
Laccase Gene Expression and Vinasse Biodegradation by Trametes hirsuta Strain Bm-2

Abstract
:
1. Introduction
2. Results and Discussion
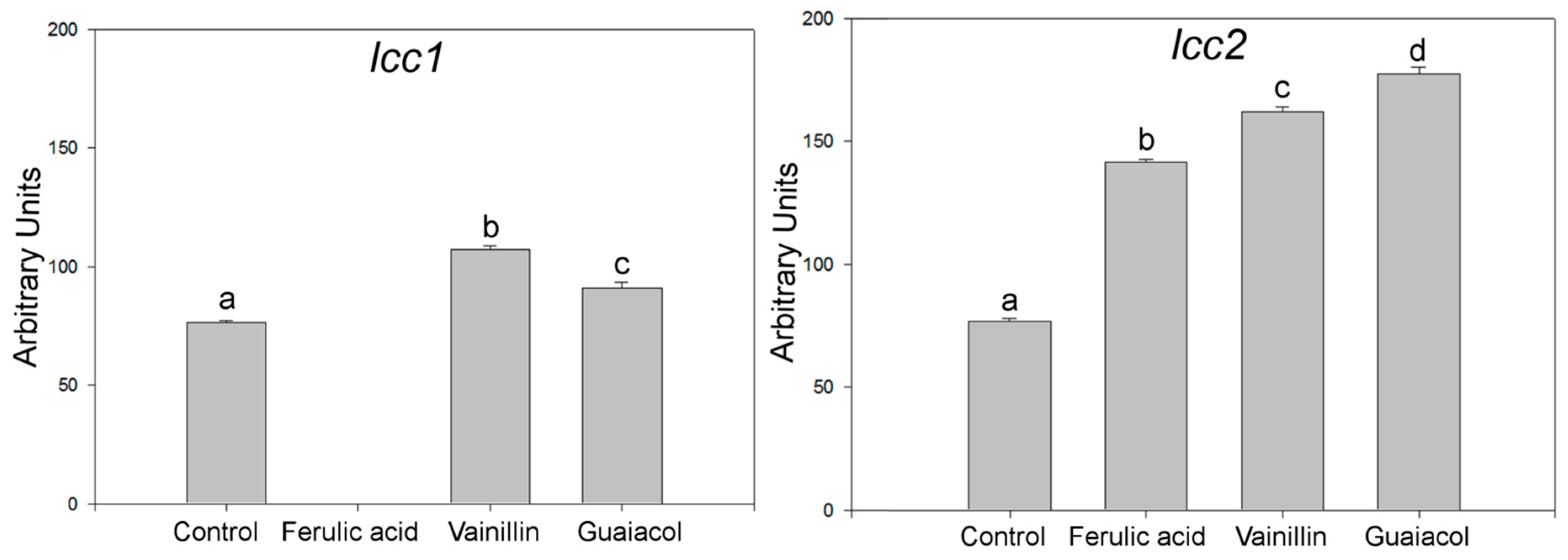
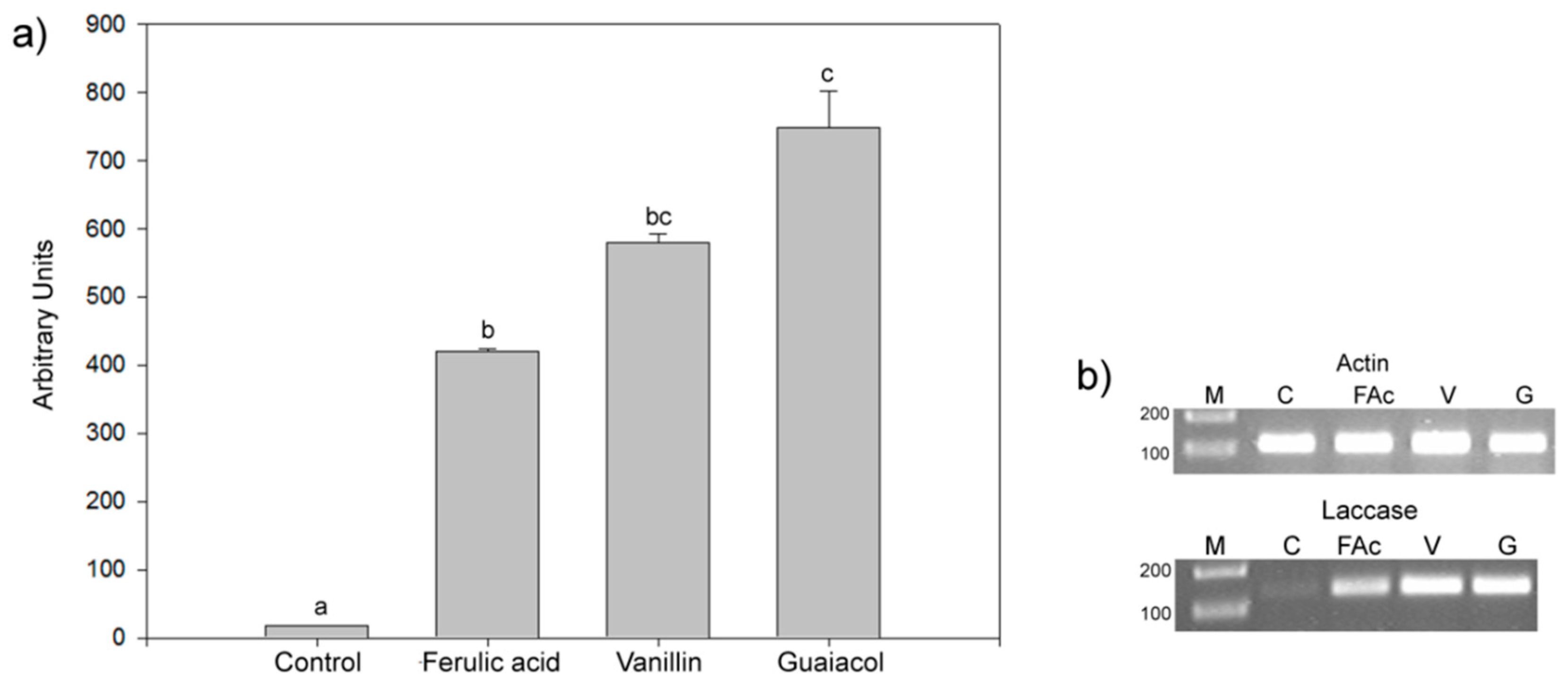
2.1. Effects of Aromatic Compounds on the Expression of Laccase Genes

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Compound | Chemical Structure | Laccase Activity (U/mL) |
|---|---|---|
| Ferulic acid |  | 380 |
| Vanillin |  | 450 |
| Guaiacol |  | 1700 |
| Control (Kirk’s medium) | --- | 150 |
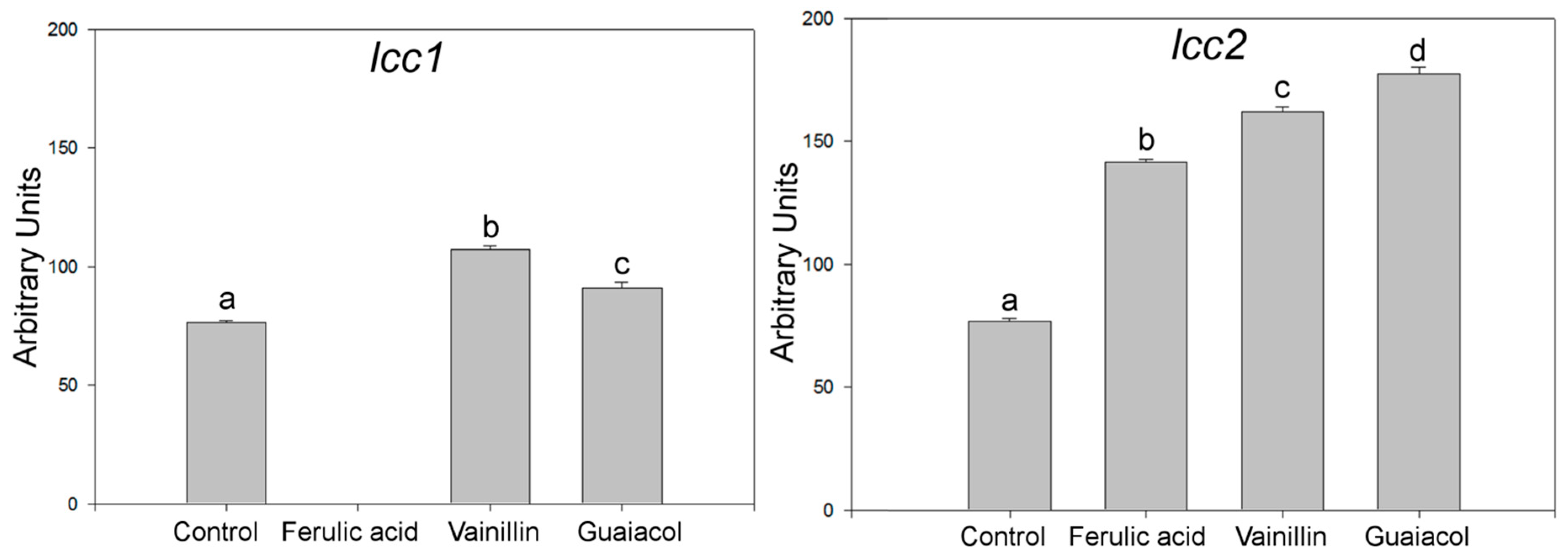
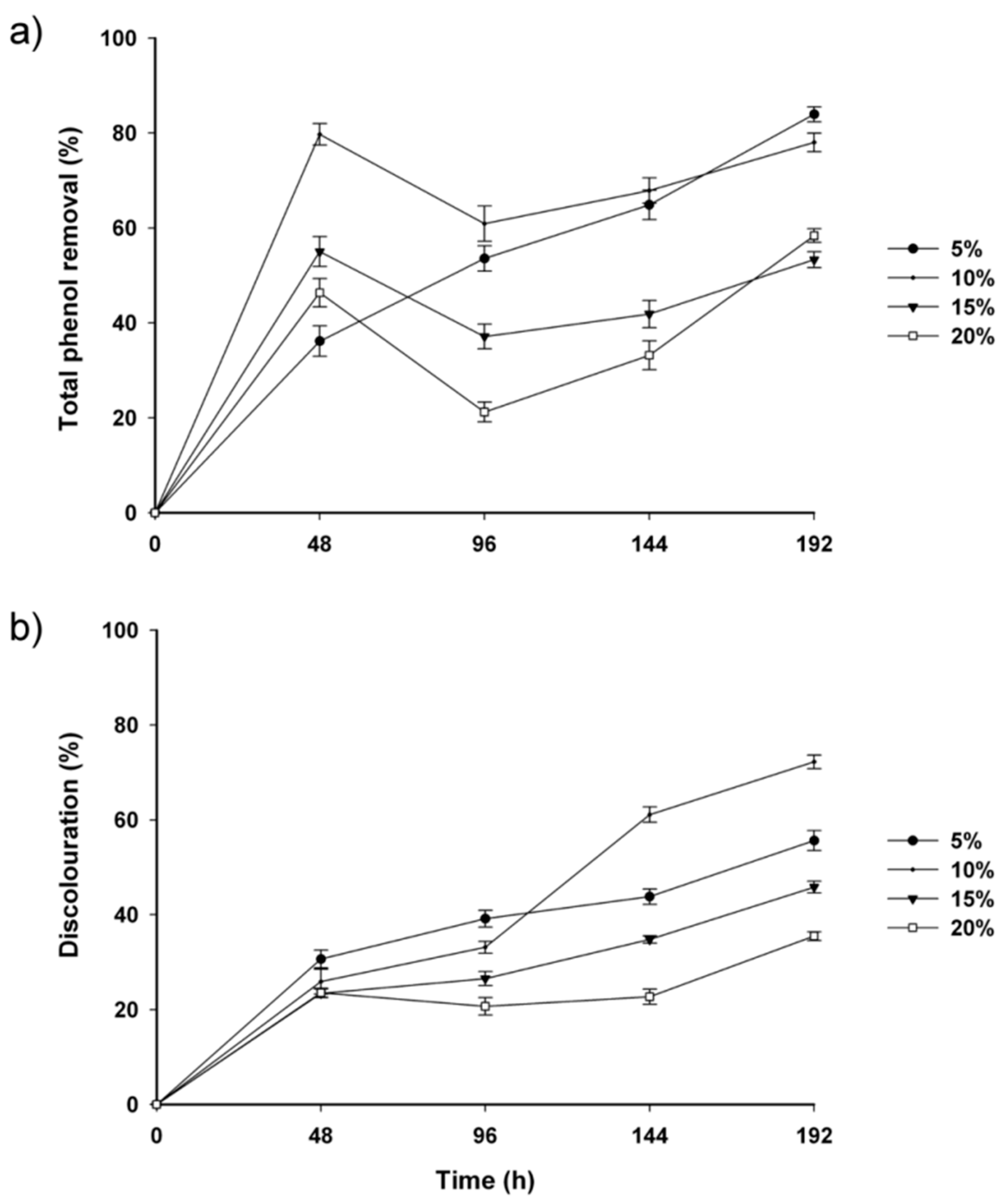
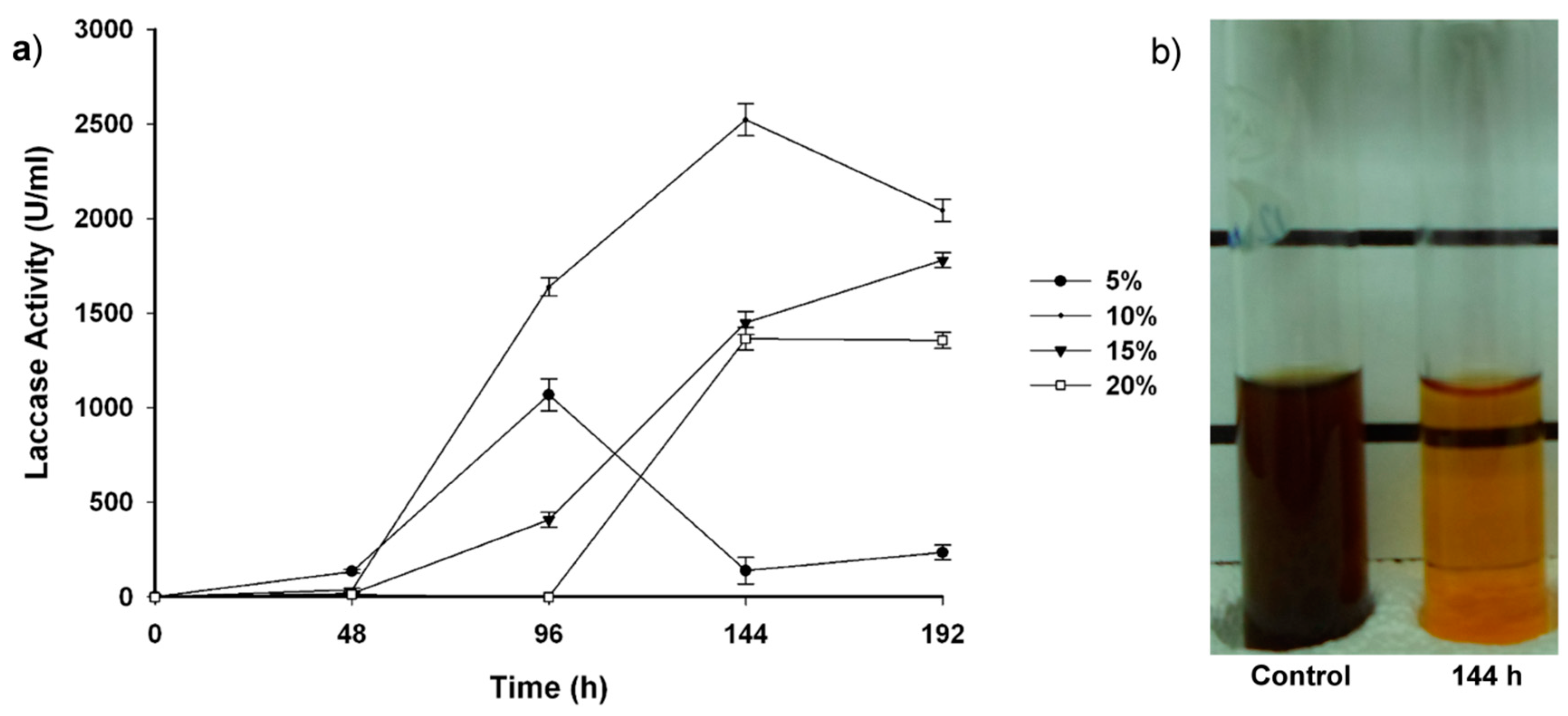
2.2. Fungal-Treated Vinasse
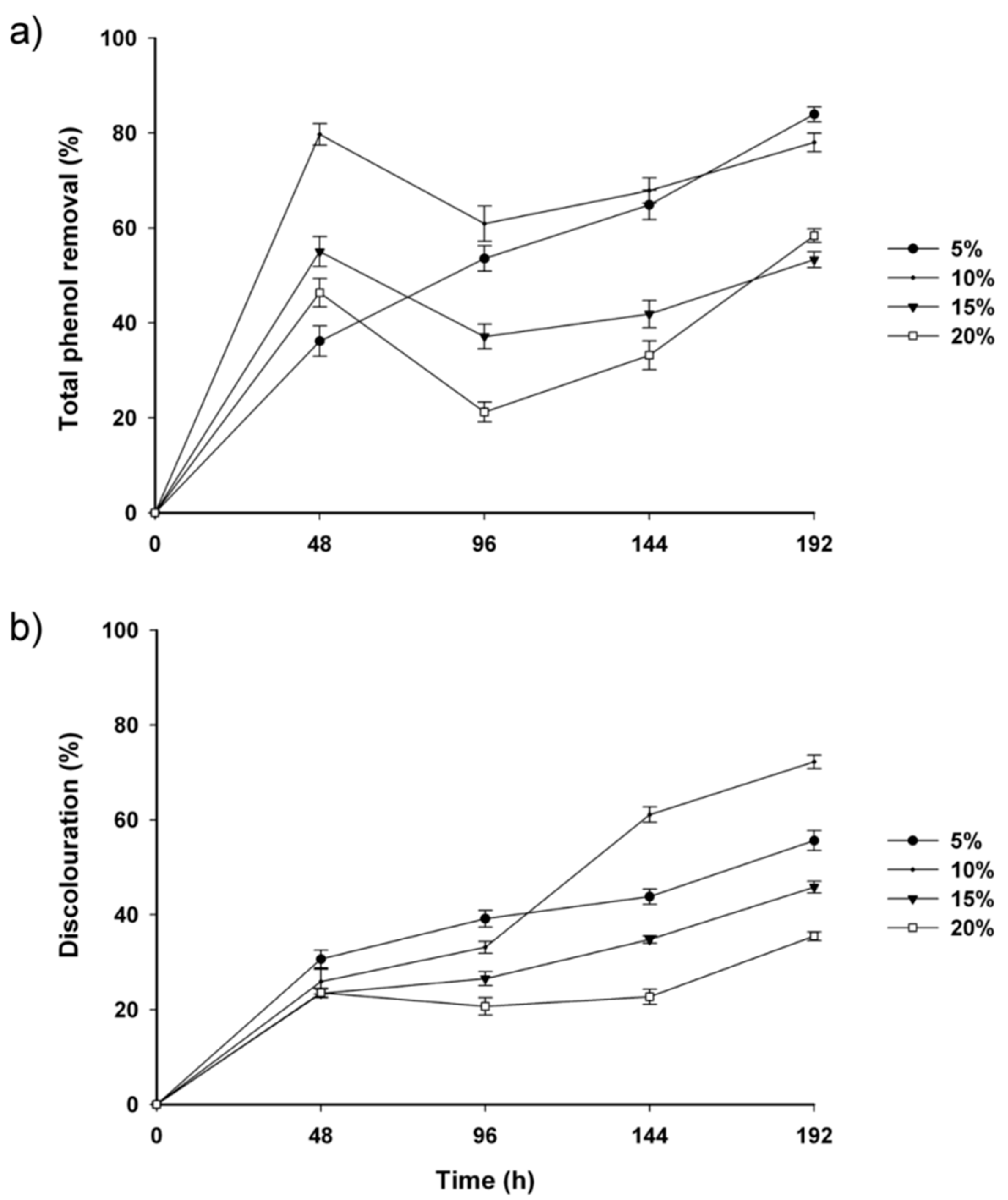
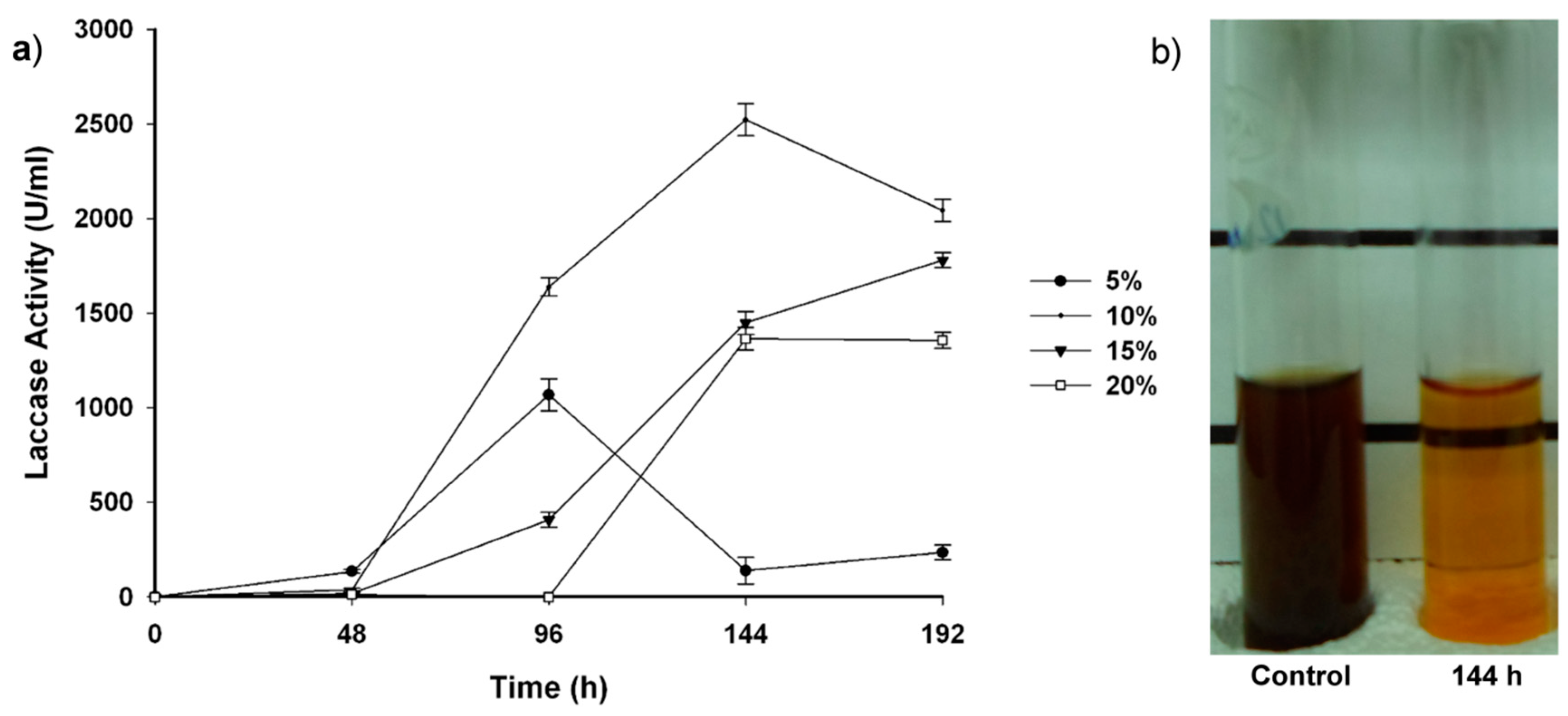
3. Experimental Section
3.1. Fungal Strain, Media, and Culture Conditions
3.2. Total RNA Preparation and cDNA Synthesis
3.3. PCR Amplification
3.4. Quantitative and Statistical Analyses
3.5. Vinasse
3.6. Preparation of Vinasse-Containing Medium
3.7. Total Phenolic Content and Discoloration
3.8. Laccase Activity
3.9. Statistical Analysis
4. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Potentini, M.F.; Rodríguez-Malaver, A.J. Vinasse biodegradation by Phanerochaete chrysosporium. J. Environ. Biol. 2006, 27, 661–665. [Google Scholar] [PubMed]
- Arimi, M.M.; Zhang, Y.; Götz, G.; Kiriamiti, K.; Geißen, S.U. Antimicrobial colorants in molasses distillery wastewater and their removal technologies. Int. Biodeterior. Biodegrad. 2014, 87, 34–43. [Google Scholar] [CrossRef]
- Ferreira, L.F.; Pompeu, G.; Aguiar, M.; Messias, T.G.; Monteiro, R.R. Selection of vinasse degrading microorganisms. World J. Microbiol. Biotechnol. 2010, 26, 1613–1621. [Google Scholar] [CrossRef]
- Mussatto, S.I.; Dragone, G.; Guimarães, P.M.; Silva, J.P.A.; Carneiro, L.M.; Roberto, I.C.; Vicente, A.; Domingues, L.; Teixeira, J.A. Technological trends, global market, and challenges of bio-ethanol production. Biotechnol. Adv. 2010, 28, 817–830. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- González, T.; Terrón, M.C.; Yague, S.; Zapico, E.; Galletti, G.C.; González, A.E. Pyrolisis/gas chromatography/mass spectrometry monitoring of fungal-biotreated distillery wastewater using Trametes sp. I-62 (CECT 20197). Rapid Commun. Mass Spectrom. 2000, 14, 1417–1424. [Google Scholar] [CrossRef]
