
Effects of Light Intensity on the Growth, Photosynthetic Characteristics, and Flavonoid Content of Epimedium pseudowushanense B.L.Guo


Abstract
:1. Introduction
2. Results
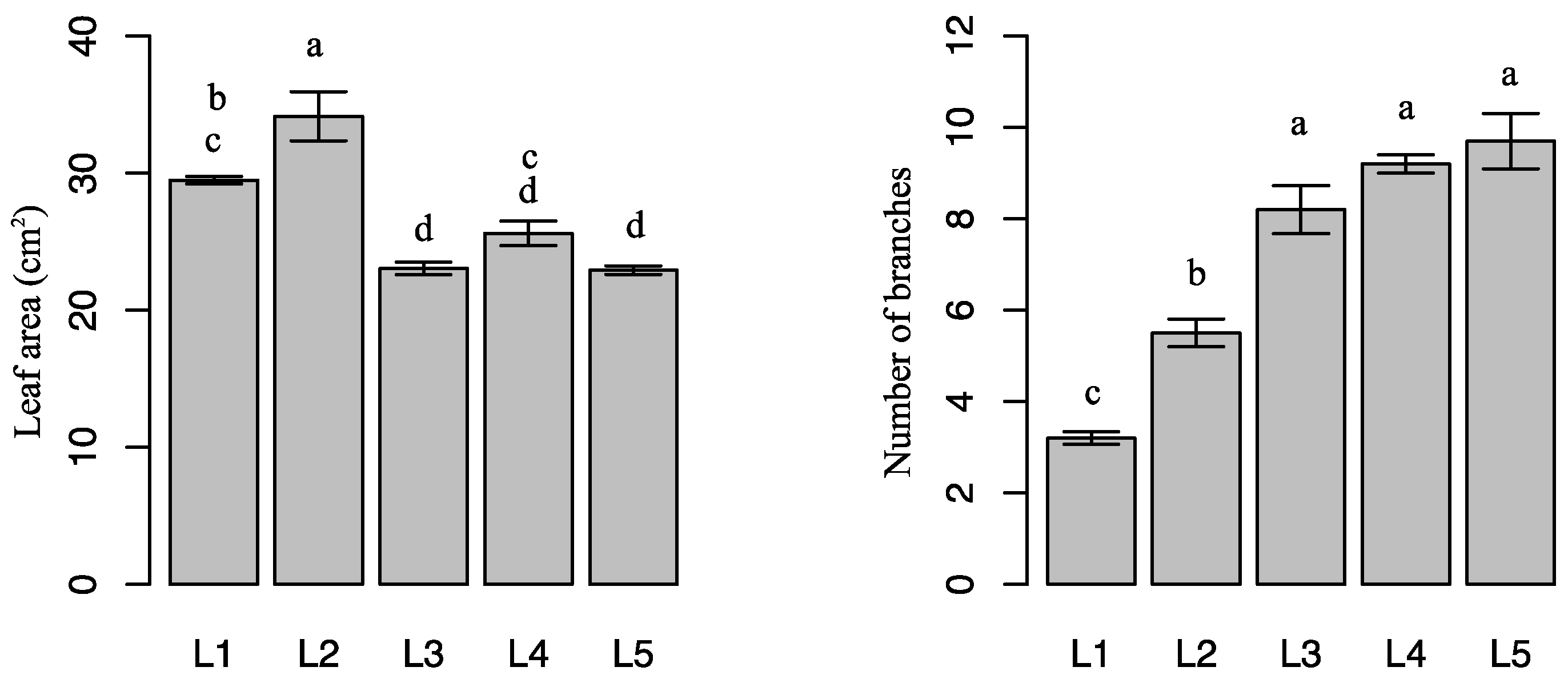
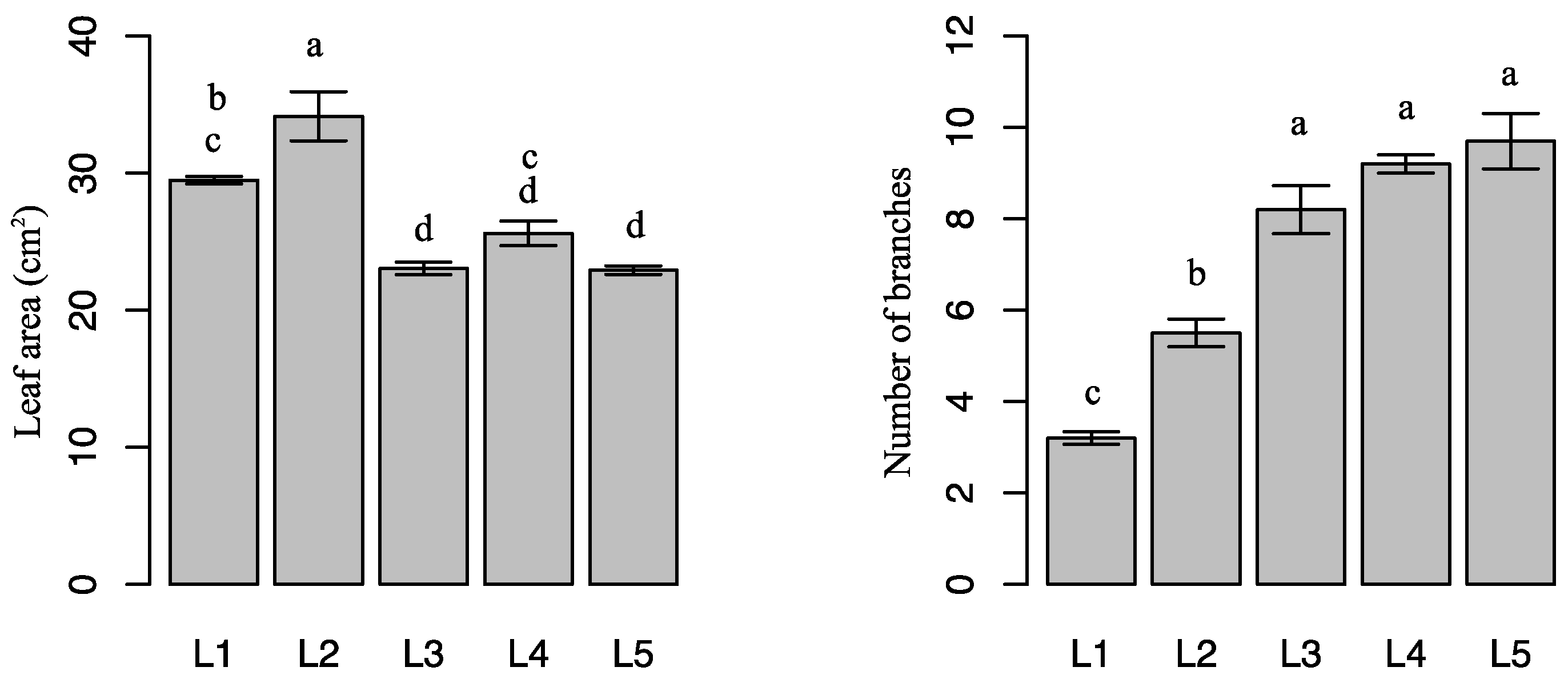
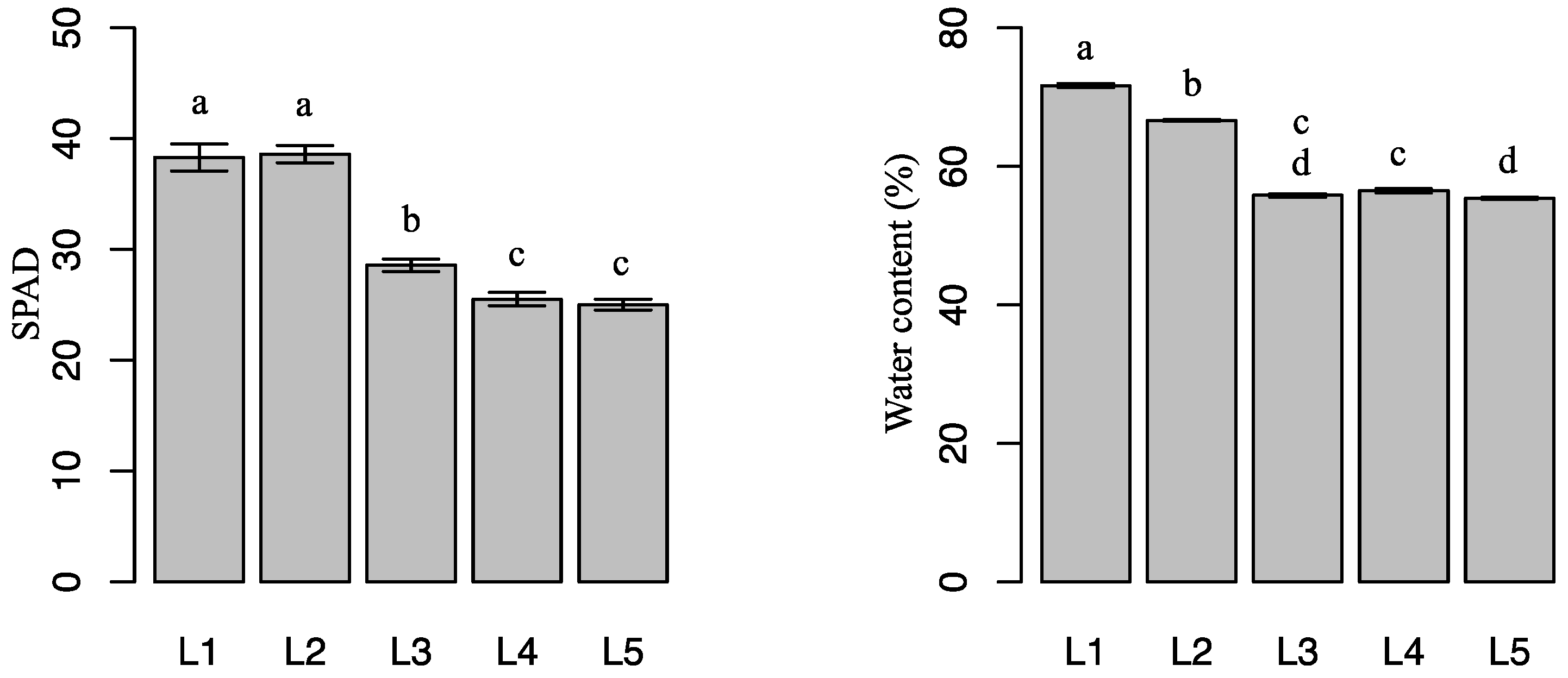
2.1. Effects of Light Intensity on Plant Growth
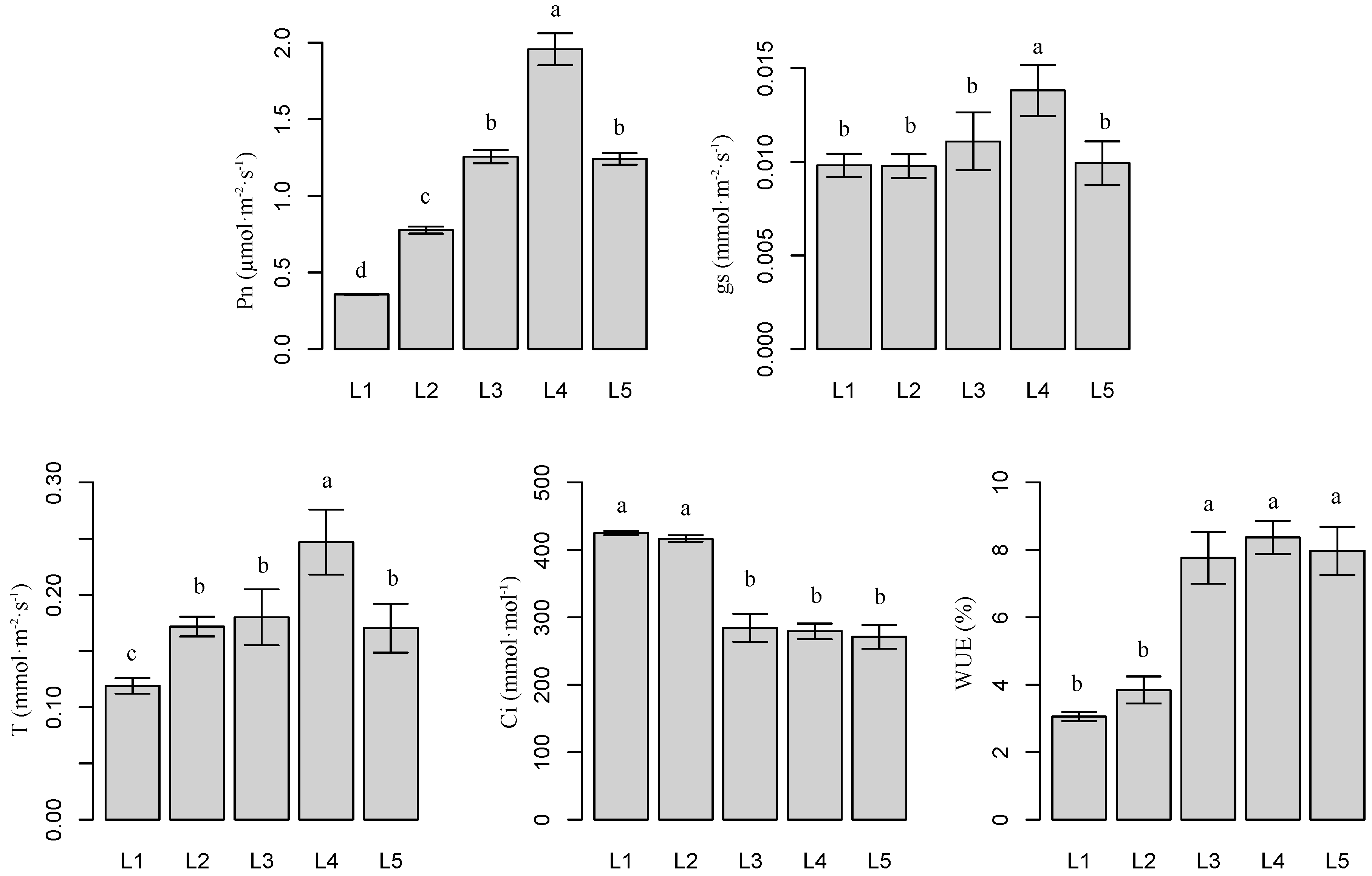
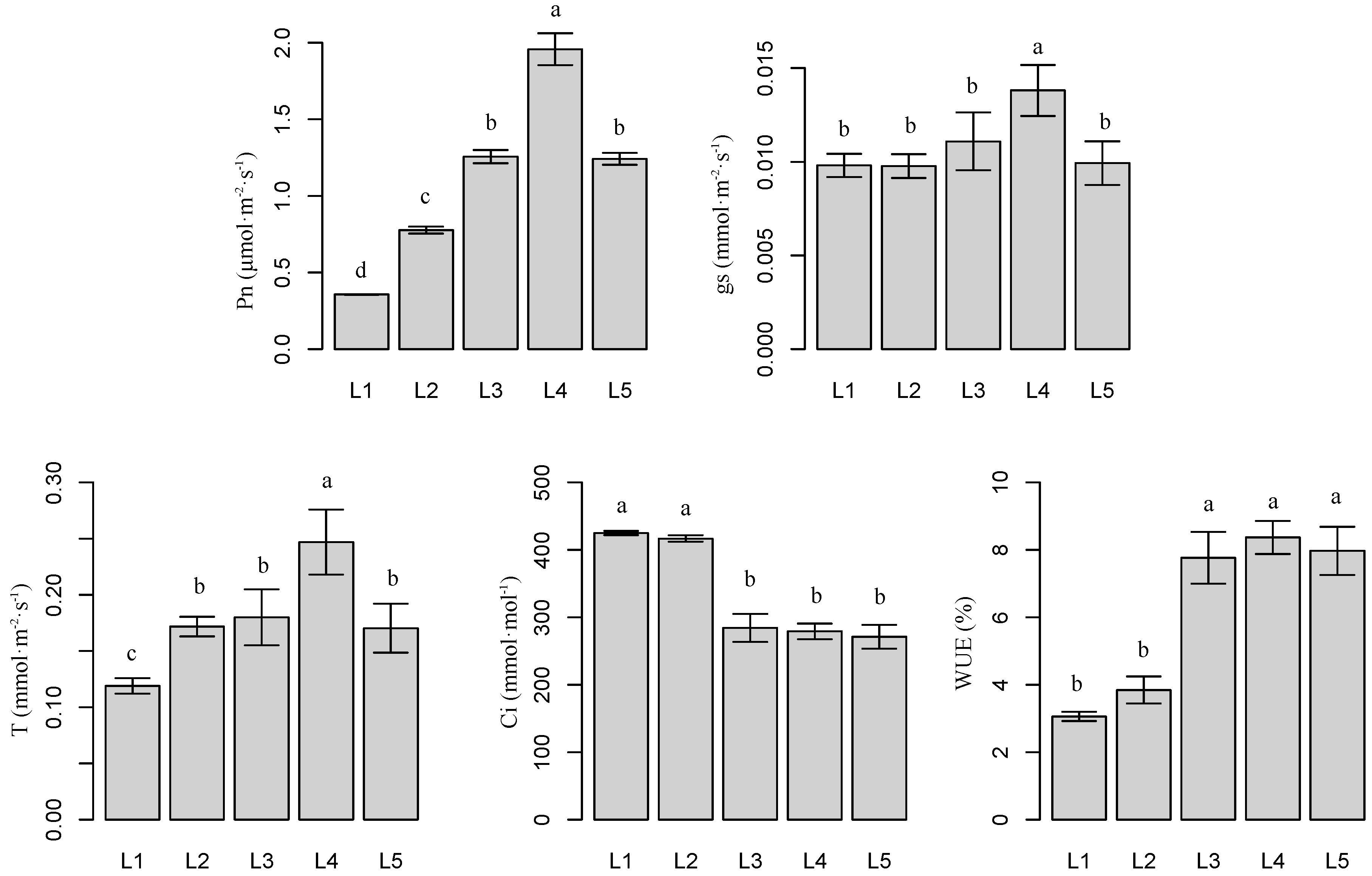
2.2. Effects of Light Intensity on Plant Photosynthetic Parameters
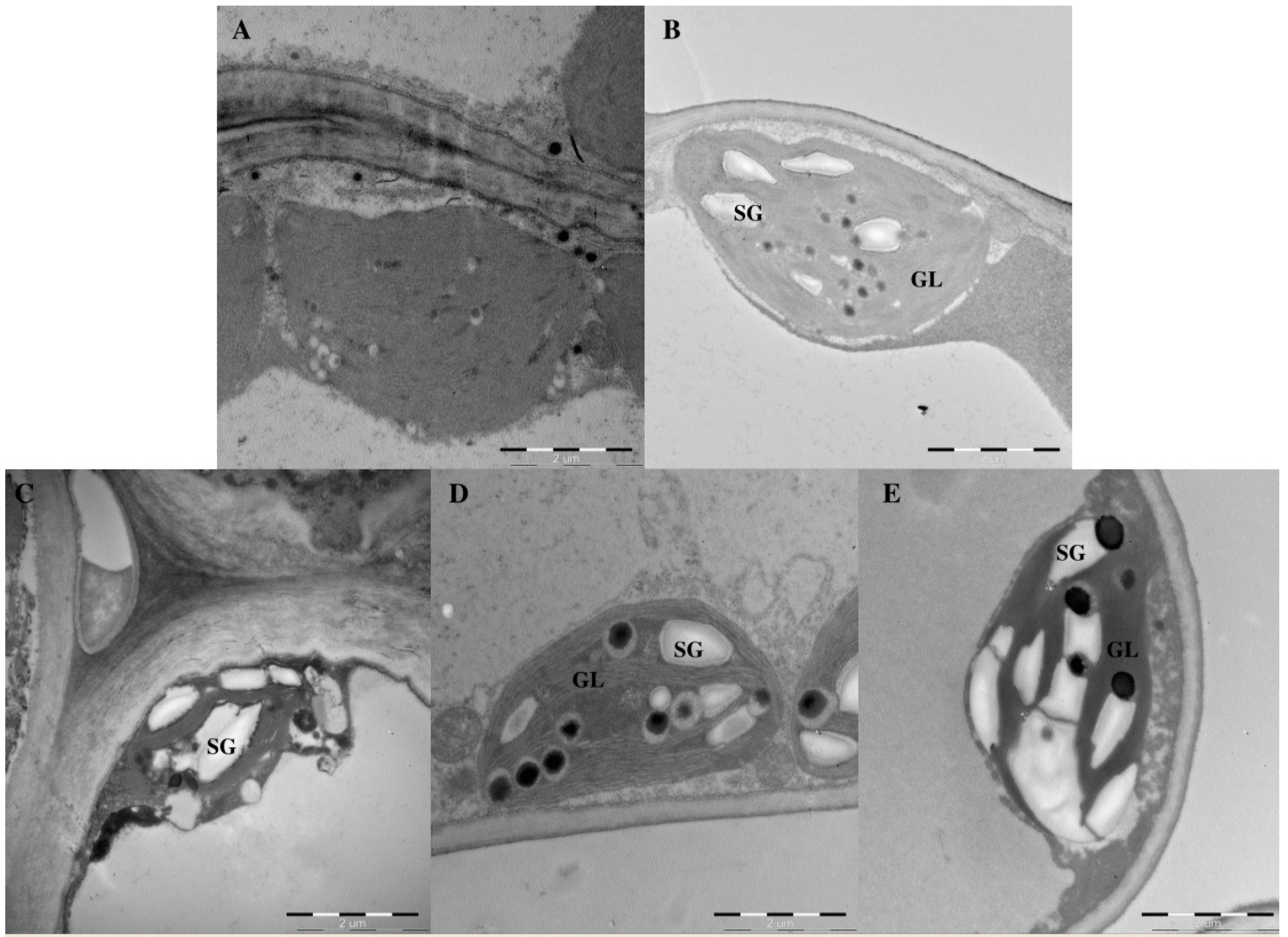
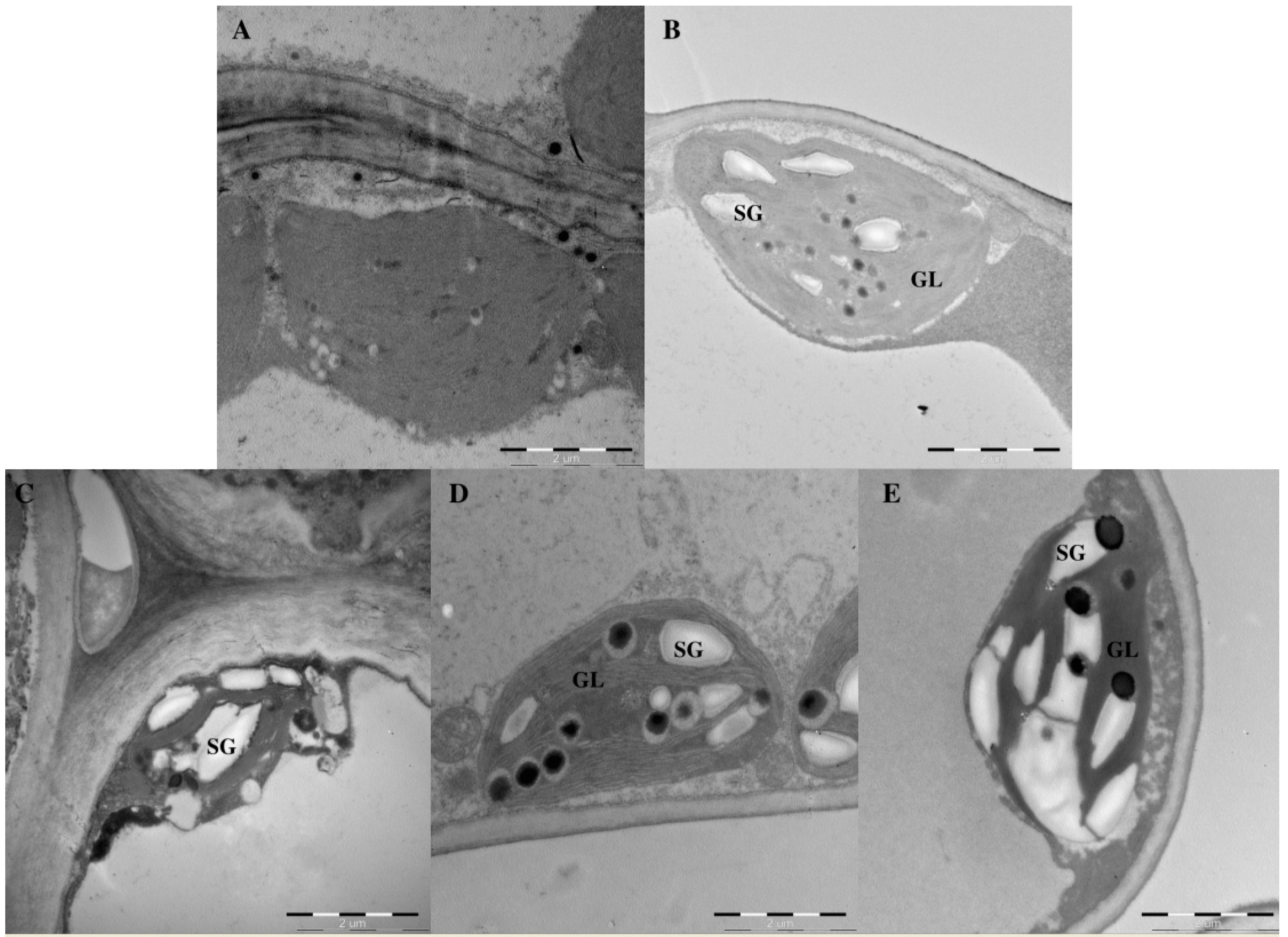
2.3. Effects of Light Intensity on Chloroplast Ultrastructure
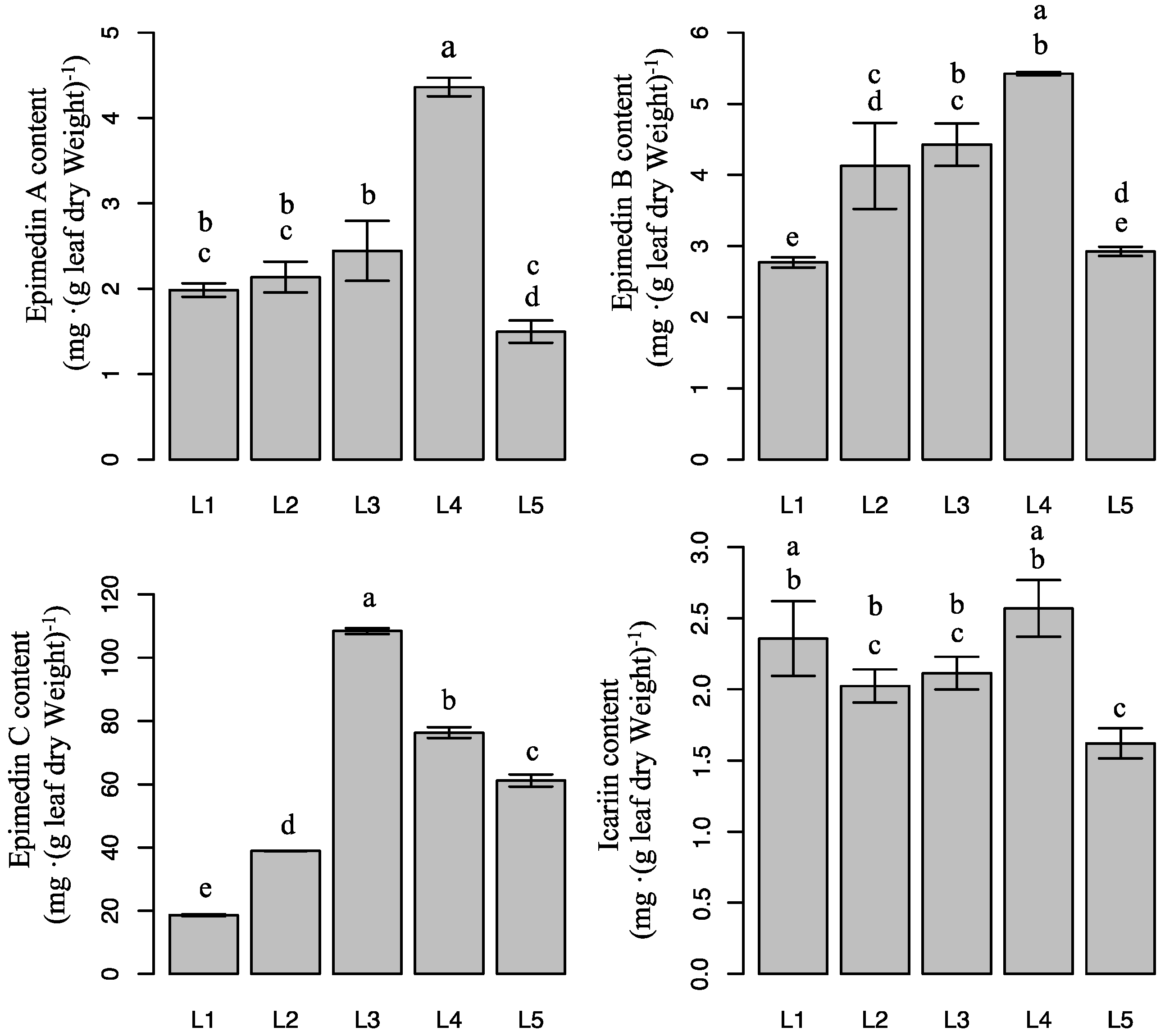
2.4. Effect of Light Intensity on Flavonoid Content in Epimedium
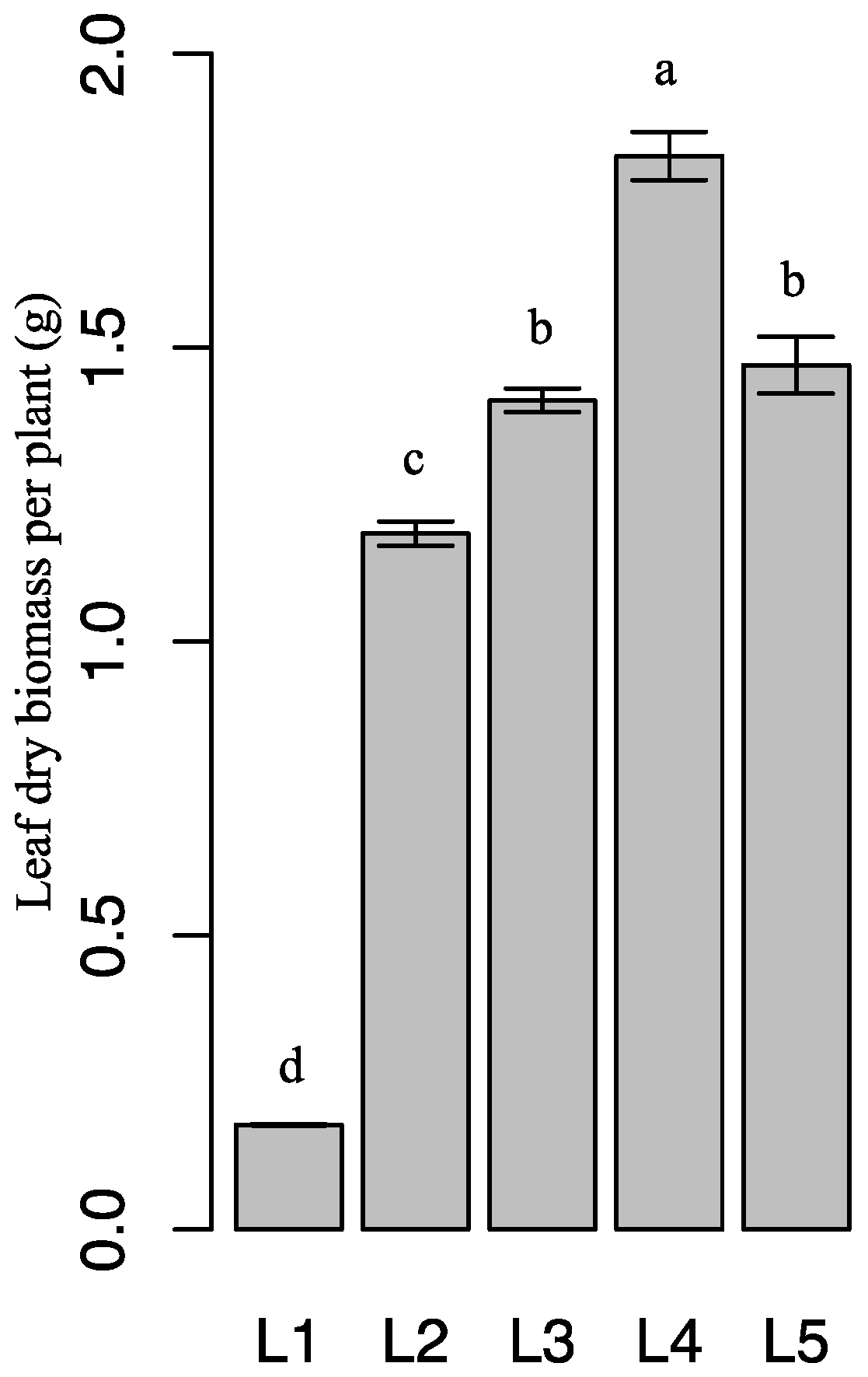
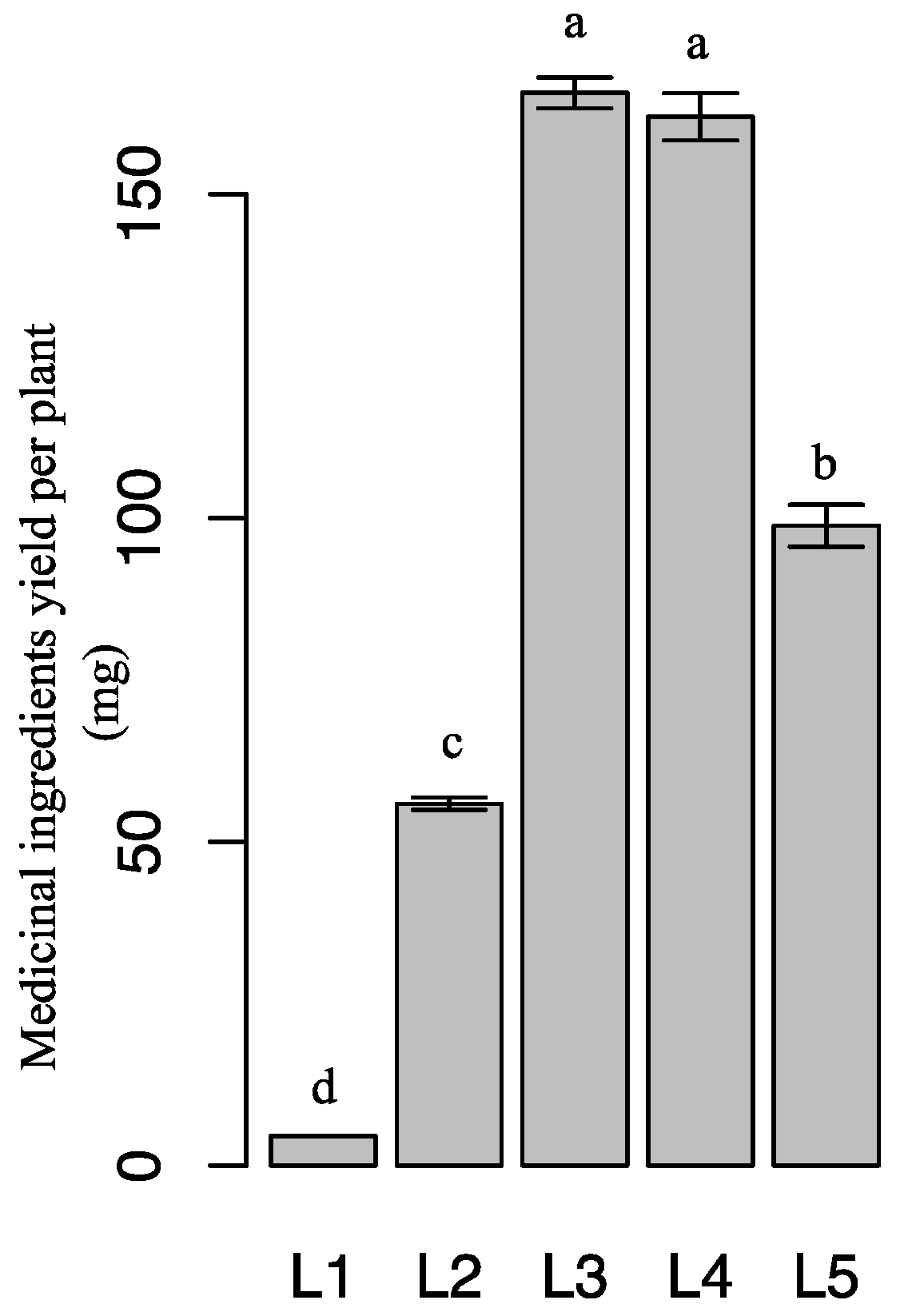
2.5. Effect of Light Intensity on Leaf Dry Biomass and Medicinal-Ingredient Yield of Epimedium
3. Discussion
4. Materials and Methods
4.1. Plant Materials and Growth Conditions
4.2. Plant Growth, Chlorophyll Content, and Leaf Dry Biomass
4.3. Photosynthetic Parameters
4.4. Assessment of Chloroplast Ultrastructure
4.5. Flavonoid Content and Medicinal-Ingredient Yield
4.6. Data Analysis
Acknowledgments
Author Contributions
Conflicts of Interest
Abbreviations
| Pn | Net photosynthetic rate |
| gs | Stomatal conductance |
