
Inhibition of Tanshinone IIA, Salvianolic Acid A and Salvianolic Acid B on Areca Nut Extract-Induced Oral Submucous Fibrosis in Vitro

 ,
,
Abstract
:1. Introduction
2. Results and Discussion
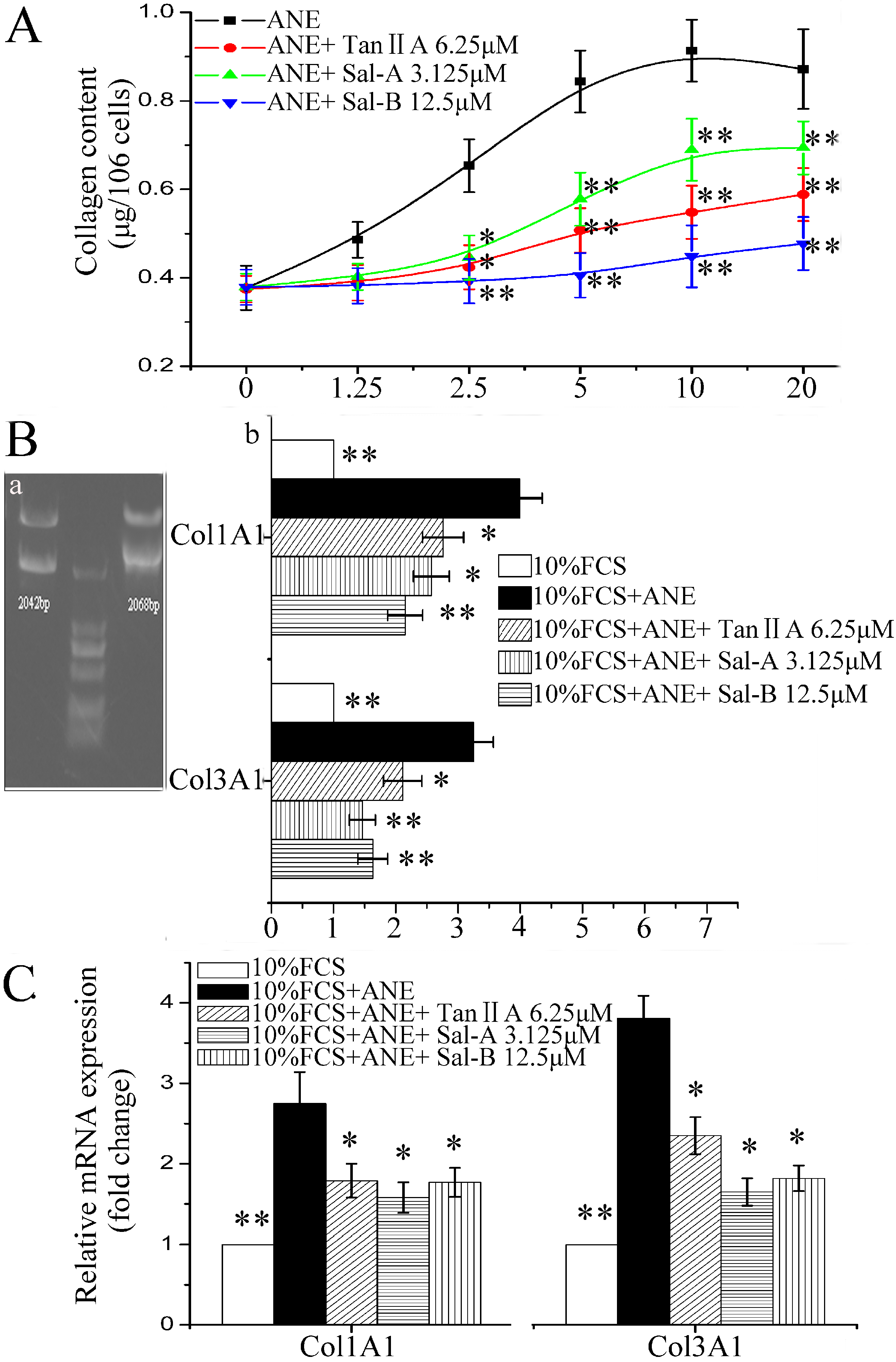
2.1. Tan-IIA1, Sal-A and Sal-B Ameliorated the ANE-Induced Abnormal Viability of MOMFs and Inhibited Collagen Accumulation
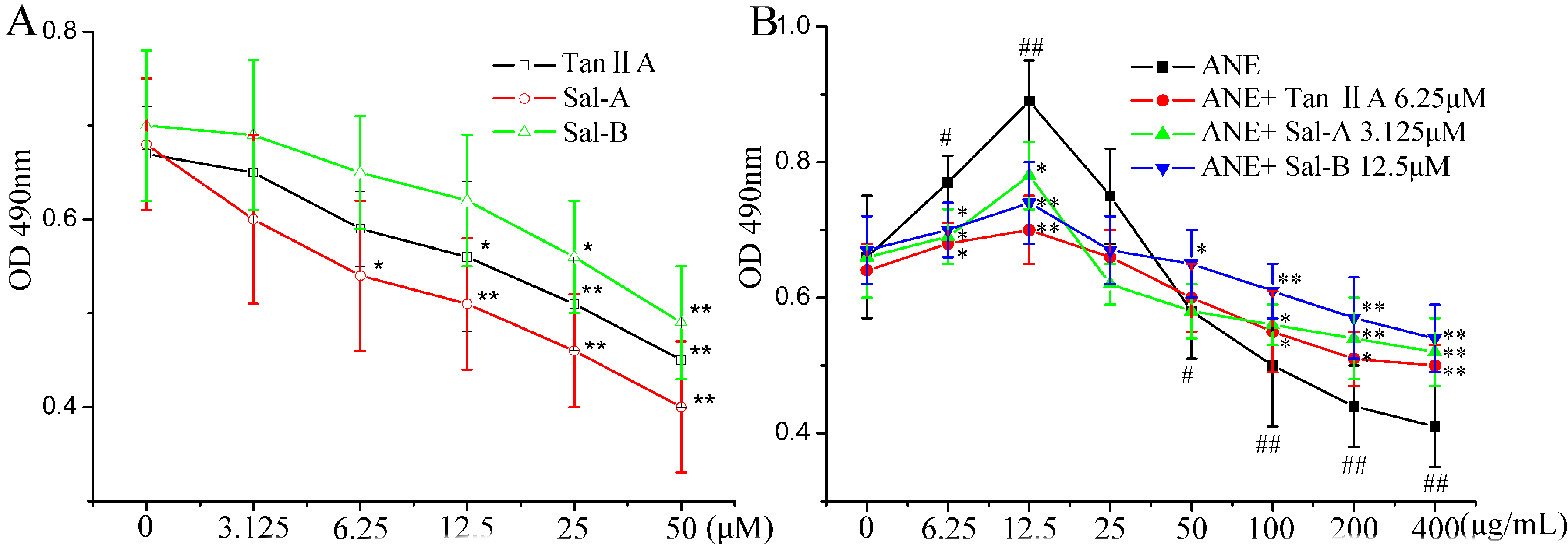


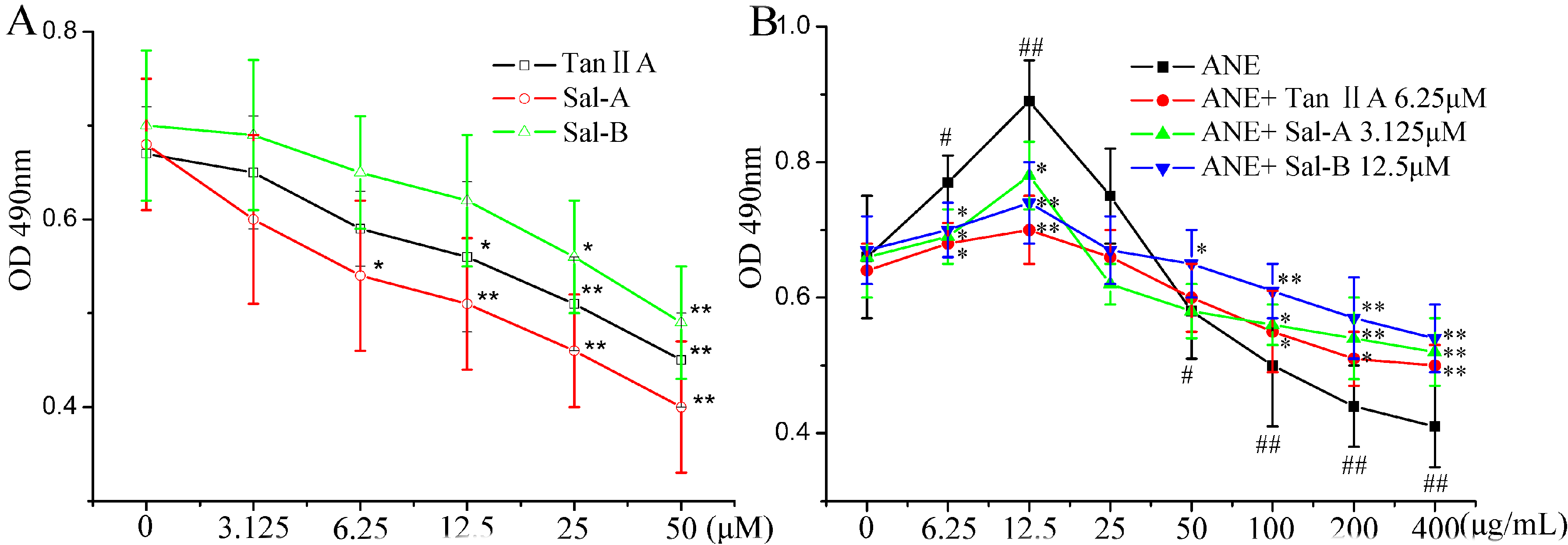{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Forward | Reverse |
|---|---|---|
| COL1A1 | 5'-GTGCTAAAGGTGCCAATGGT-3' | 5'-ACCAGGTTCACCGCTGTTAC-3' |
| COL3A1 | 5'-AGGGGAGCTGGCTACTTCT-3' | 5'-CCTCCTTCAACAGCTTCCTG-3' |
| MMP-1 | 5'-CTTCTTCTTGTTGAGCTGGACTC-3' | 5'-CTGTGGAGGTCACTGTAGACT-3' |
| MMP-13 | 5'-CTTCTTCTTGTTGAGCTGGACTC-3' | 5'-CTGTGGAGGTCACTGTAGACT-3' |
| TIMP1 | 5'-TGACATCCGGTTCGTCTACA-3' | 5'-TGCAGTTTTCCAGCAATGAG-3' |
| TIMP2 | 5'-AGTTCTTCGCCTGCATCAAG-3' | 5'-GTCGAGAAACTCCTGCTTGG-3' |
| CTGF | 5'-TCAACCTCAGACACTGGTTTCG-3' | 5'-TAGAGCAGGTCTGTCTGCAAGC-3' |
| TGF-β1 | 5'-AAACGGAAGCGCATCGAA-3' | 5'-GGGACTGGCGAGCCTTAGTT-3' |
| IL-6 | 5'-TTCCATCCAGTTGCCTTCTT-3' | 5'-ATTTCCACGATTTCCCAGAG-3' |
| IL-1 | 5'-CCCAAGCAATACCCAAAGAA-3' | 5'-CATCAGAGGCAAGGAGGAAA-3' |
| TNF-a | 5'-CAAATGGCCTCCCTCTCAT-3' | 5'-TGGGCTACAGGCTTGTCACT-3' |
| β-actin | 5'-GGAGACAACCTGGTCCTCAG-3' | 5'-ACCCAGAAGACTGTGGATGG-3' |
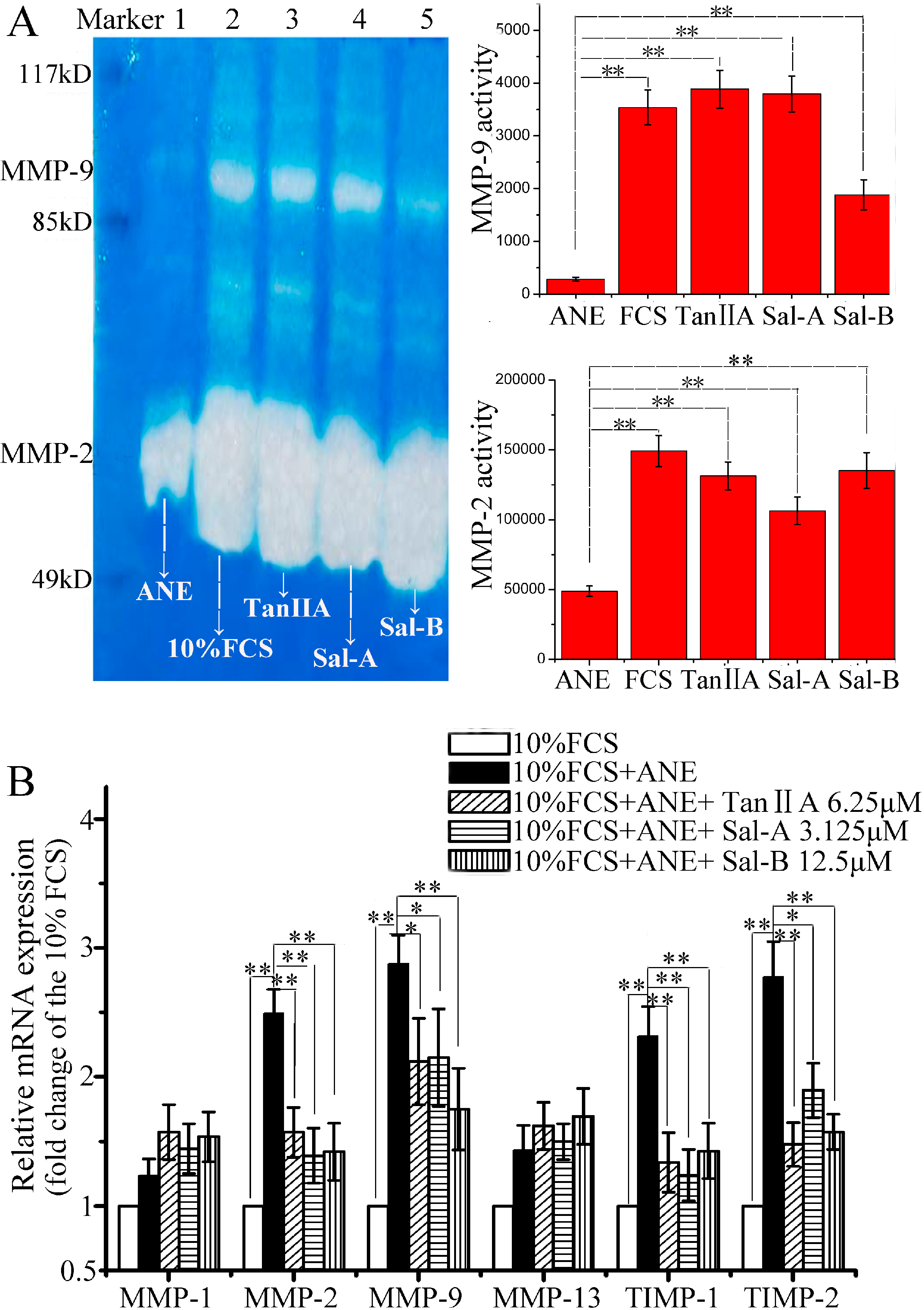
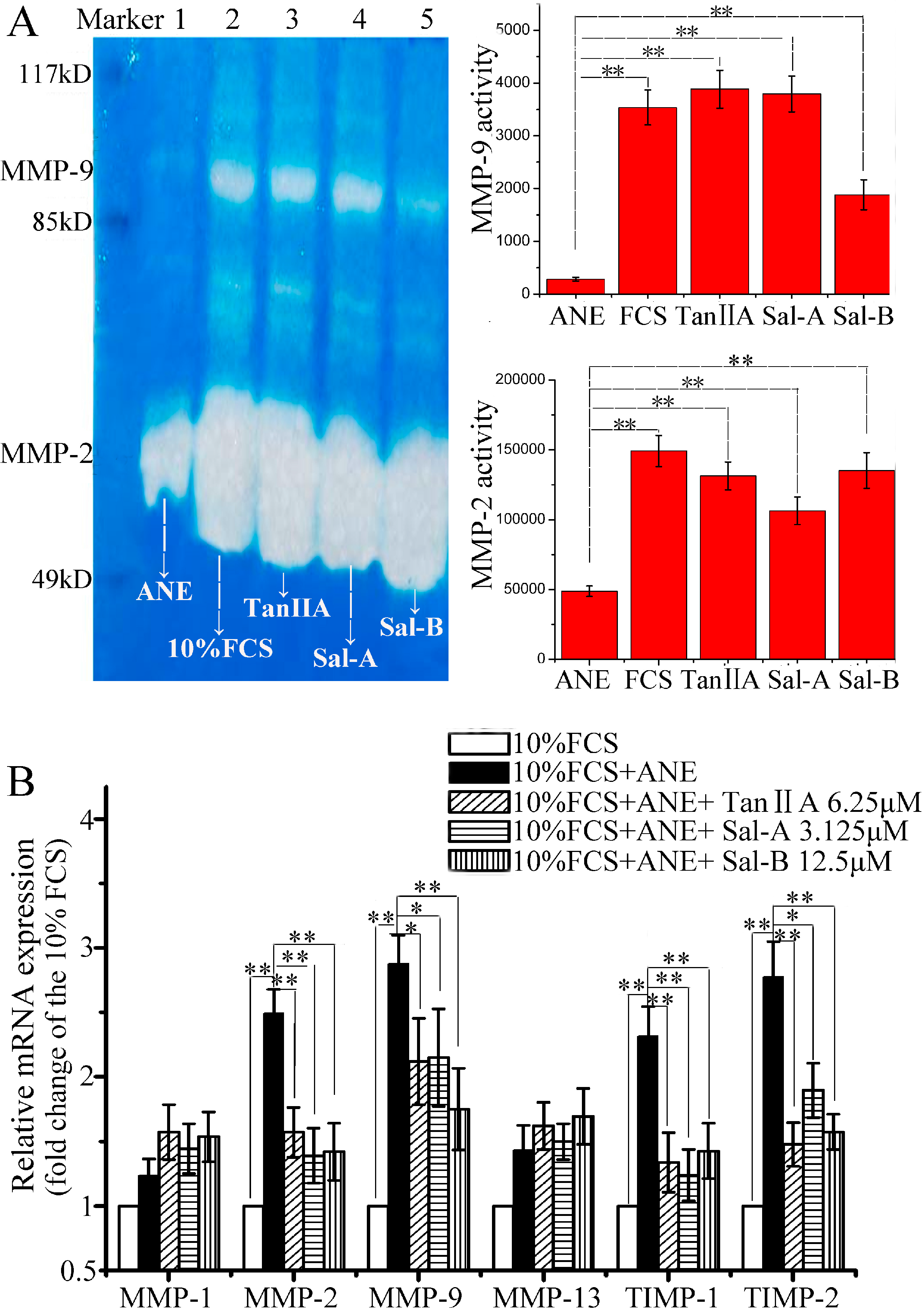
2.2. Tan-IIA1, Sal-A and Sal-B Improved MMP-2/-9 Activity and Inhibited TIMP-1/-2 Expression after ANE Treatment