- Bibi, I.; Bhatti, H.N.; Asgher, M. Comparative study of natural and synthetic phenolic compounds as efficient laccase mediators for the transformation of cationic dye. Biochem. Eng. J. 2011, 56, 225–231. [Google Scholar] [CrossRef]
- Saraiva, J.A.; Tavares, A.P.M.; Xavier, A.M.R.B. Effect of the inducers veratryl alcohol, xylidine, and ligninosulphonates on activity and thermal stability and inactivation kinetics of laccase from Trametes versicolor. Appl. Biochem. Biotechnol. 2012, 167, 685–693. [Google Scholar] [CrossRef] [PubMed]
- Strong, P.J.; Claus, H. Laccase: A review of its past and its future in bioremediation. Crit. Rev. Environ. Sci. Technol. 2011, 41, 373–434. [Google Scholar] [CrossRef]
- Thurston, C.F. The structure and function of fungal laccases. Microbiology 1994, 140, 19–26. [Google Scholar] [CrossRef]
- Goudopoulou, A.; Krimitzas, A.; Milton, A.T. Differential gene expression of ligninolytic enzymes in Pleurotus ostreatus grown on olive oil mill wastewater. Appl. Microbiol. Biotechnol. 2010, 88, 541–551. [Google Scholar] [CrossRef] [PubMed]
