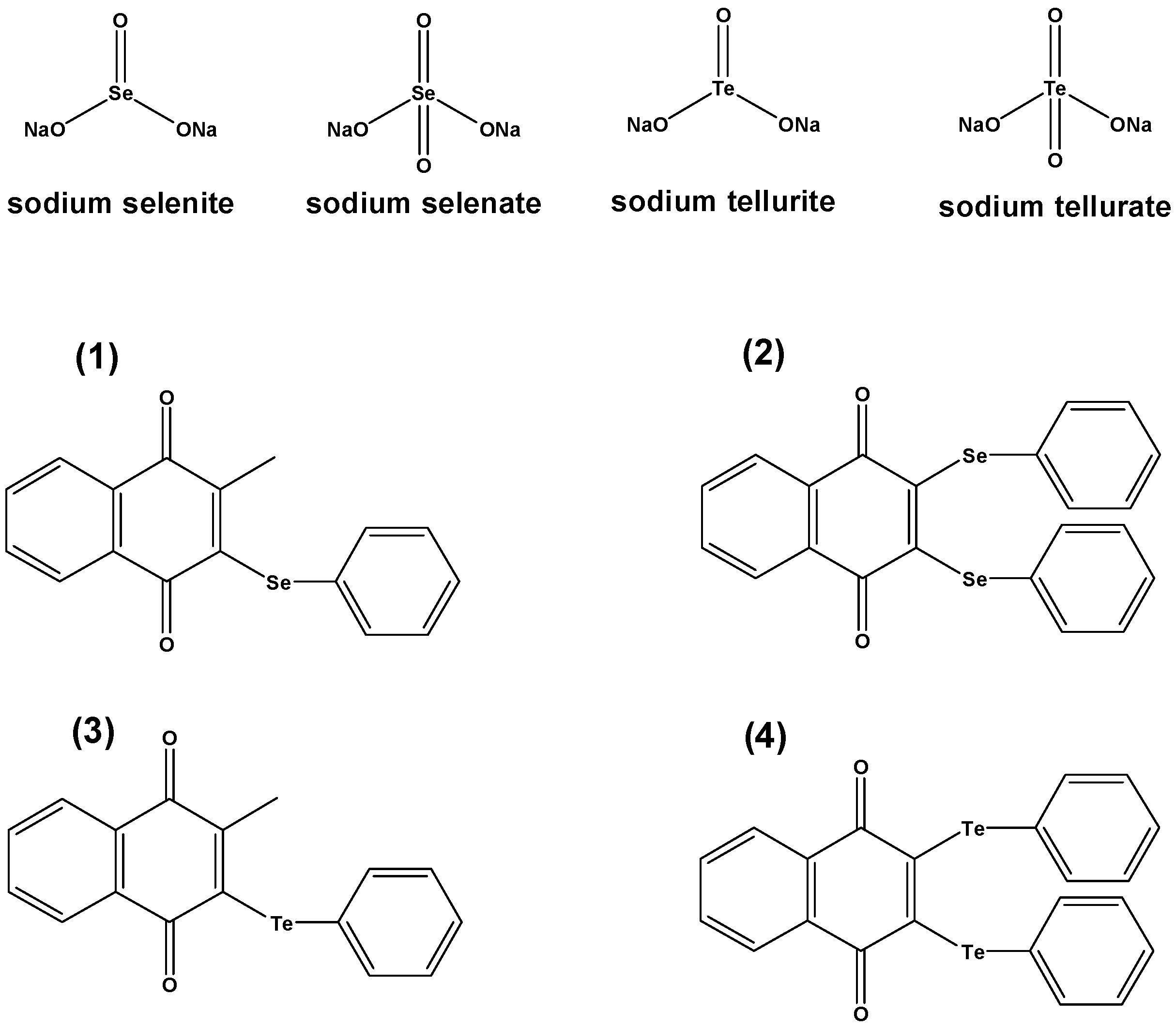
To our great surprise we have found that some simple selenium and tellurium salts act in concert with known antibiotics, hence apparently “sensitizing” such bacteria—which include strains of MRSA—against conventional antibiotics, possibly by inferring with the protease machinery/proteasome. Since such salts are not optimized for a particular biological activity and are also unlikely to penetrate the cell easily, the concentrations required for action were high. At the same time, the formation of red and black deposits could be observed in the case of certain selenium and tellurium salts, respectively. We have therefore subsequently turned our attention towards organic selenium (and tellurium) compounds, and as expected, these compounds show a higher activity. These findings will now be presented and discussed in more detail.
2.1. Antimicrobial Activity of Selenium Salts
Since selenium salts, such as Na
2SeO
3, nowadays are used widely in form of food supplements, we have first investigated the potential antimicrobial activity of such simple selenium salts and compared this activity to the one of their sulfur and tellurium analogues. This comparison is reasonable as tellurite has long been considered as possible antibiotic and sulfite in fact is used extensively as antimicrobial agent to preserve fruits and vegetable nuts. At the high concentrations used, however, neither selenite or selenate, nor any of the other salts, showed any notable activity against
S. aureus reference strain ATCC 25923 or multidrug resistant MRSA HEMSA 5 or MRSA HEMSA 5M strains. Indeed, it appears that the MIC values against those bacteria in most instances are well above 1 mM, which is much higher than those of conventional effective antibiotics (
Table 1).
Table 1.
MIC values (in μM) of selected chalcogen salts against three strains of S. aureus. Whilst these salts are clearly not active against any of the strains at pharmaceutically relevant concentrations, conventional antibiotics show some activity, which is reduced significantly in the case of the two resistant strains.
Table 1.
MIC values (in μM) of selected chalcogen salts against three strains of S. aureus. Whilst these salts are clearly not active against any of the strains at pharmaceutically relevant concentrations, conventional antibiotics show some activity, which is reduced significantly in the case of the two resistant strains.
| Compound | MIC Values (μM) in S. aureus Strains |
|---|
| ATCC 25923 | MRSA HEMSA 5 | MRSA HEMSA 5M |
|---|
| Salt | Na2TeO3 | 1000 | >2000 | >2000 |
| Na2SeO3 | >2000 | >2000 | >2000 |
| Na2SeO4 | >2000 | >2000 | >2000 |
| Na2SO3 | >2000 | >2000 | >2000 |
| Na2SO4 | >2000 | >2000 | >2000 |
| Organic | 1 | 31.25 | 125 | 500 |
| 2 | 250 | 500 | >1000 |
| 3 | 62.5 | 62.5 | >1000 |
| 4 | 31.25 | 31.25 | 125 |
| Antibiotic | Oxacillin | 0.45 | 340 | 5400 |
| Cloxacillin | 0.42 | 160 | 1300 |
| Ampicillin/sulbactam | 0.54 | 200 | 200 |
| Ciprofloxacin | 0.38 | 14 | 28 |
| Neomycin | 1.0 | 240 | 240 |
This implies, of course, that neither the selenium nor the tellurium or sulfur salts used as part of this study on their own could be employed as antibiotics in practice. Similar results have been obtained against
Saccharomyces cerevisiae, where neither selenite, selenate nor tellurite showed any significant activity against this type of fungus [
3]. It should be noted that tellurate (Na
2TeO
4) was also considered, yet could not be tested reliably because of solubility issues in the buffers used which resulted in irreproducible results.
This rather disappointing finding is not entirely unexpected: such simple, highly polar salts are not optimized for antibiotic action and their ability to cross cell membranes is limited. Nonetheless, the interactions of such salts with biomolecules, especially cysteine containing proteins, are well documented
in vitro, and one would be tempted to expect at least some impact on the bacterial, fungal or mammalian cell. Based on our previous studies in this field, one may speculate, for instance, that some of these compounds, particularly Na
2SeO
3, could “weaken” the cell by promoting oxidative stress and damage to DNA or by affecting the cellular thiolstat [
10,
11].