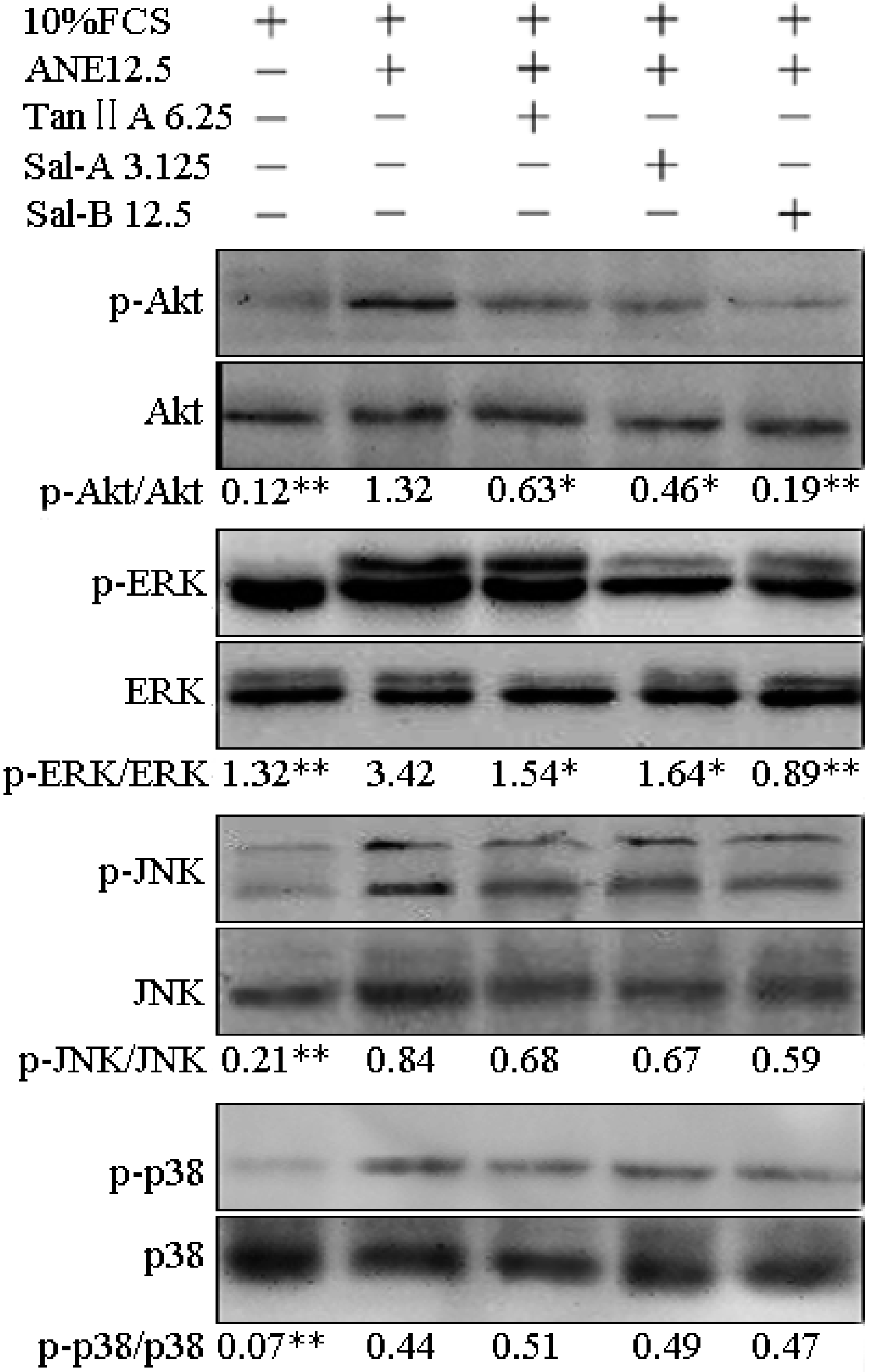
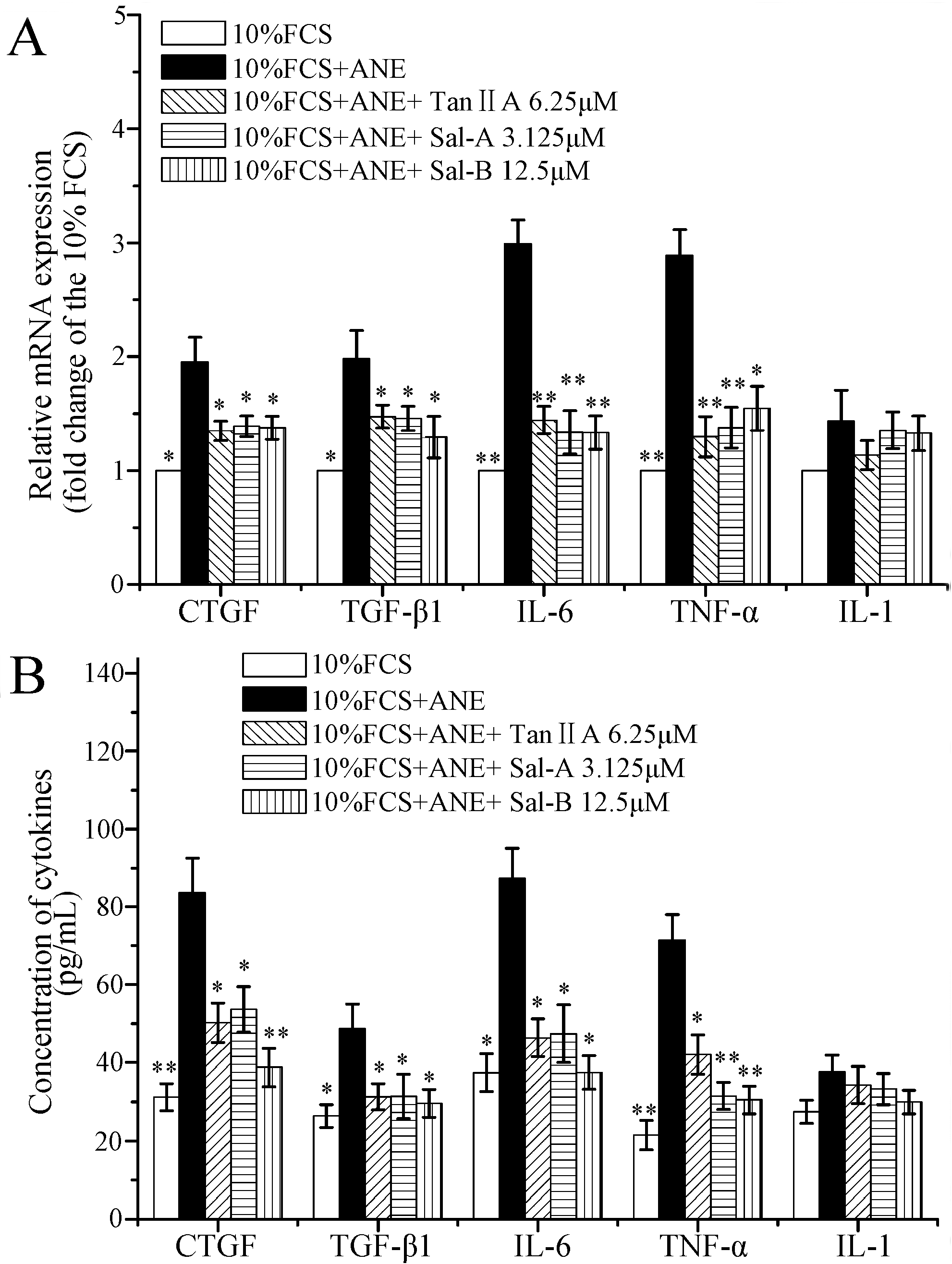
2.3. Tan-IIA1, Sal-A and Sal-B Inhibited the Activation of AKT and ERK MAPK Pathways and Influenced the Release of Cytokines in MOMFs after ANE Treatment
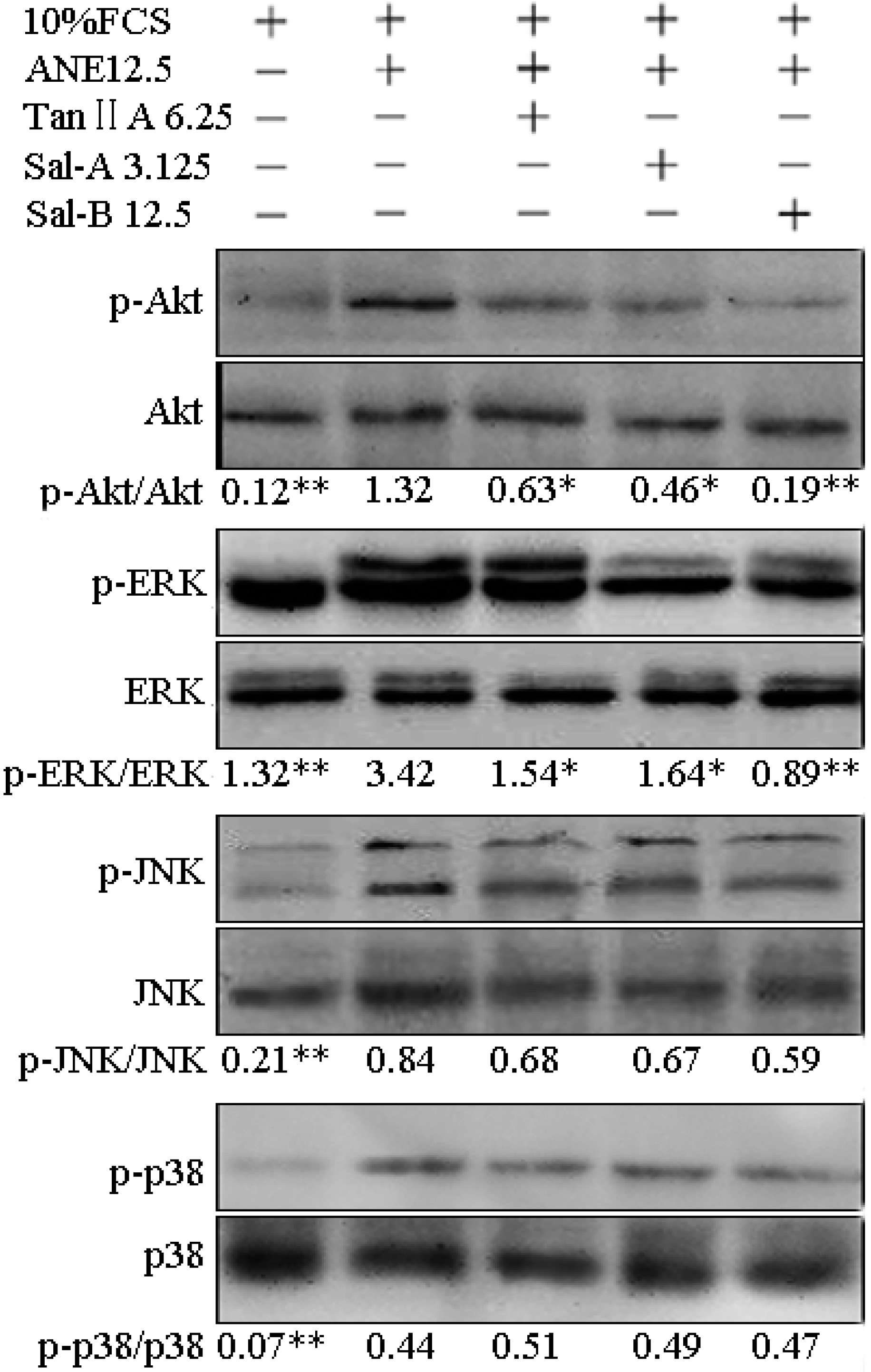
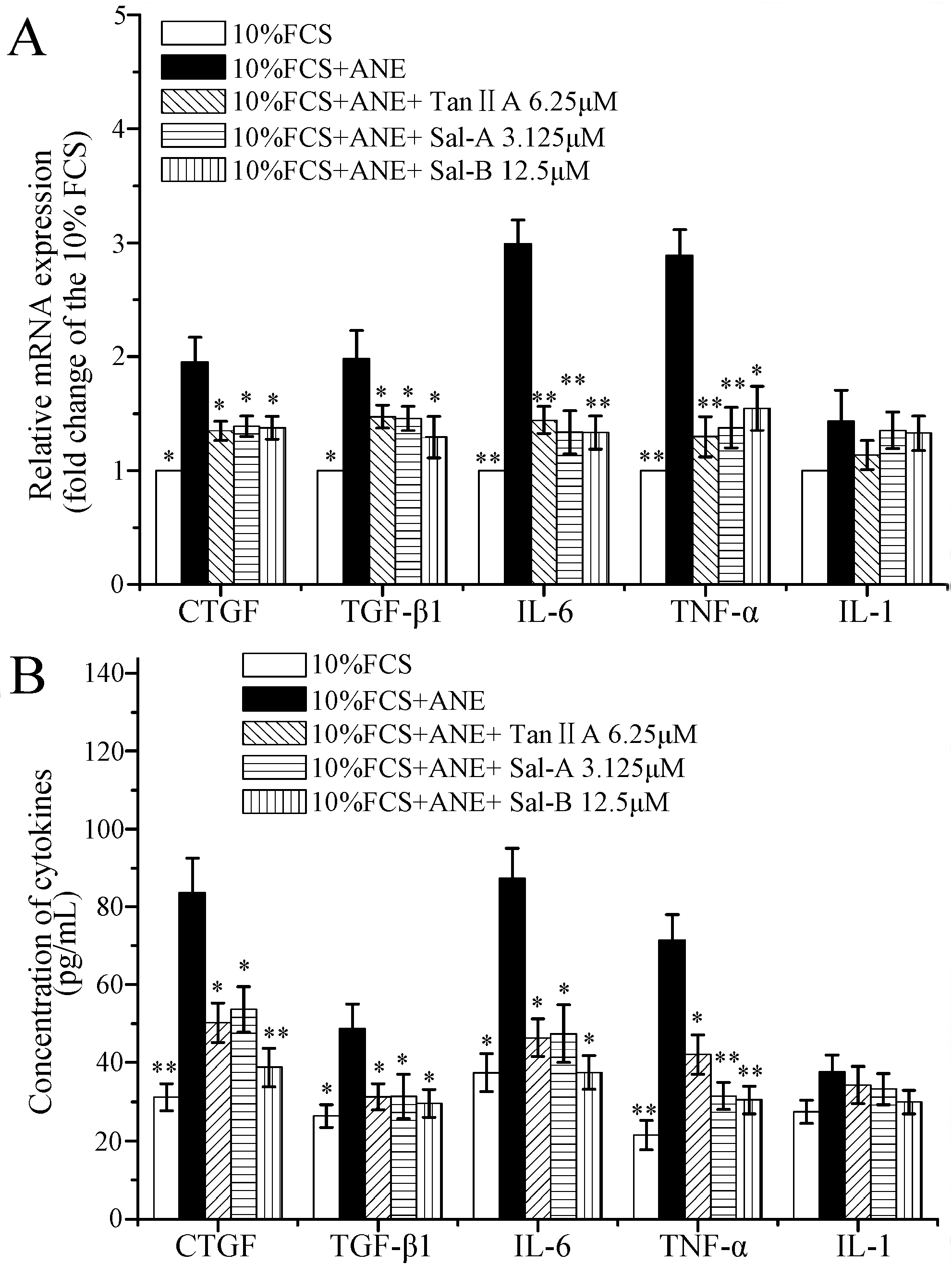
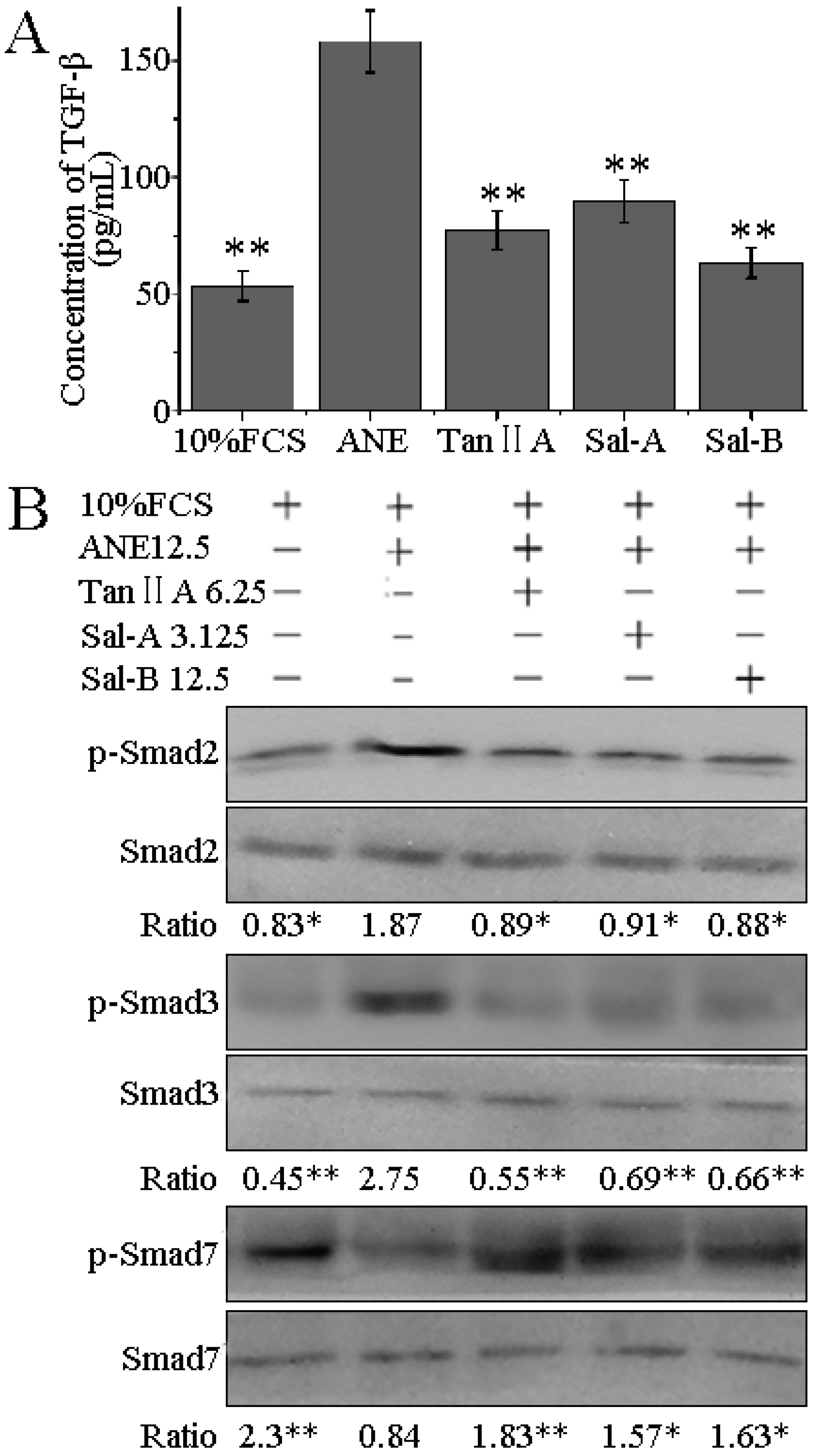
2.4. Tan-IIA1, Sal-A and Sal-B Inhibited the Release of TGF-β and Activation of TGF-β/Smads Pathway in HaCaT Cells
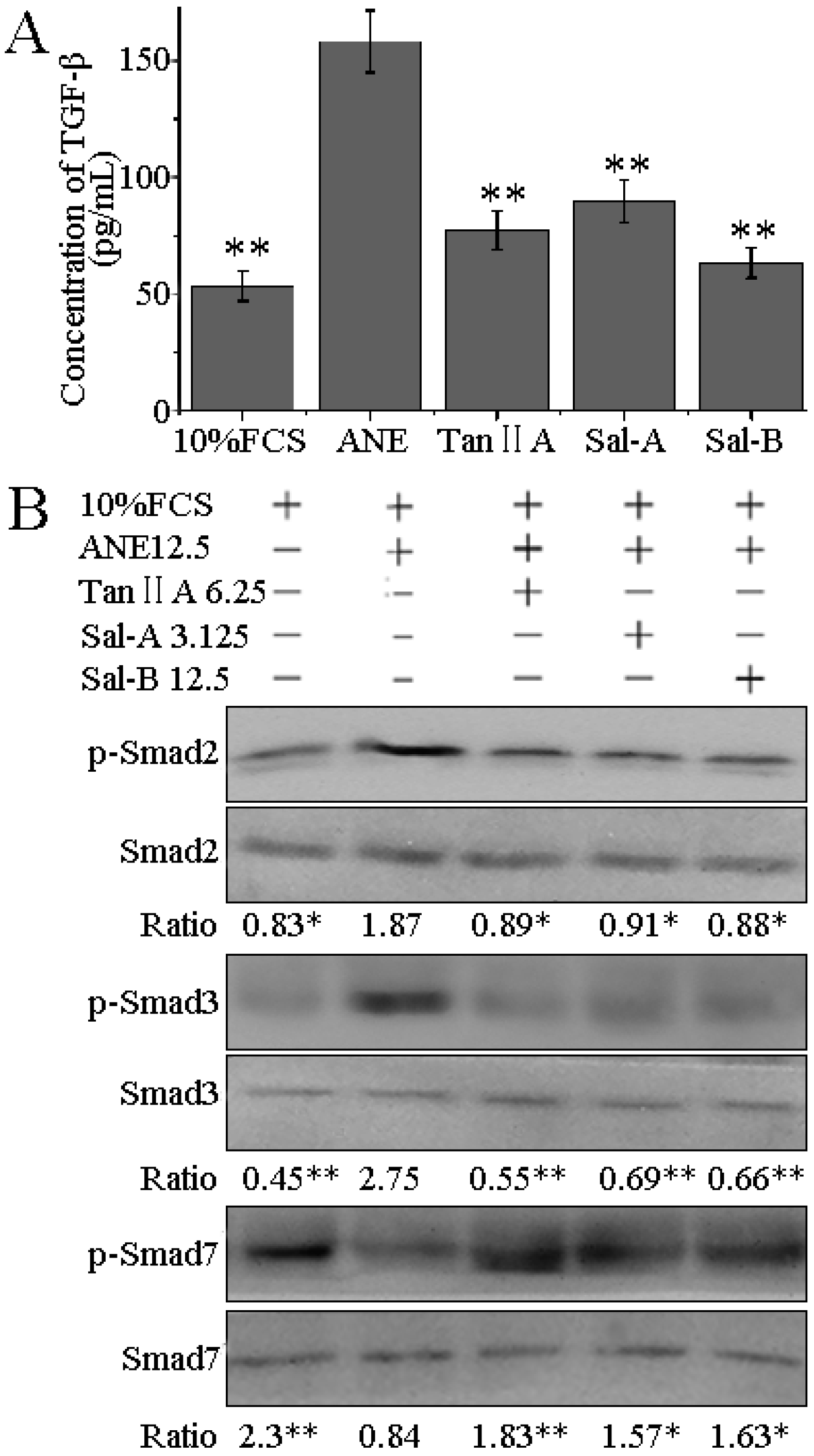
3. Experimental Section
3.1. Material
3.2. Cell Culture and MTT Assay
3.3. Plasmid Construction, Transfection and Luciferase Assay
3.4. Hydroxyproline Assay
3.5. Gelatin Zymography Assay
3.6. qRT-PCR Assay
3.7. ELISA Assay
3.8. Western Blot Assay
3.9. Statistical Analysis
4. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- IARC. Betel-quid and areca-nut chewing and some areca-nut-derived nitrosamines—Summary of data reported and evaluation. IARC Monogr. Eval. Carcinog. Risks Hum. 2004, 85, 1–334. [Google Scholar]
- Tilakaratne, W.M.; Klinikowski, M.F.; Saku, T.; Peters, T.J.; Warnakulasuriya, S. Oral submucous fibrosis: Review on aetiology and pathogenesis. Oral Oncol. 2006, 42, 561–568. [Google Scholar]
- Utsunomiya, H.; Tilakaratne, W.M.; Oshiro, K.; Maruyama, S.; Suzuki, M.; Ida-Yonemochi, H.; Cheng, J.; Saku, T. Extracellular matrix remodeling in oral submucous fibrosis: Its stage-specific modes revealed by immunohistochemistry and in situ hybridization. J. Oral Pathol. Med. 2005, 34, 498–507. [Google Scholar]