- Terrón, M.C.; González, T.; Carbajo, J.M.; Yagüe, S.; Arana-Cuenca, A.; Téllez, A.; Dobson, A.D.W.; González, A.E. Structural close-related aromatic compounds have different effects on laccase activity and lcc gene expression in the ligninolytic fungus Trametes sp. I-62. Fungal Genet. Biol. 2004, 41, 954–962. [Google Scholar] [CrossRef] [PubMed]
- González, T.; Terrón, M.C.; Zapico, E.J.; Yagüe, S.; Téllez, A.; Junca, H.; González, A.E. Identification of a new laccase gene and confirmation of genomic preditions by cDNA sequences of Trametes sp. I-62 laccase familiy. Mycol. Res. 2003, 107, 727–735. [Google Scholar] [CrossRef] [PubMed]
- Luis, P.; Kellner, H.; Martin, F.; Buscot, F. A molecular method to evaluate basidiomycete laccase gene expression in forest soils. Geoderma 2005, 128, 18–27. [Google Scholar] [CrossRef]
- Pérez, S.R.; Savón, R.C.; Diaz, M.S.; Kourouma, A. Selección de cepas de Pleurotus ostreatus para la decoloración de efluentes industriales. Rev. Mex. Micol. 2006, 23, 9–15. [Google Scholar]
- Chandra, R.; Bharagava, R.N.; Rai, V. Melanoidins as major colourant in sugarcane molasses based distillery effluent and its degradation. Bioresour. Technol. 2008, 99, 4648–4660. [Google Scholar] [CrossRef] [PubMed]
