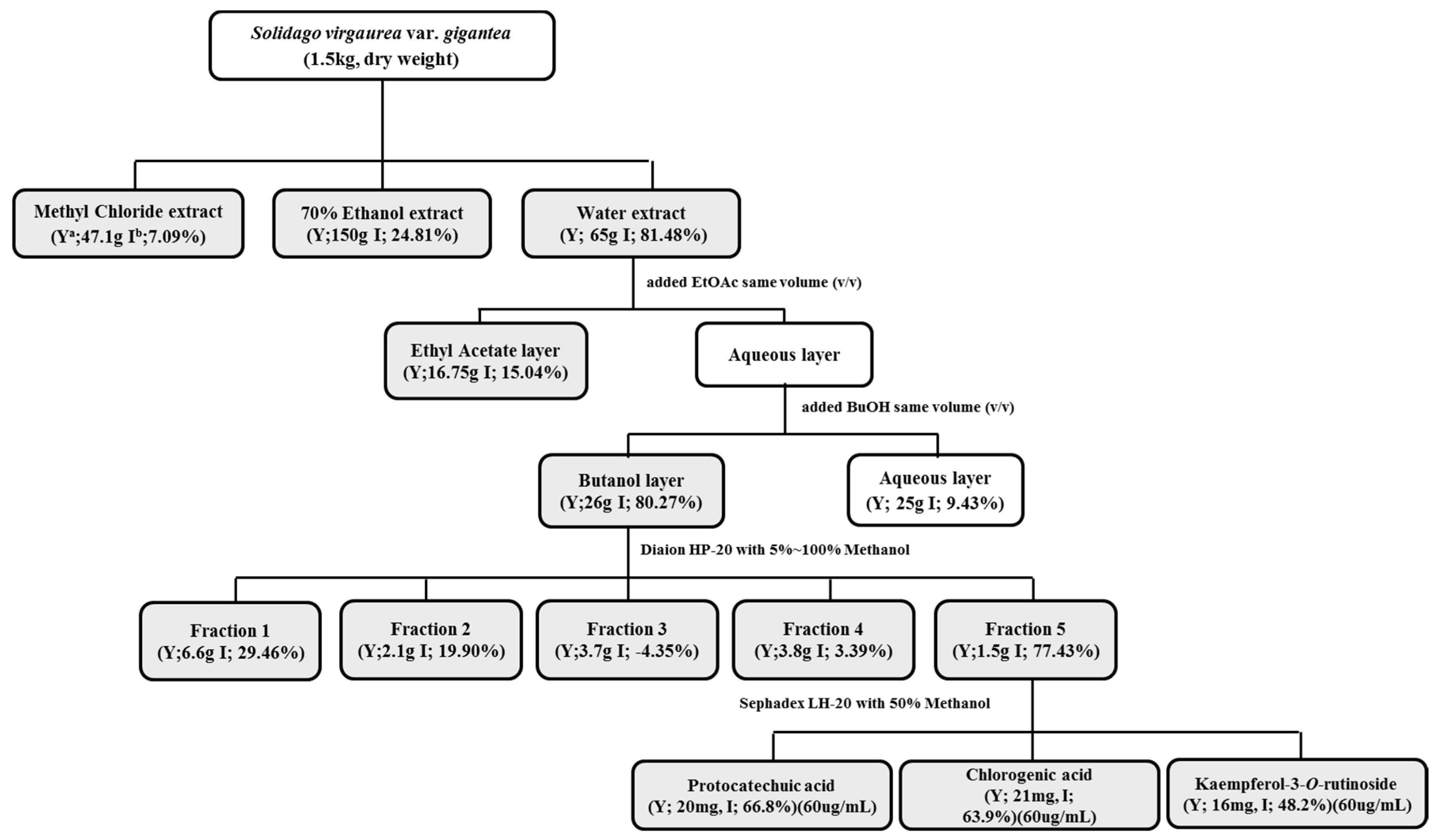
4.4. Preparation of S. virgaurea var. gigantea
4.5. Cell Culture and Differentiation
4.6. Oil Red O Staining
4.7. Cell Viability Assay
4.8. Western Blot Analysis
4.9. Statistical Analysis
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Kopelman, P.G. Obesity as a medical problem. Nature 2000, 404, 635–643. [Google Scholar] [PubMed]
- Bray, G.A.; Tartaglia, L.A. Medicinal strategies in the treatment of obesity. Nature 2000, 404, 672–677. [Google Scholar] [PubMed]
- Vázquez-Vela, M.E. F.; Torres, N.; Tovar, A.R. White adipose tissue as endocrine organ and its role in obesity. Arch. Med. Res. 2008, 39, 715–728. [Google Scholar] [CrossRef] [PubMed]
- Reilly, S.M.; Lee, C.-H. PPARδ as a therapeutic target in metabolic disease. FEBS Lett. 2008, 582, 26–31. [Google Scholar] [CrossRef] [PubMed]
- Li, S.; Luo, X. Compendium of Materia Medica: (Bencao Gangmu); Foreign Languages Press: Beijing, China, 2003. [Google Scholar]
- Lee, T.B. Illustrated Flora of Korea; H.M.S.: Seoul, Korea, 1979. [Google Scholar]
- Kim, H.S. Studies on the antimicrobial and antioxidant activity of Solidago virga-aurea LI and Solidago virgaurea Linne var. asiatica Nakai. MS Thesis, Pusan National University, Pusan, Korea, 1996. [Google Scholar]
- Leuschner, J. Anti-inflammatory, spasmolytic and diuretic effects of a commercially available Solidago gigantea Herb. extract. Arzneimittelforschung 1995, 45, 165–168. [Google Scholar] [PubMed]
- Lee, J. Effect of Solidago Virga-aurea var. giagantea Mig. Root extract on the activity of osteoblastic cells and bone metabolism. MS Thesis, Graduate School of Keimyung University, Daegu, Korea, 2004. [Google Scholar]
- Watanabe, M.; Inukai, K.; Katagiri, H.; Awata, T.; Oka, Y.; Katayama, S. Regulation of PPARγ transcriptional activity in 3T3-L1 adipocytes. Biochem. Biophys. Res. Commun. 2003, 300, 429–436. [Google Scholar] [CrossRef]
- Cao, Z.; Umek, R.M.; McKnight, S.L. Regulated expression of three C/EBP isoforms during adipose conversion of 3T3-L1 cells. Genes Dev. 1991, 5, 1538–1552. [Google Scholar] [CrossRef] [PubMed]
- Tarantino, G. Should nonalcoholic fatty liver disease be regarded as a hepatic illness only? World J. Gastroenterol. 2007, 13, 4669–4672. [Google Scholar] [CrossRef] [PubMed]
- Yang, J.Y.; Della-Fera, M.A.; Rayalam, S.; Ambati, S.; Hartzell, D.L.; Park, H.J.; Baile, C.A. Enhanced inhibition of adipogenesis and induction of apoptosis in 3T3-L1 adipocytes with combinations of resveratrol and quercetin. Life Sci. 2008, 82, 1032–1039. [Google Scholar] [CrossRef] [PubMed]
- Rosen, E.D.; Spiegelman, B.M. Adipocytes as regulators of energy balance and glucose homeostasis. Nature 2006, 444, 847–853. [Google Scholar] [CrossRef] [PubMed]
- Roncari, D.A.K.; Lau, D.C.W.; Kindler, S. Exaggerated replication in culture of adipocyte precursors from massively obese persons. Metabolism 1981, 30, 425–427. [Google Scholar] [CrossRef]