| SPAD | Soil and Plant Analyzer Development (the relative content of chlorophyll) |
| T | Transpiration rate |
| WUE | Water use efficiency |
| Ci | Intercellular CO2 concentration |
| SG | Starch grains |
| GL | Grana lamellae |
| VPD | Vapor pressure deficit |
References
- Wu, H.; Lien, E.J.; Lien, L.L. Chemical and pharmacological investigations of Epimedium species: A survey. In Progress in Drug Research; Jucker, E., Ed.; Birkhäuser Basel: Basel, Switzerland, 2003; pp. 1–57. [Google Scholar]
- Liu, J.J.; Li, S.P.; Wang, Y.T. Optimization for quantitative determination of four flavonoids in Epimedium by capillary zone electrophoresis coupled with diode array detection using central composite design. J. Chromatogr. A 2006, 1103, 344–349. [Google Scholar] [CrossRef] [PubMed]
- Kim, B.; Park, B. Baohuoside I suppresses invasion of cervical and breast cancer cells through the downregulation of CXCR4 chemokine receptor expression. Biochemistry 2014, 53, 7562–7569. [Google Scholar] [CrossRef] [PubMed]
- Lin, C.C.; Ng, L.T.; Hsu, F.F.; Shieh, D.-E.; Chiang, L.C. Cytotoxic effects of coptis chinensis and Epimedium sagittatum extracts and their major constituents (berberine, coptisine and icariin) on hepatoma and leukaemia cell growth. Clin. Exp. Pharmacol. Physiol. 2004, 31, 65–69. [Google Scholar] [CrossRef] [PubMed]