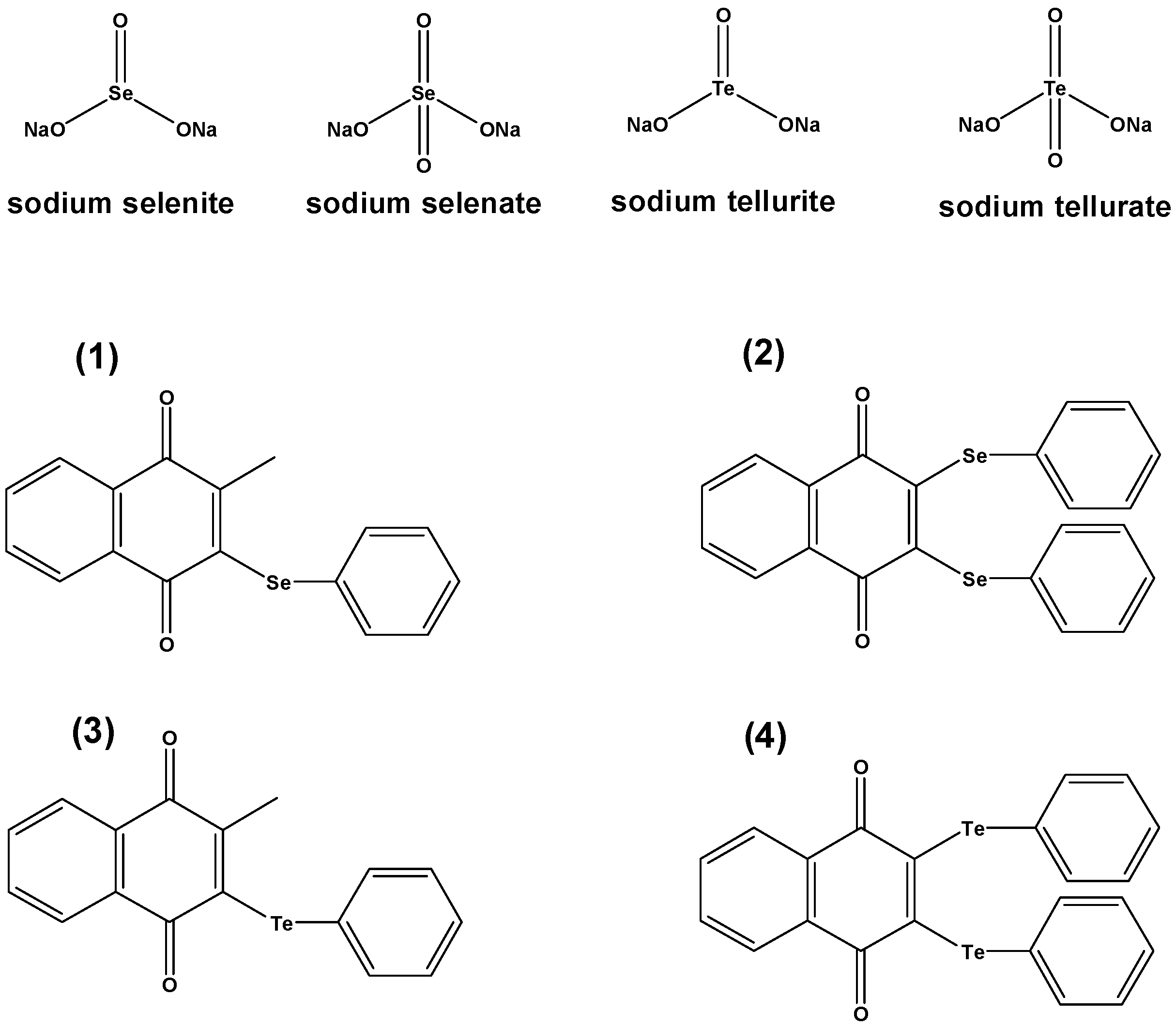
In the case of the organic selenium and tellurium compounds
1–
4, a slight antibacterial activity could be observed when these compounds were used in micromolar concentrations. While this activity often was distinctively lower than that of the reference antibiotics used, it was much higher when compared to the inorganic salts—and was also rather competitive in the case of the drug resistant strains. The tellurium compounds
3 and
4, in particular, showed some promising activity against the multidrug resistant strain HEMSA-5. The
bis-phenyltellanyl derivative
4 was even able to inhibit the growth of both MDR MRSA strains (HEMSA 5 and extremely resistant HEMSA 5M) at a dose significantly lower than that of most antibiotics tested (excluding ciprofloxacin, see
Table 1).
Since it appears that the selenium and tellurium compounds act differently, probably on different targets when compared to the conventional antibiotics, we have posed the question if they may not be able to also act synergistically,
i.e., in concert with conventional agents. We have therefore investigated both, the organic compounds
1–
4 and some of the salts in combination with known antibiotic agents, such as oxacillin, cloxacillin, ampicillin/sulbactam, ciprofloxacin and neomycin, and indeed observed a significant enhancement in the antibacterial activity of such traditional antibiotics, even against some of the strains of MRSA (
Table 2). Notably, whilst the inorganic salts often enhanced the efficiency of the antibiotics used, the organic compounds
1–
4 were not particularly useful in those combination assays, bearing in mind, of course, that compounds
3 and
4 are fairly potent antibiotics on their own (see above). Particularly noteworthy, for instance, is the influence of tellurite (Na
2TeO
3), which at a concentration of just 500 µM enhances the toxicity of oxacillin and cloxacillin against MRSA HEMSA 5M by more than 1000-fold (
Table 2). Notably, Na
2TeO
3 does not seem to exhibit any synergistic effects against the reference strain
S. aureus ATTC 25923.
Table 2.
Ability of the chalcogen compounds tested to enhance the antibacterial activity of selected antibiotics against S. aureus strains at a chalcogen compound concentration of 500 µM. Results are expressed as the quotient of the MIC value of antibiotics in the absence to that in the presence of the corresponding chalcogen compound.
Table 2.
Ability of the chalcogen compounds tested to enhance the antibacterial activity of selected antibiotics against S. aureus strains at a chalcogen compound concentration of 500 µM. Results are expressed as the quotient of the MIC value of antibiotics in the absence to that in the presence of the corresponding chalcogen compound.