- Rajalalitha, P.; Vali, S. Molecular pathogenesis of oral submucous fibrosis—A collagen metabolic disorder. J. Oral Pathol. Med. 2005, 34, 321–328. [Google Scholar]
- Dai, J.P.; Chen, J.; Bei, Y.F.; Han, B.X.; Wang, S. Influence of borneol on primary mice oral fibroblasts: A penetration enhancer may be used in oral submucous fibrosis. J. Oral Pathol. Med. 2009, 38, 276–281. [Google Scholar]
- Moutasim, K.A.; Jenei, V.; Sapienza, K.; Marsh, D.; Weinreb, P.H.; Violette, S.M.; Lewis, M.P.; Marshall, J.F.; Fortune, F.; Tilakaratne, W.M.; et al. Betel-derived alkaloid up-regulates keratinocyte alphavbeta6 integrin expression and promotes oral submucous fibrosis. J. Pathol. 2011, 223, 366–377. [Google Scholar]
- Tsai, C.H.; Yang, S.F.; Lee, S.S.; Chang, Y.C. Augmented heme oxygenase-1 expression in areca quid chewing-associated oral submucous fibrosis. Oral Dis. 2009, 15, 281–286. [Google Scholar]
- Khan, I.; Kumar, N.; Pant, I.; Narra, S.; Kondaiah, P. Activation of tgf-beta pathway by areca nut constituents: A possible cause of oral submucous fibrosis. PLoS ONE 2012, 7, e51806. [Google Scholar]
- Lu, H.H.; Liu, C.J.; Liu, T.Y.; Kao, S.Y.; Lin, S.C.; Chang, K.W. Areca-treated fibroblasts enhance tumorigenesis of oral epithelial cells. J. Dent. Res. 2008, 87, 1069–1074. [Google Scholar]
- Lee, S.S.; Tsai, C.H.; Ho, Y.C.; Yu, C.C.; Chang, Y.C. Heat shock protein 27 expression in areca quid chewing-associated oral squamous cell carcinomas. Oral Dis. 2012, 18, 713–719. [Google Scholar]