- Kang, S.H.; Jeong, S.J.; Kim, S.H.; Kim, J.H.; Jung, J.H.; Koh, W.; Kim, J.H.; Kim, D.K.; Chen, C.Y.; Kim, S.H. Icariside ii induces apoptosis in U937 acute myeloid leukemia cells: Role of inactivation of STAT3-related signaling. PLoS ONE 2012, 7, e28706. [Google Scholar]
- Zoratti, L.; Karppinen, K.; Luengo Escobar, A.; Häggman, H.; Jaakola, L. Light-controlled flavonoid biosynthesis in fruits. Front. Plant Sci. 2014, 5, 534. [Google Scholar] [CrossRef] [PubMed]
- Guo, Y.-P.; Guo, D.-P.; Zhou, H.-F.; Hu, M.-J.; Shen, Y.-G. Photoinhibition and xanthophyll cycle activity in bayberry (myrica rubra) leaves induced by high irradiance. Photosynthetica 2006, 44, 439–446. [Google Scholar] [CrossRef]
- Ma, X.; Song, L.; Yu, W.; Hu, Y.; Liu, Y.; Wu, J.; Ying, Y. Growth, physiological, and biochemical responses of Camptotheca acuminata seedlings to different light environments. Front. Plant Sci. 2015, 6, 321. [Google Scholar] [CrossRef] [PubMed]
- Jaakola, L.; Hohtola, A. Effect of latitude on flavonoid biosynthesis in plants. Plant Cell Environ. 2010, 33, 1239–1247. [Google Scholar] [CrossRef] [PubMed]
- Guo, J.; Han, W.; Wang, M.-H. Ultraviolet and environmental stresses involved in the induction and regulation of anthocyanin biosynthesis: A review. Afr. J. Biotechnol. 2008, 7, 4966–4972. [Google Scholar]