| Antibiotic | Strain of S. aureus | Antibiotic Efficacy Enhancement |
|---|
| 1–4 | Na2TeO3 | Na2SeO3 | Na2SeO4 | Na2SO3 | Na2SO4 |
|---|
| Oxacillin | ATCC 25923 | NE | NE * | 4 | 4 | NE | NE |
| MRSA HEMSA 5 | NE | 16 | NE | NE | NE | NE |
| MRSA HEMSA 5M | NE | 1024 | NE | NE | NE | NE |
| Cloxacillin | ATCC 25923 | NE | 2 * | 4 | 4 | NE | NE |
| MRSA HEMSA 5 | NE | 256 | NE | NE | 32 | 32 |
| MRSA HEMSA 5M | NE | ≥1024 | NE | NE | NE | NE |
| Ampicillin/Sulbactam | ATCC 25923 | NE | 4 * | 2 | 2 | NE | NE |
| MRSA HEMSA 5 | NE | 8 | NE | NE | 4 | 4 |
| MRSA HEMSA 5M | NE | 8 | NE | NE | NE | NE |
| Ciprofloxacin | ATCC 25923 | NE | NE * | NE | NE | NE | NE |
| MRSA HEMSA 5 | NE | NE | NE | NE | NE | NE |
| MRSA HEMSA 5M | NE | NE | NE | NE | NE | NE |
| Neomycin | ATCC 25923 | NE | 2 * | NE | NE | NE | NE |
| MRSA HEMSA 5 | NE | 4–8 | 8 | NE | NE | NE |
| MRSA HEMSA 5M | NE | 4 | NE | NE | NE | NE |
These findings are rather interesting, as they indicate a certain “re-sensitization” of the drug resistant strains against classic antibiotics in the presence of certain selenium, and possibly also of tellurium or sulfur compounds. Tellurite, for instance promotes the activity of β-lactam antibiotics oxacillin and cloxacillin against both strains of MRSA studied, reducing the MIC values for these antibiotics from well over 100 µM in the case of HEMSA 5 and over 1000 µM in the case of HEMSA 5M to acceptable values in the sub- or low micromolar range, respectively. Furthermore, tellurite increases the action of ampicillin combined with sulbactam or of the aminoglycoside antibiotic neomycin against multidrug resistant MRSA strains (
Table 2).
This finding in itself obviously is exceptionally attractive from the perspective of future drug combination therapies, which to a large extent will rely on this kind of re-sensitization of resistant strains. It also agrees with a very recent report in the literature, which shows that nanomolar concentrations of Na
2TeO
3 can enhance the toxic effects of the cephalosporin antibiotic cefotaxime against
Escherichia coli [
12]. In this study, the enhancement by Na
2TeO
3 appeared to be due to a tellurite-induced oxidative stress, and a widespread damage to DNA and to proteins, possible mode(s) of action which will be considered in more detail as part of the following sections.
Unfortunately, tellurite is also rather toxic in humans, and the two selenium salts are less active, yet also seem to be able to somewhat, up to 4- to 8-fold, promote the toxicity of different antibiotics. Na2SeO3, in particular, seems to enhance the action of aminoglycosides against multidrug resistant MRSA HEMSA 5, decreasing the effective dose of neomycin required from 240 µM to 30 µM. Bearing in mind that Na2SeO3 and Na2SeO4 are considerably less toxic towards humans when compared to their tellurium analogues, such an enhancement of antibiotic action in MRSA strains is rather interesting. The ultimate choice for a potential therapeutic use therefore may well consider selenite and selenate, rather than tellurite, and obviously at concentrations not damaging or even lethal to humans. Incidentally, an interesting activity of sulfite and sulfate—which at first were included in the study as controls—could be demonstrated. The toxicity of cloxacillin against MRSA HEMSA 5, for instance, is enhanced around 32-fold by 500 µM concentrations of either Na2SO3 or Na2SO4. Since sulfate, in particular, is non-toxic to humans even when administered at higher concentrations, this finding—and the possible usage of sulfur-based salts as potential “adjuvants” of this β-lactam antibiotic—should also be considered further in earnest.
2.2. Cellular Targets of Selenite and Selenate in Bacteria and Yeast
Whilst some of the selenium and tellurium salts studied seem to enhance the antibiotic activity of traditional antibiotics, such as β-lactams and aminoglycosides, none of chalcogen salts employed was able to increase the bactericidal activity of the fluoroquinolone ciprofloxacin. Ciprofloxacin is an inhibitor of bacterial gyrase and hence DNA replication. In contrast, the penicillins interfere with the bacterial cell wall synthesis. It is therefore possible that the synergistic effects observed in the case of the penicillins point toward a particular mode of action of salts such as Na
2TeO
3, Na
2SeO
3 or Na
2SeO
4. We have already mentioned in passing that based on the chemistry and
in vitro studies of such redox active selenium and tellurium compounds, one would expect a “weakening” of cells exposed to higher concentrations of these agents. Although speculative at this time, it may be possible, for instance, that these salts interfere with the cell membrane or components thereof. They may hinder the cell wall synthesis, perhaps inhibit (cysteine containing) β-lactamases or stress the cell more generally, for instance by the generation of Reactive Oxygen Species (ROS) [
13,
14]. Indeed, such actions have been associated with selenite and tellurite before [
3,
12].
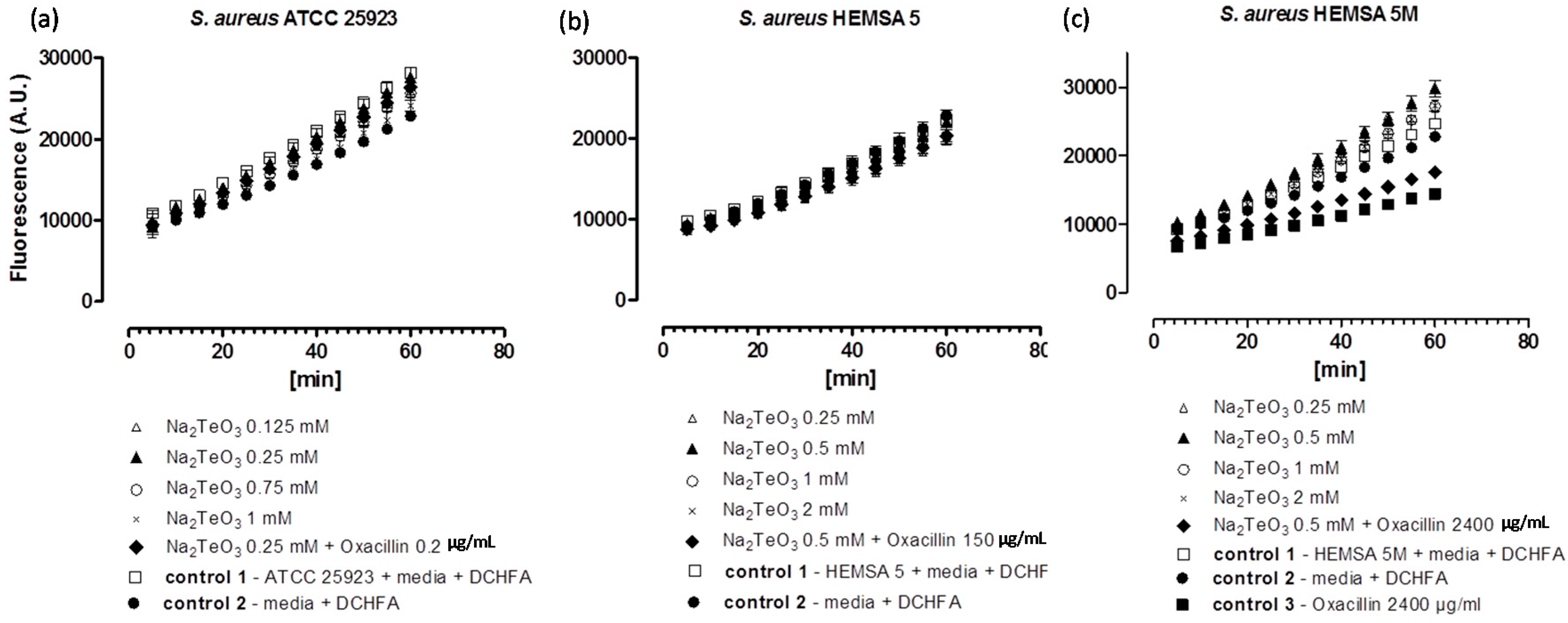
To investigate this issue further, we have therefore considered the impact of these chalcogen salts on intracellular ROS levels in
S. aureus ATCC 25923, HEMSA 5 and HEMSA 5M using 2ʹ,7ʹ-dichlorodihydrofluorescein diacetate (DCHFA) as redox-sensitive fluorescent indicator dye. The results of this crude ROS assay are presented for the different salts, and against different strains of
S. aureus in
Figure 2,
Figure 3 and
Figure 4.