- Naik, N.M.; Jagadeesh, K.S.; Alagawadi, A.R. Microbial decolorization of spentwash: A review. Indian J. Microbiol. 2008, 48, 41–48. [Google Scholar] [CrossRef] [PubMed]
- Hammel, K.E. Oxidation of aromatic pollutants by lignin degrading fungi and their extracellular peroxidases. In Metal Ions in Biological Systems; Siegel, H., Siegel, A., Eds.; Marcel Dekker: New York, NY, USA, 1992; Volume 28, p. 41. [Google Scholar]
- Payet, B.; Shum Cheong Sing, A.; Smadja, J. Comparison of the concentrations of phenolic constituents in cane sugar manufacturing products with their antioxidant activities. J. Agric. Food Chem. 2006, 54, 7270–7276. [Google Scholar] [CrossRef] [PubMed]
- Vaquero, M.J.R.G.; Alberto, M.R.; de Nadra, M.C.M. Antibacterial effect of phenolic compounds from different wines. Food Control 2007, 18, 93–101. [Google Scholar] [CrossRef]
- Jimenez, A.M.; Borja, R.; Martin, A. Aerobic-anaerobic biodegradation of beet molasses alcoholic fermentation wastewater. Process. Biochem. 2003, 38, 1275–1284. [Google Scholar] [CrossRef]