- Agati, G.; Stefano, G.; Biricolti, S.; Tattini, M. Mesophyll distribution of ‘’antioxidant“ flavonoid glycosides in ligustrum vulgare leaves under contrasting sunlight irradiance. Ann. Bot. 2009, 104, 853–861. [Google Scholar] [CrossRef] [PubMed]
- Agati, G.; Biricolti, S.; Guidi, L.; Ferrini, F.; Fini, A.; Tattini, M. The biosynthesis of flavonoids is enhanced similarly by UV radiation and root zone salinity in L. Vulgare leaves. J. Plant Physiol. 2011, 168, 204–212. [Google Scholar] [CrossRef] [PubMed]
- Koyama, K.; Ikeda, H.; Poudel, P.R.; Goto-Yamamoto, N. Light quality affects flavonoid biosynthesis in young berries of cabernet sauvignon grape. Phytochemistry 2012, 78, 54–64. [Google Scholar] [CrossRef] [PubMed]
- Agati, G.; Brunetti, C.; di Ferdinando, M.; Ferrini, F.; Pollastri, S.; Tattini, M. Functional roles of flavonoids in photoprotection: New evidence, lessons from the past. Plant Physiol. Biochem. 2013, 72, 35–45. [Google Scholar] [CrossRef] [PubMed]
- Pacheco, F.V.; Alvarenga, I.C.A.; Ribeiro Junior, P.M.; Pinto, J.E.B.P.; Avelar, R.D.P.; Alvarenga, A.A. Growth and production of secondary compounds in monkey-pepper (Piper aduncum L.) leaves cultivated under altered ambient light. Aust. J. Crop Sci. 2014, 8, 1510–1516. [Google Scholar]
- Gottschalk, K.W. Shade, leaf growth and crown development of Quercus rubra, Quercus velutina, Prunus serotina and Acer rubrum seedlings. Tree Physiol. 1994, 14, 735–749. [Google Scholar] [CrossRef] [PubMed]