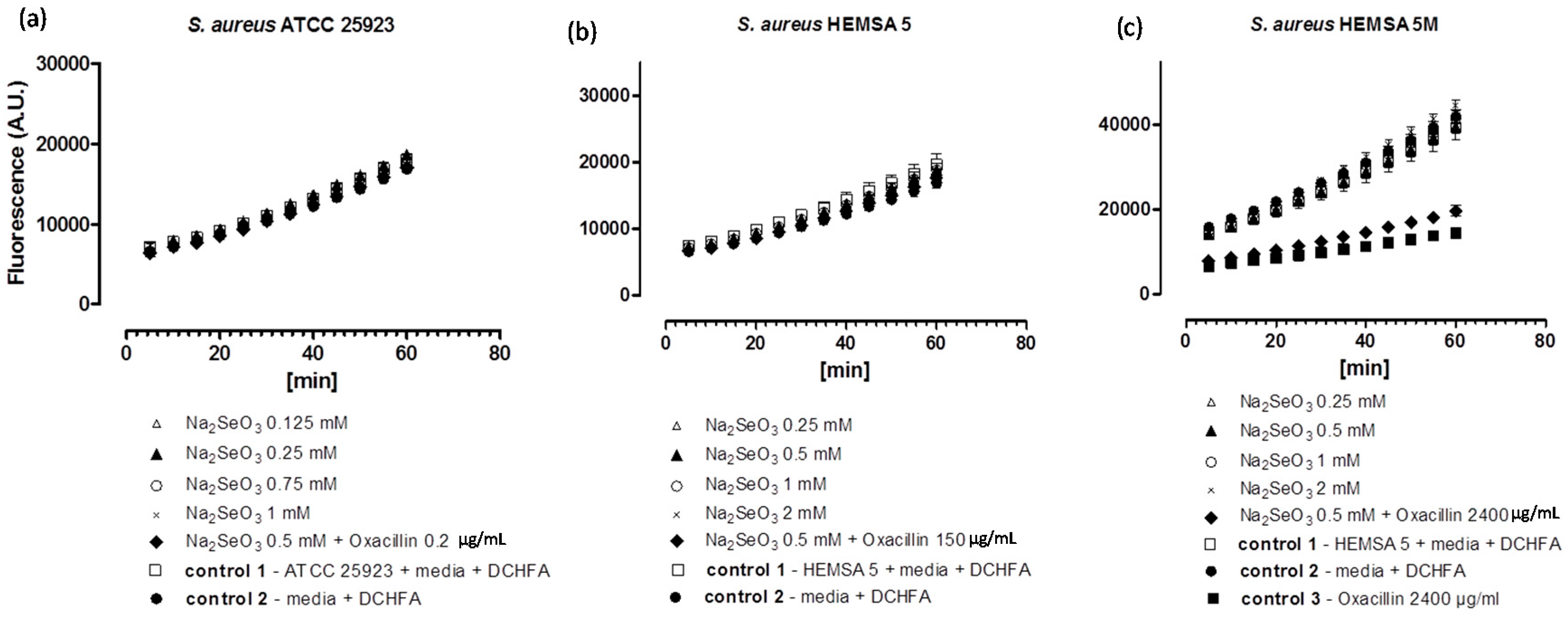
Figure 2.
Generation of intracellular ROS by Na
2SeO
3 in different strains of
S. aureus. ATCC 25923 (
a); HEMSA 5 (
b) and HEMSA 5M (
c). Data is shown in terms of fluorescence emitted (in arbitrary units) by (oxidized) DCFA. The latter is generated by the reaction of certain ROS and Reactive Nitrogen Species (RNS) with DCHFA. Different concentrations of Na
2SeO
3 were assayed and the influence of the dual presence of the chalcogen salt and an antibiotic (oxacillin) on ROS generation was also evaluated. Values shown represent mean values with
n = 4. The error bars represent the standard deviation (SD) and statistical significances were calculated using an one-way ANOVA followed by Bonferroni’s multiple comparison test (
Figure 2a,b:
p > 0.05; 2c:
p < 0.05).
Figure 2.
Generation of intracellular ROS by Na
2SeO
3 in different strains of
S. aureus. ATCC 25923 (
a); HEMSA 5 (
b) and HEMSA 5M (
c). Data is shown in terms of fluorescence emitted (in arbitrary units) by (oxidized) DCFA. The latter is generated by the reaction of certain ROS and Reactive Nitrogen Species (RNS) with DCHFA. Different concentrations of Na
2SeO
3 were assayed and the influence of the dual presence of the chalcogen salt and an antibiotic (oxacillin) on ROS generation was also evaluated. Values shown represent mean values with
n = 4. The error bars represent the standard deviation (SD) and statistical significances were calculated using an one-way ANOVA followed by Bonferroni’s multiple comparison test (
Figure 2a,b:
p > 0.05; 2c:
p < 0.05).
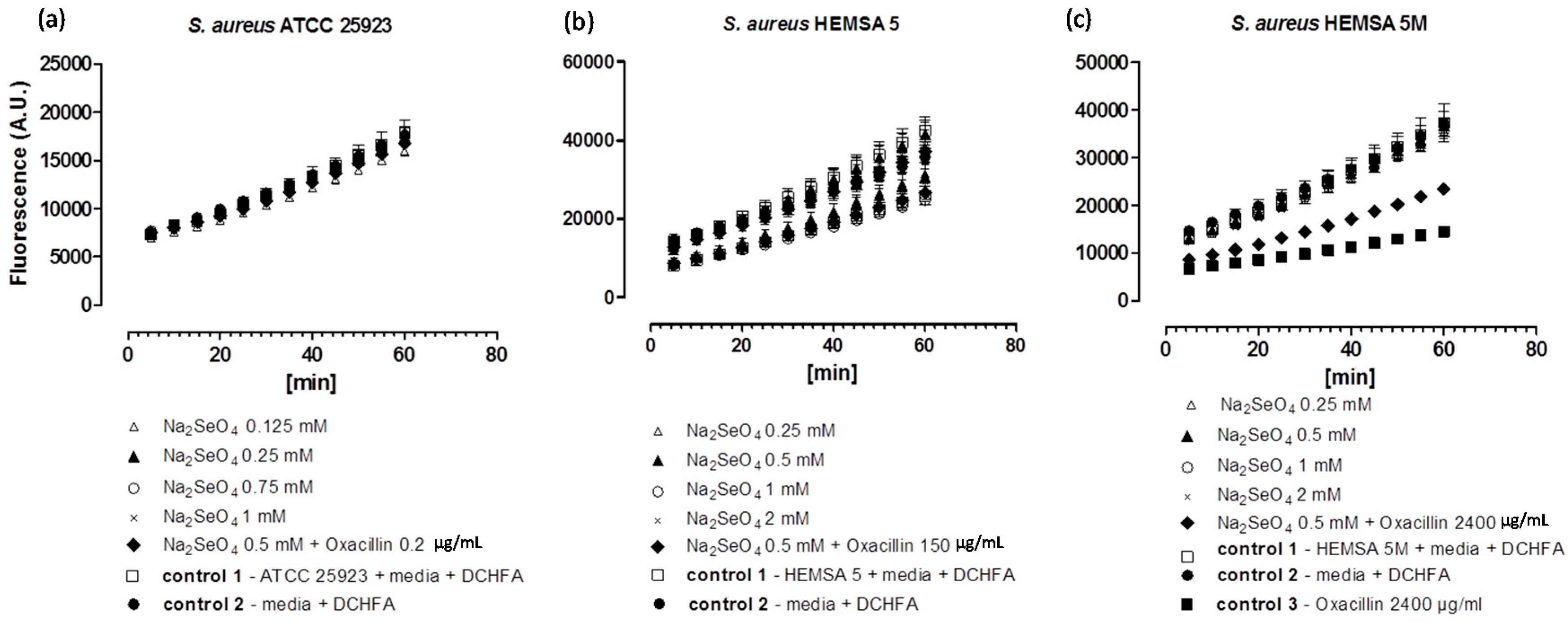
Figure 3.