- Tapia-Tussell, R.; Pérez-Brito, D.; Rojas-Herrera, R.; Cortes-Velazquez, A.; Rivera- Muñoz, G.; Solis-Pereira, S. New laccase-producing fungi isolates with biotechnological potential in dye decolorization. Afr. J. Biotechnol. 2011, 10, 10134–10142. [Google Scholar]
- Tien, M.; Kirk, K. Lignin-degrading enzymes from Phanerochaete chrysosporium: Purification, characterization, and catalytic properties of a unique H2O2-requiring oxygenase. Proc. Natl. Acad. Sci. USA 1984, 81, 2280–2284. [Google Scholar] [CrossRef] [PubMed]
- Kirk, T.K.; Croan, S.; Tien, M.; Murtagh, K.E.; Farrell, R.L. Production of multiple ligninases by Phanerochaete chrysosporium, effect of the selected growth conditions and the use of a mutant strain. Enzyme Microb. Technol. 1986, 8, 27–32. [Google Scholar] [CrossRef]
- Burgos, J.M.N. Regulación de la expresión de lacasas por metales y fenoles simples en Trametes hirsuta Bm-2. Master’s Thesis, Instituto Tecnológico de Mérida, Yucatán, Mexico, 12 November 2010. [Google Scholar]
- D’Souza, T.M.; Boominathan, K.; Reddy, C.A. Isolation of laccase gene-specific sequences from white rot and brown rot fungi by PCR. Appl. Environ. Microbiol. 1996, 62, 3739–3744. [Google Scholar] [PubMed]