- Müller, V.; Albert, A.; Barbro Winkler, J.; Lankes, C.; Noga, G.; Hunsche, M. Ecologically relevant UV-B dose combined with high PAR intensity distinctly affect plant growth and accumulation of secondary metabolites in leaves of Centella asiatica L. Urban. J. Photochem. Photobiol. B Biol. 2013, 127, 161–169. [Google Scholar] [CrossRef] [PubMed]
- Wang, M.L.; Jiang, Y.S.; Wei, J.Q.; Wei, X.; Qi, X.X.; Jiang, S.Y.; Wang, Z.M. Effects of irradiance on growth, photosynthetic characteristics, and artemisinin content of Artemisia annua L. Photosynthetica 2008, 46, 17–20. [Google Scholar] [CrossRef]
- Dai, Y.; Shen, Z.; Liu, Y.; Wang, L.; Hannaway, D.; Lu, H. Effects of shade treatments on the photosynthetic capacity, chlorophyll fluorescence, and chlorophyll content of Tetrastigma hemsleyanum Diels et Gilg. Environ. Exp. Bot. 2009, 65, 177–182. [Google Scholar] [CrossRef]
- Valladares, F.; Chico, J.; Aranda, I.; Balaguer, L.; Dizengremel, P.; Manrique, E.; Dreyer, E. The greater seedling high-light tolerance of Quercus robur over Fagus sylvatica is linked to a greater physiological plasticity. Trees 2002, 16, 395–403. [Google Scholar]
- Aleric, K.M.; Katherine Kirkman, L. Growth and photosynthetic responses of the federally endangered shrub, Lindera melissifolia (Lauraceae), to varied light environments. Am. J. Bot. 2005, 92, 682–689. [Google Scholar] [CrossRef] [PubMed]