Generation of intracellular ROS by Na
2SeO
4 in different strains of
S. aureus. ATCC 25923 (
a); HEMSA 5 (
b) and HEMSA 5M (
c). See legend of
Figure 2 for further details.
Figure 3.
Generation of intracellular ROS by Na
2SeO
4 in different strains of
S. aureus. ATCC 25923 (
a); HEMSA 5 (
b) and HEMSA 5M (
c). See legend of
Figure 2 for further details.
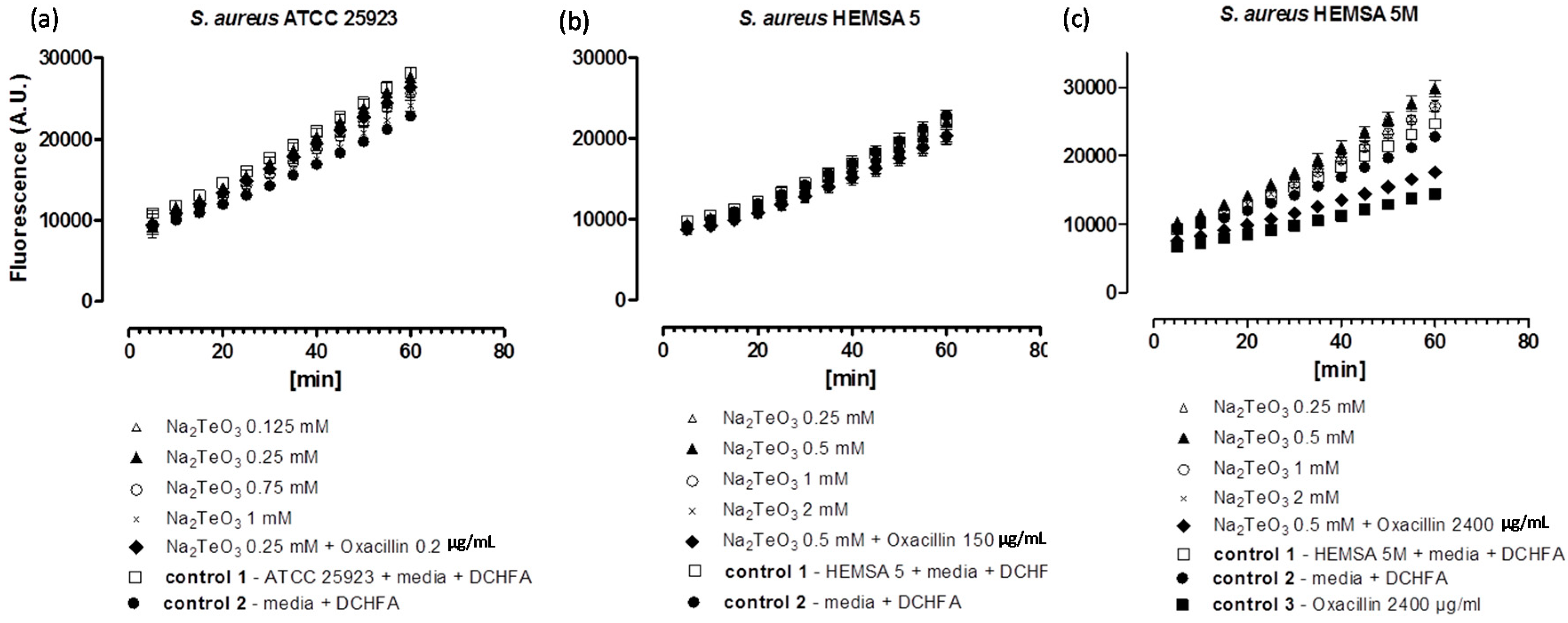
Figure 4.
Generation of intracellular ROS by Na
2TeO
3 in different strains of
S. aureus. ATCC 25923 (
a); HEMSA 5 (
b) and HEMSA 5M (
c). See legend of
Figure 2 for further details.
Figure 4.
Generation of intracellular ROS by Na
2TeO
3 in different strains of
S. aureus. ATCC 25923 (
a); HEMSA 5 (
b) and HEMSA 5M (
c). See legend of
Figure 2 for further details.
Despite the fact that some of the salts notably increased the efficiency of some of the antibiotics, none of the inorganic selenium or tellurium salts studied, either on its own or in combination with oxacillin, increased the intracellular levels of ROS significantly. This finding is truly unexpected, as compounds such as selenite are known to generate ROS
in vitro [
15,
16,
17], are able to modify the thiol redox state and cellular thiolstat, and also have the potential to inhibit enzymes involved in the reduction of oxidative stress. Selenite even promotes DNA strand breaks in yeast, possibly also via a redox mechanism [
3,
18,
19,
20]. As already mentioned above, there are also reports that K
2TeO
3 causes a rather strong increase in intracellular ROS levels in Gram-negative
E. coli [
21]. Hence, at least in theory, one may have expected an increase in ROS levels for Na
2TeO
3 and Na
2SeO
3, either by the direct chemical generation of additional ROS, by damaging cellular components subsequently leading to an indirect increase of ROS concentrations, or by the inhibition of ROS removal systems. Still, it seems that at the concentrations used, none of these redox events plays any major role in the bacteria studied here.
Noteworthy, there is also no consistent evidence of any “antioxidant” activity of Na
2TeO
3, Na
2SeO
3 and Na
2SeO
4 in the bacteria investigated. Generally, no significant differences in fluorescence in the presence of the compound and the controls were observed. Only in one case,
i.e., when the HEMSA 5M strain was treated with oxacillin (2400 µg/mL) (with or without the presence of the salts examined), could a significant decrease (
p < 0.005) of fluorescence compared to the control be observed (
Figure 2c,
Figure 3c and
Figure 4c). This observed decrease in fluorescence, however, could be due to the high concentration of oxacillin used in the ROS assay of this strain, which may have influenced the natural conversion of DCHFA into DCFA. Indeed, a specific control (
i.e., control 3) has been used in those cases to confirm that oxacillin, and not the selenium or tellurium salts, is probably responsible for this particular observation.
Eventually, one must also emphasize that the DCHFA-assay is widely used yet far from perfect: it does not capture all ROS and its outcome is also affected by the activity of certain cellular esterases (see also
Section 3). Furthermore, Kalyanaraman has suggested that the intracellular oxidation of this dye may be mediated to some extent by iron or by cytochrome
c [
22]. Hence any results obtained by this fluorescent assay are rough estimates only and need to be treated with some caution. Whilst the DCHFA assay is still widely accepted as a good measure of intracellular levels of ROS, a more sensible,
i.e., sensitive and selective method should be used in future studies to confirm the lack of pro-oxidant activity of the salts tested.