- González, T.; Terrón, M.C.; Zapico, E.J.; Téllez, A.; Yague, S.; Carbajo, J.M.; González, A.E. Use of multiplex reverse transcription-PCR to study the expression of a laccase gene family in a basidiomycetous fungus. Appl. Environ. Microbiol. 2003, 69, 7083–7090. [Google Scholar] [CrossRef] [PubMed]
- Carbone, I.; Kohn, L.M. A method for designing primers sets for speciation studies in filamentous ascomycetes. Mycologia 1999, 91, 553–556. [Google Scholar] [CrossRef]
- Waterman, P.G.; Mole, S. Methods in Ecology: Analysis of Phenolic Plant Metabolites; Blackwell Scientific Publications: Oxford, Boston, MA, USA, 1994. [Google Scholar]
- Sirianuntapiboon, S.; Sihanonth, P.; Somchai, P.; Atthasampunna, P.; Hayashida, S. An adsorption mechanism for melanoidin decolorization by Rhizoctonia sp. Biosci. Biotechnol. Biochem. 1995, 59, 1185–1189. [Google Scholar] [CrossRef]
- Wolfenden, B.S.; Willson, R.L. Radical-cations as reference chromogens in kinetic studies of one-electron transfer reactions: pulse radiolysis studies of 2,2′-azinobis-(3-ethylbenzthiazoline-6-sulphonate). J. Chem. Soc. Perkin Trans. 2 1982, 7, 805–812. [Google Scholar] [CrossRef]