- Finelli, C.; Tarantino, G. Is there any consensus as to what diet or lifestyle approach is the right one for NAFLD patients? J. Gastrointestin. Liver Dis. 2012, 21, 293–302. [Google Scholar] [PubMed]
- Seo, J.B.; Choe, S.S.; Jeong, H.W.; Park, S.W.; Shin, H.J.; Choi, S.M.; Park, J.Y.; Choi, E.W.; Kim, J.B.; Seen, D.S.; et al. Anti-obesity effects of Lysimachia feonum-graecum characterized by decreased adipogenesis and regulated lipid metabolism. Exp. Mol. Med. 2011, 43, 205–215. [Google Scholar] [CrossRef] [PubMed]
- Kong, C.S.; Kim, J.A.; Kim, S.K. Anti-obesity effect of sulfated glucosamine by AMPK signal pathway in 3T3-L1 adipocytes. Food Chem. Toxicol. 2009, 47, 2401–2406. [Google Scholar] [CrossRef] [PubMed]
- Kwon, C.-S.; Sohn, H.Y.; Kim, S.H.; Kim, J.H.; Son, K.H.; Lee, J.S.; Lim, J.K.; Kim, J.-S. Anti-obesity effect of Dioscorea nipponica Makino with lipase-inhibitory activity in rodents. Biosci. Biotechnol. Biochem. 2003, 67, 1451–1456. [Google Scholar] [CrossRef] [PubMed]
- Lahrita, L.; Kato, E.; Kawabata, J. Uncovering potential of Indonesian medicinal plants on glucose uptake enhancement and lipid suppression in 3T3-L1 adipocytes. J. Ethnopharmacol. 2015, 168, 229–236. [Google Scholar] [CrossRef] [PubMed]
- Demir, H.; Açik, L.; Bali, E.B.; Koç, L.Y.; Kaynak, G. Antioxidant and antimicrobial activities of Solidago virgaurea extracts. Afr. J. Biotechnol. 2009, 8, 274–279. [Google Scholar]
- El-Tantawy, W.H. Biochemical effects of Solidago virgaurea extract on experimental cardiotoxicity. J. Physiol. Biochem. 2014, 70, 33–42. [Google Scholar] [CrossRef] [PubMed]
- Jafari, S.; Saeidnia, S.; Hajimehdipoor, H.; Ardekani, M.R.S.; Faramarzi, M.A.; Hadjiakhoondi, A.; Khanavi, M. Cytotoxic evaluation of Melia azedarach in comparison with, Azadirachta indica and its phytochemical investigation. DARU 2013, 21. [Google Scholar] [CrossRef] [PubMed]
- Christensen, K.B.; Petersen, R.K.; Kristiansen, K.; Christensen, L.P. Identification of bioactive compounds from flowers of black elder (Sambucus nigra L.) that active the human peroxisome proliferator-active receptor (PPAR) γ. Phytother. Res. 2010, 24, S129–S132. [Google Scholar] [CrossRef] [PubMed]
- Scazzocchio, B.; Varì, R.; Filesi, C.; D’Archivio, M.; Santangelo, C.; Giovannini, C.; Iacovelli, A.; Silecchia, G.; Volti, G.L.; Galvano, F.; et al. Cyanidin-3-O-β-glucoside and protocatechuic acid exert insulin-like effects by upregulating PPARγ activity in human omental adipocytes. Diabetes 2011, 60, 2234–2244. [Google Scholar] [CrossRef] [PubMed]
- Habtemariam, S. α-Glucosidase inhibitory activity of kaempferol-3-O-rutinoside. Nat. Prod. Commun. 2011, 6, 201–203. [Google Scholar] [PubMed]
- Calderón-Montaño, J.M.; Burgos-Morón, E.; Pérez-Guerrero, C.; López-Lázaro, M. A review on the dietary flavonoid kaempferol. Mini-Rev. Med. Chem. 2011, 11, 298–344. [Google Scholar] [CrossRef] [PubMed]