More or less by coincidence, however, we have observed a rather different, perhaps not entirely expected biological redox activity associated with some of these salts, which may well explain some of the findings shown in
Table 2 and which may also provide a new angle to the discussion. Upon exposure to tellurite and selenite salts, the strains of bacteria investigated changed color, and in fact generated and then obviously accumulated solid red and black deposits, which based on previous reports appear to consist of elemental selenium and elemental tellurium, respectively. Indeed, a closer look at the literature on bacteria dealing with selenium and tellurium salts shows that this finding is not wholly unanticipated. Many bacteria, including
Halococcus salifodinae BK18,
Azospirillum brasilense,
Sulfurospirillum and
Thauera selenatis and also some more common ones such as
Pseudomonas,
Lactobacillus and
Bacillus cereus are known to reduce selenite to elemental selenium (nano)particles [
23,
24,
25,
26,
27,
28,
29].
Thauera selenatis is even able to reduce selenate to form elemental selenium deposits. In fact, different species of
Lactobacillus are used as a natural source of selenium nanoparticles, for instance to enrich probiotics with selenium [
30]. Similarly, bacteria such as
Bacillus beveridgei,
Rhodobacter capsulatus and even
E. coli readily reduce Na
2TeO
3 to generate elemental tellurium, usually in form of nanoscopic or microscopic rods and needles [
31,
32,
33]. Such a reactive cascade leading to elemental selenium or tellurium is shown in
Figure 5. It should be noted that to the best of our knowledge, there have been no reports so far that
Staphylococcus converts selenite or tellurite to elemental selenium or tellurium, respectively.
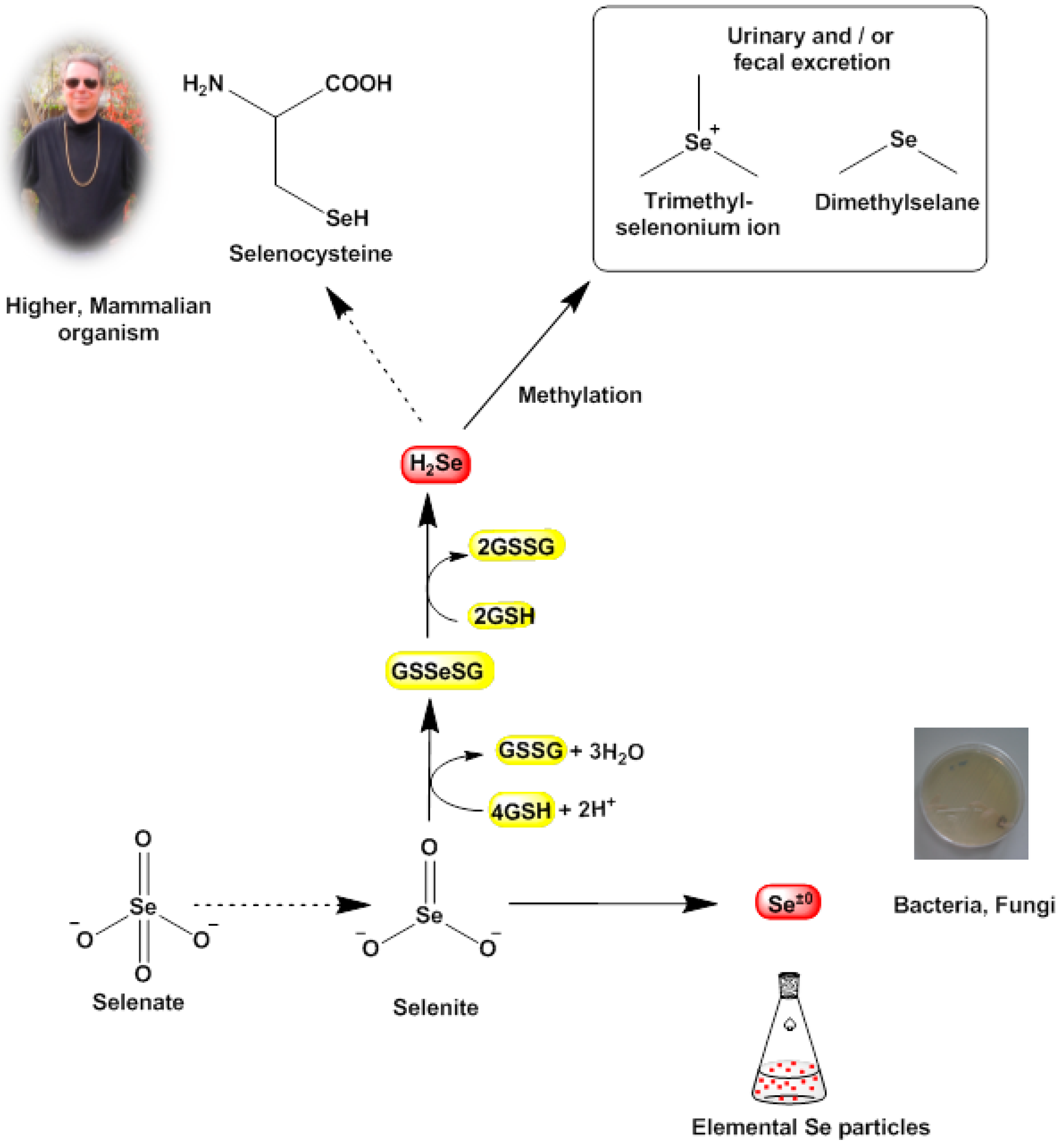
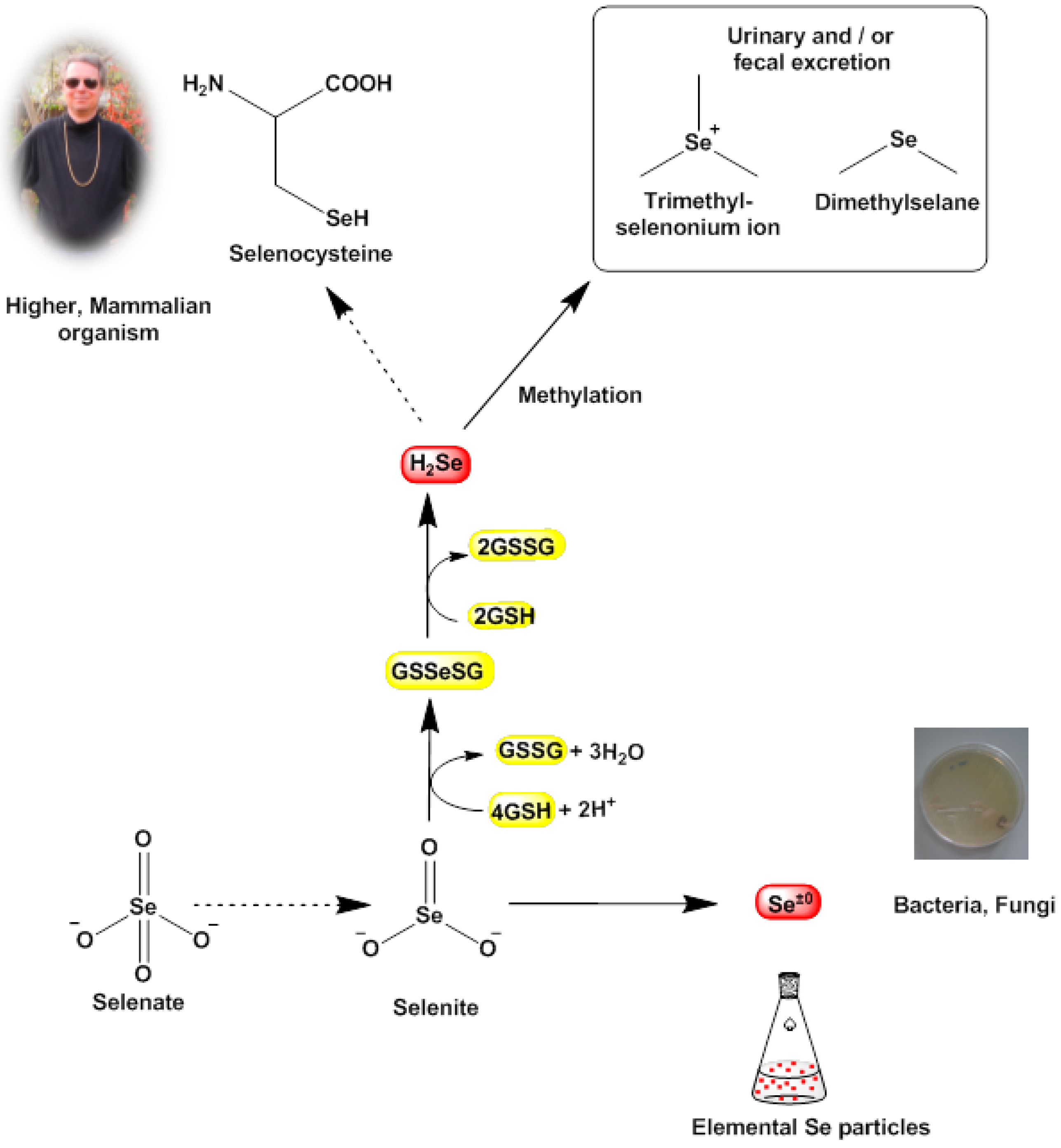
Indeed, the elemental precipitates formed not only stain the bacterial cells red or black, those particles also exert a massive mechanical stress on the bacterial cell which may ultimately cause cell death via a combination of mechanical stresses and unfavorable cellular responses toward them. Eventually, the bioreductive formation of these solid deposits in the bacterial cell may weaken the cell and hence may also explain—at least in part—the notable efficiency of traditional antibiotics in such compromised cells. This process may also contribute to the selective action of such simple chalcogen salts, as mammalian cells do not reduce selenite to elemental selenium. Here, selenite is mostly “sequestered” chemically by glutathione (GSH) in form of glutathione selenotrisulfide (GSSeSG). The latter is reduced further to hydrogen selenide (H
2Se) which either serves as a selenium source for the synthesis of selenocysteine or is methylated to dimethylselenide (CH
3SeCH
3) and ionic trimethylselenonium ((CH
3)
3Se
+) and excreted (
Figure 5) [
34,
35]. In any case, mammalian cells normally do not form solid selenium or tellurium particles and hence are not affected by the mechanical stresses and toxicity caused by such particles [
36].
Since the reduction of salts such as Na
2SeO
3 often proceeds spontaneously in the presence of thiols (such as GSH), another mode of (cytotoxic) action has been discussed for various selenium and tellurium compounds [