- Tang, H.; Hu, Y.-Y.; Yu, W.-W.; Song, L.-L.; Wu, J.-S. Growth, photosynthetic and physiological responses of Torreya grandis seedlings to varied light environments. Trees 2015, 29, 1011–1022. [Google Scholar] [CrossRef]
- Shao, Q.; Wang, H.; Guo, H.; Zhou, A.; Huang, Y.; Sun, Y.; Li, M. Effects of shade treatments on photosynthetic characteristics, chloroplast ultrastructure, and physiology of Anoectochilus roxburghii. PLoS ONE 2014, 9, e85996. [Google Scholar] [CrossRef] [PubMed]
- Murchie, E.H.; Horton, P. Acclimation of photosynthesis to irradiance and spectral quality in british plant species: Chlorophyll content, photosynthetic capacity and habitat preference. Plant Cell Environ. 1997, 20, 438–448. [Google Scholar] [CrossRef]
- Liu, Y.; Li, X.; Liu, M.; Cao, B.; Tan, H.; Wang, J.; Li, X. Responses of three different ecotypes of reed (Phragmites communis trin.) to their natural habitats: Leaf surface micro-morphology, anatomy, chloroplast ultrastructure and physio-chemical characteristics. Plant Physiol. Biochem. 2012, 51, 159–167. [Google Scholar] [CrossRef] [PubMed]
- Saunders, J.A.; McClure, J.W. The distribution of flavonoids in chloroplasts of twenty five species of vascular plants. Phytochemistry 1976, 15, 809–810. [Google Scholar] [CrossRef]