- Lin, S.C.; Lu, S.Y.; Lee, S.Y.; Lin, C.Y.; Chen, C.H.; Chang, K.W. Areca (betel) nut extract activates mitogen-activated protein kinases and nf-kappab in oral keratinocytes. Int. J. Cancer 2005, 116, 526–535. [Google Scholar]
- Yang, S.C.; Lin, S.C.; Chiang, W.F.; Yen, C.Y.; Lin, C.H.; Liu, S.Y. Areca nut extract treatment elicits the fibroblastoid morphological changes, actin re-organization and signaling activation in oral keratinocytes. J. Oral Pathol. Med. 2003, 32, 600–605. [Google Scholar]
- Tseng, Y.H.; Chang, C.S.; Liu, T.Y.; Kao, S.Y.; Chang, K.W.; Lin, S.C. Areca nut extract treatment down-regulates involucrin in normal human oral keratinocyte through p13k/akt activation. Oral Oncol. 2007, 43, 670–679. [Google Scholar]
- Dai, J.; Chen, J.; Han, B.; Bei, Y.; Zhou, X. Regulation of calculus bovis on the function of mice oral fibroblasts. Sheng Wu Gong Cheng Xue Bao 2009, 25, 448–451. [Google Scholar]
- Ma, S.; Yang, D.; Wang, K.; Tang, B.; Li, D.; Yang, Y. Cryptotanshinone attenuates isoprenaline-induced cardiac fibrosis in mice associated with upregulation and activation of matrix metalloproteinase-2. Mol. Med. Rep. 2012, 6, 145–150. [Google Scholar]
- Zhang, Y.; Zhang, Y.; Xie, Y.; Gao, Y.; Ma, J.; Yuan, J.; Li, J.; Wang, J.; Li, L.; Zhang, J.; et al. Multitargeted inhibition of hepatic fibrosis in chronic iron-overloaded mice by salvia miltiorrhiza. J. Ethnopharmacol. 2013, 148, 671–681. [Google Scholar]
- Pan, T.L.; Wang, P.W. Explore the molecular mechanism of apoptosis induced by tanshinone iia on activated rat hepatic stellate cells. Evid. Based Complement. Altern. 2012, 2012, 734987. [Google Scholar]