37]. The underlying chemical mechanism includes the—probably spontaneous and more or less random—reaction of such compounds with thiol groups in proteins. Such reactions result in various posttranslational oxidative modifications which may also impact on the function and activity of the proteins and enzymes affected. Since many cysteine proteins and enzymes are involved in pivotal cellular processes (metabolism, signaling, maintenance of redox homeostasis
etc.), and hence have recently been subsumed under the notion of the “cellular thiolstat”, a modification of their cysteine residues and thus function and activity may have serious consequences for the cell [
10].
Figure 5.
Known bioreductive pathways for SeO32− (and TeO32−) in certain bacterial cells. These pathways often terminate in the formation of (solid) elemental selenium and tellurium (nano)particles, which subsequently may cause damage to the cell. In our studies, it appears that selenite (SeO32−) and tellurite (TeO32−) are also reduced by S. aureus, as different reports have observed similar changes in color and the formation of deposits and have linked these physical changes to the formation of selenium and tellurium, respectively. In sharp contrast, mammalian cells tend to by-pass the “zero oxidation state” and form H2Se as a valuable raw product for the synthesis of the amino acid selenocysteine, whilst excess selenium is excreted in form of methylated products. Hence the bioreductively generated toxicity of selenium and tellurium (nano)particles is probably more or less specific for lower organisms.
Figure 5.
Known bioreductive pathways for SeO32− (and TeO32−) in certain bacterial cells. These pathways often terminate in the formation of (solid) elemental selenium and tellurium (nano)particles, which subsequently may cause damage to the cell. In our studies, it appears that selenite (SeO32−) and tellurite (TeO32−) are also reduced by S. aureus, as different reports have observed similar changes in color and the formation of deposits and have linked these physical changes to the formation of selenium and tellurium, respectively. In sharp contrast, mammalian cells tend to by-pass the “zero oxidation state” and form H2Se as a valuable raw product for the synthesis of the amino acid selenocysteine, whilst excess selenium is excreted in form of methylated products. Hence the bioreductively generated toxicity of selenium and tellurium (nano)particles is probably more or less specific for lower organisms.
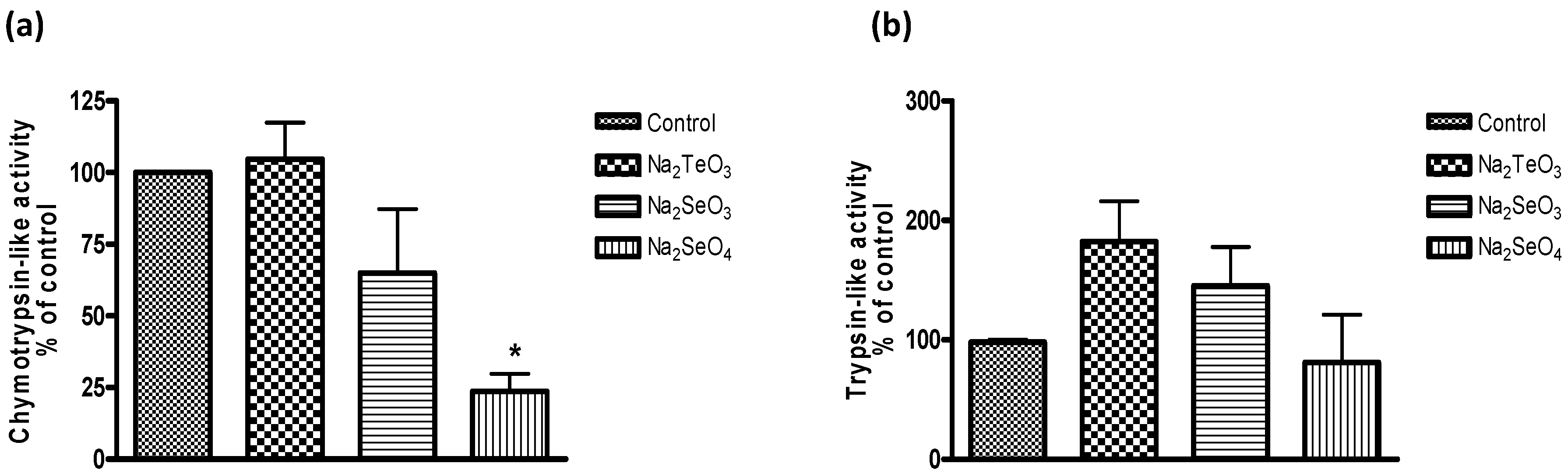
As part of our study, we have therefore considered if such compounds also interact with key cellular components or events, especially those which are related to the cellular thiolstat and whose inhibition may be highly detrimental to the cell. As part of this search for potential protein targets, we noticed a significant inhibition of the proteasome in yeast (
Figure 6). It should be pointed out that
S. cerevisiae was used as model as the yeast proteasome can be studied reliably in isolation as well as in the intact cell. Here, Na
2SeO
3 and especially Na
2SeO
4 were most active. The impact of Na
2SeO
4 on proteasome activity was observed at higher micromolar concentrations and, at a concentration of 1 mM, Na
2SeO
4 reduced the chymotrypsin-like activity of the yeast proteasome in intact cells significantly, to less than 25%.