- Agati, G.; Matteini, P.; Goti, A.; Tattini, M. Chloroplast-located flavonoids can scavenge singlet oxygen. New Phytol. 2007, 174, 77–89. [Google Scholar] [CrossRef] [PubMed]
- Zhai, Y.-K.; Guo, X.; Pan, Y.-L.; Niu, Y.-B.; Li, C.-R.; Wu, X.-L.; Mei, Q.-B. A systematic review of the efficacy and pharmacological profile of Herba epimedii in osteoporosis therapy. Pharmazie 2013, 68, 713–722. [Google Scholar] [PubMed]
- Li, C.; Li, Q.; Mei, Q.; Lu, T. Pharmacological effects and pharmacokinetic properties of icariin, the major bioactive component in Herba epimedii. Life Sci. 2015, 126, 57–68. [Google Scholar] [CrossRef] [PubMed]
- Liang, Q.; Wei, G.; Chen, J.; Wang, Y.; Huang, H. Variation of medicinal components in a unique geographical accession of horny goat weed Epimedium sagittatum maxim. (Berberidaceae). Molecules 2012, 17, 13345–13356. [Google Scholar] [CrossRef] [PubMed]
- Guo, B.-L.; He, S.-Z.; Zhong, G.-Y.; Xiao, P.-G. Two new species of Epimedium (Berberidaceae) from China. Acta Phytotaxon. Sin. 2007, 45, 813–821. [Google Scholar] [CrossRef]
- Sample Availability: Samples of the compounds are available from the authors.






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Analyte a | Linear Range (μg·mL−1) | Calibration Equation b | r2 | LOD (μg·mL−1) | LOQ (μg·mL−1) | Intra-Day RSD (%) (n = 6) | Inter-Day RSD (%) (n = 6) | Recovery and RSD (%) (Mean, n = 6) | ||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| L1 | L3 | L5 | L1 | L3 | L5 | |||||||