- Won, S.H.; Lee, H.J.; Jeong, S.J.; Lee, H.J.; Lee, E.O.; Jung, D.B.; Shin, J.M.; Kwon, T.R.; Yun, S.M.; Lee, M.H.; et al. Tanshinone iia induces mitochondria dependent apoptosis in prostate cancer cells in association with an inhibition of phosphoinositide 3-kinase/akt pathway. Biol. Pharm. Bull. 2010, 33, 1828–1834. [Google Scholar]
- Zhou, Z.T.; Yang, Y.; Ge, J.P. The preventive effect of salvianolic acid b on malignant transformation of dmba-induced oral premalignant lesion in hamsters. Carcinogenesis 2006, 27, 826–832. [Google Scholar]
- Wang, R.; Yu, X.Y.; Guo, Z.Y.; Wang, Y.J.; Wu, Y.; Yuan, Y.F. Inhibitory effects of salvianolic acid b on ccl(4)-induced hepatic fibrosis through regulating nf-kappab/ikappabalpha signaling. J. Ethnopharmacol. 2012, 144, 592–598. [Google Scholar]
- Jiang, B.; Li, D.; Deng, Y.; Teng, F.; Chen, J.; Xue, S.; Kong, X.; Luo, C.; Shen, X.; Jiang, H.; et al. Salvianolic acid a, a novel matrix metalloproteinase-9 inhibitor, prevents cardiac remodeling in spontaneously hypertensive rats. PLoS ONE 2013, 8, e59621. [Google Scholar]
- Zhang, H.A.; Gao, M.; Zhang, L.; Zhao, Y.; Shi, L.L.; Chen, B.N.; Wang, Y.H.; Wang, S.B.; Du, G.H. Salvianolic acid a protects human sh-sy5y neuroblastoma cells against h(2)o(2)-induced injury by increasing stress tolerance ability. Biochem. Biophys. Res. Commun. 2012, 421, 479–483. [Google Scholar]
- Lv, Z.; Xu, L. Salvianolic acid b inhibits erk and p38 mapk signaling in tgf-beta1-stimulated human hepatic stellate cell line (lx-2) via distinct pathways. Evid. Based Complement. Altern. 2012, 2012. [Google Scholar] [CrossRef]