Arguably, these concentrations of selenate are high and not relevant physiologically. Still, one needs to bear in mind that these experiments were performed with intact cells, and intracellular concentrations of Na
2SeO
4 may be lower. In fact, the known proteasome inhibitor bortezomib, which was used in this part of the study as positive control, failed to inhibit proteasome activity in yeast cells altogether when used at its standard concentration of 1 micromolar. Whilst bortezomib and similar inhibitors penetrate human cells readily, they cannot penetrate wild-type yeast cells due to an inherent impermeability of the cell wall or membranes [
38]. For this reason, yeast has been used simply as a model system, and not as a target for therapeutic intervention.
Notably, both selenium salts did not inhibit the trypsin-like activity and Na
2TeO
3, often considered as the most active of these chalcogen salts, had no inhibitory effect on the yeast proteasome under the conditions used. Indeed, at closer inspection, it even appears that Na
2SeO
3 and Na
2TeO
3 may actually stimulate the trypsin-like activity, although those increases are statistically not significant (
Figure 6b).
Figure 6.
Inhibition of the proteasome in intact cells of S. cerevisiae (chymotrypsin-like activity (a) and trypsin-like activity (b)). Data was obtained from cell extracts after 3 h incubation in the presence of 1000 µM of tellurium and selenium salts. Values represent means with SD bars from at least three independent experiments, and statistical significances were calculated using a one-way ANOVA followed by Bonferroni’s multiple comparison test. * p < 0.05.
Figure 6.
Inhibition of the proteasome in intact cells of S. cerevisiae (chymotrypsin-like activity (a) and trypsin-like activity (b)). Data was obtained from cell extracts after 3 h incubation in the presence of 1000 µM of tellurium and selenium salts. Values represent means with SD bars from at least three independent experiments, and statistical significances were calculated using a one-way ANOVA followed by Bonferroni’s multiple comparison test. * p < 0.05.
Once more, one may speculate that the compounds employed are able to react with thiol groups in proteins, and therefore may also interfere with and inhibit a range of proteins and enzymes, including parts of the proteasome, either by a direct modification of cysteine residues in components of the proteasome itself or by a widespread modification of proteins which then end up and “overload” the proteasome in a detrimental manner. Indeed, the notion that thiol-selective yet otherwise unspecific (re-)agents may be able to modify a wide range of cellular, redox-sensitive cysteine-proteins, which subsequently not only lose their function and activity, but also end up as “garbage” and overload the proteasome, is highly stimulating and deserves a more detailed consideration in the future.
On the one side, these findings are rather instructive, as the proteasome represents an important target in modern therapy. In eukaryotes, the ubiquitin-proteasome system is the main proteolytic system, whilst in bacteria there are different proteolytic complexes that are directly linked to bacterial virulence [
39,
40]. Not surprisingly, the proteolytic machinery is being considered as a promising therapeutic target, especially in multi-resistant bacterial species [
41]. Here, fellutamide B acts as inhibitor of the
Mycobacterium tuberculosis proteasome, whilst an inhibitor of the protease ClpXP increases the sensitivity of methycillin-resistant
S. aureus and
Bacillus anthracis to antibiotics that act on the bacterial wall (synthesis) and membrane [
42,
43]. Indeed, such observations on the “re-sensitization” of the drug resistant strains against conventional antibiotics acting on the cell wall synthesis via an inhibition of the proteasome mirror most of the still preliminary findings of our present study [
39,
40,
41,
42,
43]. It is therefore a highly rewarding task to study the impact of selenium food supplements and organic selenium compounds on different proteolytic systems/proteasomes (e.g., bacteria, fungi, human cancer cells, normal cells) more extensively in the future.
On the other side, the proteasome represents probably just one target for salts such as Na2SeO3 and Na2SeO4. There may be many other—yet unidentified—targets, and some of them may also differ between bacterial, fungal and mammalian cells, depending on the specific components of the respective cellular thiolstat and the metabolic processes in action (such as specific bioreductive pathways).
In any case, the observation that simple salts, such as Na2SeO3, are able to compromise bacterial cells, even employing some redox driven “nanotechnology”, underlines the considerable interest in redox active and catalytic selenium compounds—some of which form part of our daily nutrition—and their potential uses in medicine.
2.3. From Simple Selenium Salts to Redox Modulating Selenium Drugs
Whilst the results obtained so far for Na
2SeO
3 and Na
2SeO
4 (as well as for Na
2TeO
3) are interesting, such compounds are not particularly useful from a pharmaceutical point of view, not least because of the high concentrations required for a significant activity. As already mentioned, we have therefore investigated some of the most interesting findings obtained as part of this study further, this time employing some of our most potent redox modulating selenium and tellurium agents (compounds
1–
4 in
Figure 1) [
5,
7,
44,
45]. Since these compounds have been designed as potential therapeutic agents (for instance considering reactivity, stability, pharmacokinetics, Lipinski’s “Rule of Five”,
etc.), it was hardly surprising to find a pronounced antibacterial activity for the selenium compound
1 against
S. aureus ATCC25923 with MIC values as low as 31.25 µM. Interestingly, the two tellurium compounds
3 and
4 were even rather active against the MRSA strain HEMSA 5, with MIC values of 62.5 µM and 31.25 µM, respectively. These MIC values are rather low, also when compared to the MIC values listed for some of the conventional antibiotics in
Table 1. Compound
4, which contains two tellurium centers and a quinone moiety, therefore seems to be particularly promising as a potential antibacterial agent, and on its own [
46].
When considered in combination with traditional antibiotics, such as oxacillin, these compounds did not enhance the activity of the antibiotics used. Yet again, this is hardly surprising. The special bioreductive “chemistry” witnessed in bacteria in the case of salts such as Na2SeO3, Na2SeO4 and Na2TeO3, for instance, is notably absent for compounds 1–4. First and foremost, no color changes were observed when the organoselenium and organotellurium compounds were used. Secondly, there was also no evidence of any deposit formation when the cells were incubated with the organic selenium and tellurium compounds. Hence based on this evidence, it appears that no elemental selenium and tellurium particles similar to the ones observed for the salts were formed and that the kind of mechanical and (bio)chemical stresses discussed for compounds such as Na2SeO3 and Na2TeO3 was therefore almost certainly also absent.
Besides simple changes in color, these differences in “chemistry” between the inorganic and organic selenium and tellurium compounds obviously translate into further differences in biochemical activity. The organic compounds employed contain several redox active, catalytic centers and are known to interfere with the intracellular redox homeostasis in human cells. As part of a related study, it has moreover become apparent that such compounds, especially compounds
2 and
4, are able to interfere with proteasomes in intact cells as well as with isolated human proteasomes, where those two compounds at a concentration of just 1 µM reduce chymotrypsin-like activity by 80% and trypsin-like activity by 90%. We also know from our previous studies that these compounds inhibit mammalian thioredoxin reductase (TrxR)
in vitro, and at rather low concentrations. The selenium compound
2, at just 1 µM, reduces TrxR activity to less than 50%, whilst the tellurium compound
3 is even more active, with just 60% of the original TrxR activity remaining when used at 500 nM and less than 20% TrxR activity remaining when used at an 1 µM concentration [
47]. There is also evidence that selenium compounds closely related chemically to
1 and
2 interfere with protein synthetic pathways and ultimately protein synthesis [
47].


 ,
, 

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}