| 1 | 3.1~32.2 | Y = 7701.546x + 9983 | 0.9992 | 0.26 | 0.81 | 2.46 | 1.91 | 1.36 | 1.06 | 0.98 | 1.39 | 105.54, 3.32 |
| 2 | 3.1~32.2 | Y = 5548.525x + 15,762 | 0.9995 | 0.29 | 0.95 | 3.27 | 2.38 | 1.57 | 1.62 | 1.78 | 1.20 | 104.06, 2.96 |
| 3 | 20.5~520.6 | Y = 7171.274x + 31,544 | 0.9993 | 0.03 | 0.11 | 3.36 | 1.50 | 2.02 | 1.83 | 1.19 | 1.14 | 102.95, 3.37 |
| 4 | 3.1~32.2 | Y = 9108.011x + 6141 | 0.9996 | 0.11 | 0.37 | 2.50 | 1.66 | 1.46 | 1.79 | 1.54 | 1.44 | 104.92, 2.70 |
© 2016 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC-BY) license ( http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pan, J.; Guo, B. Effects of Light Intensity on the Growth, Photosynthetic Characteristics, and Flavonoid Content of Epimedium pseudowushanense B.L.Guo. Molecules 2016, 21, 1475. https://doi.org/10.3390/molecules21111475
Pan J, Guo B. Effects of Light Intensity on the Growth, Photosynthetic Characteristics, and Flavonoid Content of Epimedium pseudowushanense B.L.Guo. Molecules. 2016; 21(11):1475. https://doi.org/10.3390/molecules21111475
Chicago/Turabian StylePan, Junqian, and Baolin Guo. 2016. "Effects of Light Intensity on the Growth, Photosynthetic Characteristics, and Flavonoid Content of Epimedium pseudowushanense B.L.Guo" Molecules 21, no. 11: 1475. https://doi.org/10.3390/molecules21111475