- Lu, B.; Ye, Z.; Deng, Y.; Wu, H.; Feng, J. Mek/erk pathway mediates cytoprotection of salvianolic acid b against oxidative stress-induced apoptosis in rat bone marrow stem cells. Cell Biol. Int. 2010, 34, 1063–1068. [Google Scholar]
- Wu, H.T.; Ko, S.Y.; Fong, J.H.; Chang, K.W.; Liu, T.Y.; Kao, S.Y. Expression of phosphorylated akt in oral carcinogenesis and its induction by nicotine and alkaline stimulation. J. Oral Pathol. Med. 2009, 38, 206–213. [Google Scholar]
- Dai, J.P.; Chen, X.X.; Zhu, D.X.; Wan, Q.Y.; Chen, C.; Wang, G.F.; Li, W.Z.; Li, K.S. Panax notoginseng saponins inhibit areca nut extract-induced oral submucous fibrosis in vitro. J. Oral Pathol. Med. 2014, 43, 464–470. [Google Scholar]
- Chang, Y.C.; Yang, S.F.; Tai, K.W.; Chou, M.Y.; Hsieh, Y.S. Increased tissue inhibitor of metalloproteinase-1 expression and inhibition of gelatinase a activity in buccal mucosal fibroblasts by arecoline as possible mechanisms for oral submucous fibrosis. Oral Oncol. 2002, 38, 195–200. [Google Scholar]
- Shieh, D.H.; Chiang, L.C.; Lee, C.H.; Yang, Y.H.; Shieh, T.Y. Effects of arecoline, safrole, and nicotine on collagen phagocytosis by human buccal mucosal fibroblasts as a possible mechanism for oral submucous fibrosis in taiwan. J. Oral Pathol. Med. 2004, 33, 581–587. [Google Scholar]
- Shieh, D.H.; Chiang, L.C.; Shieh, T.Y. Augmented mrna expression of tissue inhibitor of metalloproteinase-1 in buccal mucosal fibroblasts by arecoline and safrole as a possible pathogenesis for oral submucous fibrosis. Oral Oncol. 2003, 39, 728–735. [Google Scholar]