- Lee, Y.J.; Choi, H.S.; Seo, M.J.; Jeon, H.J.; Kim, K.J.; Lee, B.Y. Kaempferol suppresses lipid accumulation by inhibiting early adipogenesis in 3T3-L1 cells and zebrafish. Food Funct. 2015, 6, 2824–2833. [Google Scholar] [CrossRef] [PubMed]
- Gregoire, F.M.; Smas, C.M.; Sul, H.S. Understanding adipocyte differentiation. Physiol. Rev. 1998, 78, 783–809. [Google Scholar] [PubMed]
- Finelli, C.; Tarantino, G. What is the role of adiponectin in obesity related non-alcoholic fatty liver disease? World J. Gastroenterol. 2013, 19, 802–812. [Google Scholar] [CrossRef] [PubMed]
- Takemura, T.; Takatsu, Y.; Kasumi, M.; Marubashi, W.; Iwashina, T. Flavonoids and their distribution patterns in the flowers of Gladiolus cultivars. Acta Hortic. 2005, 673, 487–493. [Google Scholar] [CrossRef]
- Leiss, K.A.; Maltese, F.; Choi, Y.H.; Verpoorte, R.; Klinkhamer, P.G.L. Identification of chlorogeneic acid as a resistance factor for thrips in chrysanthemum. Plant Physiol. 2009, 150, 1567–1575. [Google Scholar] [CrossRef] [PubMed]
- Jung, H.A.; Park, J.C.; Chung, H.Y.; Kim, J.; Choi, J.S. Antioxidant flavonoids and chlorogenic acid from the leaves of Eriobotrya japonica. Arch. Pham. Res. 1999, 22, 213–218. [Google Scholar] [CrossRef]
- Lee, J.C.; Lee, K.Y.; Na, C.S.; Jung, N.C.; Chung, G.H.; Jang, Y.S. Extract from Rhus verniciflua Stokes is capable of inhibiting the growth of human lymphoma cells. Food Chem. Toxicol. 2004, 42, 1383–1388. [Google Scholar] [CrossRef] [PubMed]
- Sample Availability: Samples of the compounds kaempferol-3-O-rutinoside, chlorogenic acid, and protocatechuic acid are available from the authors.





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
© 2016 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons by Attribution (CC-BY) license ( http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jang, Y.S.; Wang, Z.; Lee, J.-M.; Lee, J.-Y.; Lim, S.S. Screening of Korean Natural Products for Anti-Adipogenesis Properties and Isolation of Kaempferol-3-O-rutinoside as a Potent Anti-Adipogenetic Compound from Solidago virgaurea. Molecules 2016, 21, 226. https://doi.org/10.3390/molecules21020226
Jang YS, Wang Z, Lee J-M, Lee J-Y, Lim SS. Screening of Korean Natural Products for Anti-Adipogenesis Properties and Isolation of Kaempferol-3-O-rutinoside as a Potent Anti-Adipogenetic Compound from Solidago virgaurea. Molecules. 2016; 21(2):226. https://doi.org/10.3390/molecules21020226
Chicago/Turabian StyleJang, Young Soo, Zhiqiang Wang, Jeong-Min Lee, Jae-Young Lee, and Soon Sung Lim. 2016. "Screening of Korean Natural Products for Anti-Adipogenesis Properties and Isolation of Kaempferol-3-O-rutinoside as a Potent Anti-Adipogenetic Compound from Solidago virgaurea" Molecules 21, no. 2: 226. https://doi.org/10.3390/molecules21020226