- Sample Availability: Ferulic acid, Vanillin and guaiacol are available.
© 2015 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license ( http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tapia-Tussell, R.; Pérez-Brito, D.; Torres-Calzada, C.; Cortés-Velázquez, A.; Alzate-Gaviria, L.; Chablé-Villacís, R.; Solís-Pereira, S. Laccase Gene Expression and Vinasse Biodegradation by Trametes hirsuta Strain Bm-2. Molecules 2015, 20, 15147-15157. https://doi.org/10.3390/molecules200815147
Tapia-Tussell R, Pérez-Brito D, Torres-Calzada C, Cortés-Velázquez A, Alzate-Gaviria L, Chablé-Villacís R, Solís-Pereira S. Laccase Gene Expression and Vinasse Biodegradation by Trametes hirsuta Strain Bm-2. Molecules. 2015; 20(8):15147-15157. https://doi.org/10.3390/molecules200815147
Chicago/Turabian StyleTapia-Tussell, Raúl, Daisy Pérez-Brito, Claudia Torres-Calzada, Alberto Cortés-Velázquez, Liliana Alzate-Gaviria, Rubí Chablé-Villacís, and Sara Solís-Pereira. 2015. "Laccase Gene Expression and Vinasse Biodegradation by Trametes hirsuta Strain Bm-2" Molecules 20, no. 8: 15147-15157. https://doi.org/10.3390/molecules200815147