- Xia, L.; Tian-You, L.; Yi-Jun, G.; Dong-Sheng, T.; Wen-Hui, L. Arecoline and oral keratinocytes may affect the collagen metabolism of fibroblasts. J. Oral Pathol. Med. 2009, 38, 422–426. [Google Scholar]
- Chang, M.C.; Chan, C.P.; Wang, W.T.; Chang, B.E.; Lee, J.J.; Tseng, S.K.; Yeung, S.Y.; Hahn, L.J.; Jeng, J.H. Toxicity of areca nut ingredients: Activation of chk1/chk2, induction of cell cycle arrest, and regulation of mmp-9 and timps production in sas epithelial cells. Head Neck 2012, 35, 1295–1302. [Google Scholar]
- Tseng, Y.H.; Chang, K.W.; Liu, C.J.; Lin, C.Y.; Yang, S.C.; Lin, S.C. Areca nut extract represses migration and differentiation while activating matrix metalloproteinase-9 of normal gingival epithelial cells. J. Periodontal Res. 2008, 43, 490–499. [Google Scholar]
- Chang, M.C.; Wu, H.L.; Lee, J.J.; Lee, P.H.; Chang, H.H.; Hahn, L.J.; Lin, B.R.; Chen, Y.J.; Jeng, J.H. The induction of prostaglandin e2 production, interleukin-6 production, cell cycle arrest, and cytotoxicity in primary oral keratinocytes and kb cancer cells by areca nut ingredients is differentially regulated by mek/erk activation. J. Biol. Chem. 2004, 279, 50676–50683. [Google Scholar]
- Jeng, J.H.; Wang, Y.J.; Chiang, B.L.; Lee, P.H.; Chan, C.P.; Ho, Y.S.; Wang, T.M.; Lee, J.J.; Hahn, L.J.; Chang, M.C. Roles of keratinocyte inflammation in oral cancer: Regulating the prostaglandin e2, interleukin-6 and tnf-alpha production of oral epithelial cells by areca nut extract and arecoline. Carcinogenesis 2003, 24, 1301–1315. [Google Scholar]
- Dai, J.P.; Zhao, X.F.; Zeng, J.; Wan, Q.Y.; Yang, J.C.; Li, W.Z.; Chen, X.X.; Wang, G.F.; Li, K.S. Drug screening for autophagy inhibitors based on the dissociation of beclin1-bcl2 complex using bifc technique and mechanism of eugenol on anti-influenza a virus activity. PLoS ONE 2013, 8, e61026. [Google Scholar]
- Sample Availability: Samples of the compounds tanshinone IIA1, salvianolic acid A and salvianolic acid B are available from the authors.
© 2015 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license ( http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Dai, J.-P.; Zhu, D.-X.; Sheng, J.-T.; Chen, X.-X.; Li, W.-Z.; Wang, G.-F.; Li, K.-S.; Su, Y. Inhibition of Tanshinone IIA, Salvianolic Acid A and Salvianolic Acid B on Areca Nut Extract-Induced Oral Submucous Fibrosis in Vitro. Molecules 2015, 20, 6794-6807. https://doi.org/10.3390/molecules20046794
Dai J-P, Zhu D-X, Sheng J-T, Chen X-X, Li W-Z, Wang G-F, Li K-S, Su Y. Inhibition of Tanshinone IIA, Salvianolic Acid A and Salvianolic Acid B on Areca Nut Extract-Induced Oral Submucous Fibrosis in Vitro. Molecules. 2015; 20(4):6794-6807. https://doi.org/10.3390/molecules20046794
Chicago/Turabian StyleDai, Jian-Ping, Dan-Xia Zhu, Jiang-Tao Sheng, Xiao-Xuan Chen, Wei-Zhong Li, Ge-Fei Wang, Kang-Sheng Li, and Yun Su. 2015. "Inhibition of Tanshinone IIA, Salvianolic Acid A and Salvianolic Acid B on Areca Nut Extract-Induced Oral Submucous Fibrosis in Vitro" Molecules 20, no. 4: 6794-6807. https://doi.org/10.3390/molecules20046794