
Bacillus toyonensis Strain AEMREG6, a Bacterium Isolated from South African Marine Environment Sediment Samples Produces a Glycoprotein Bioflocculant


Abstract
:1. Introduction
2. Results and Discussion
2.1. Identification of the Bioflocculant-Producing Bacteria
2.2. Optimization of Culture Conditions for REG-6 Production
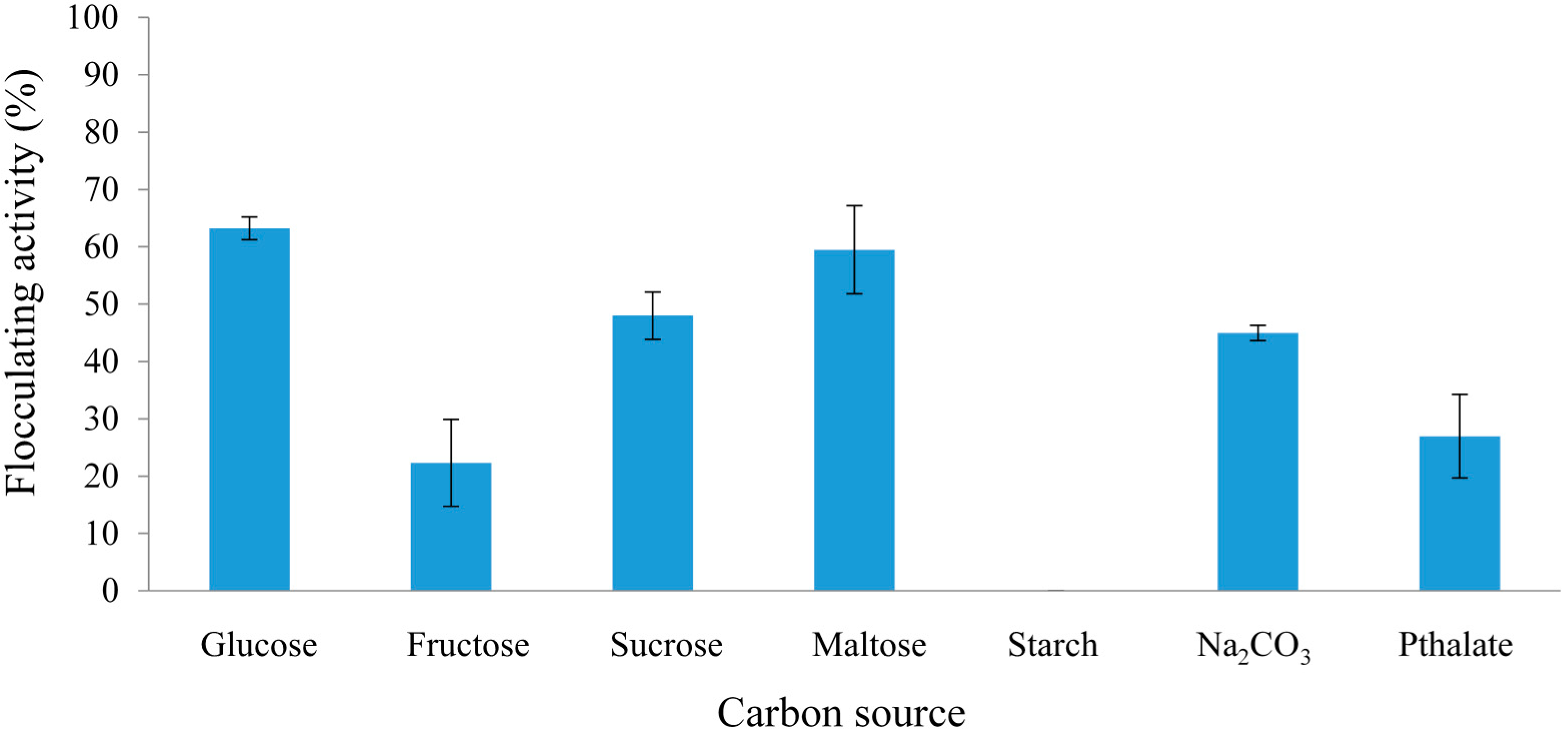
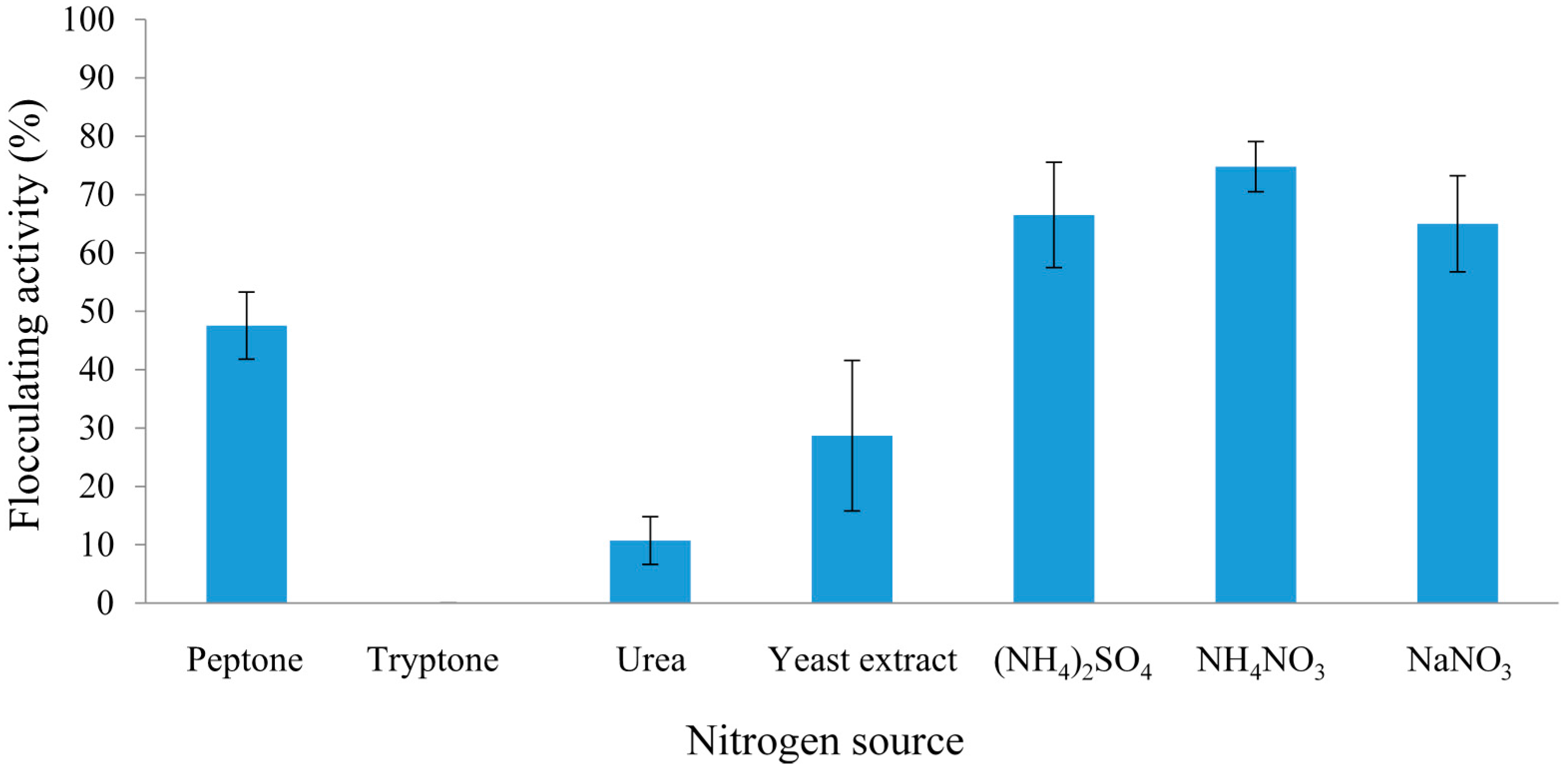
2.2.1. Effect of Carbon and Nitrogen Sources on REG-6 Production
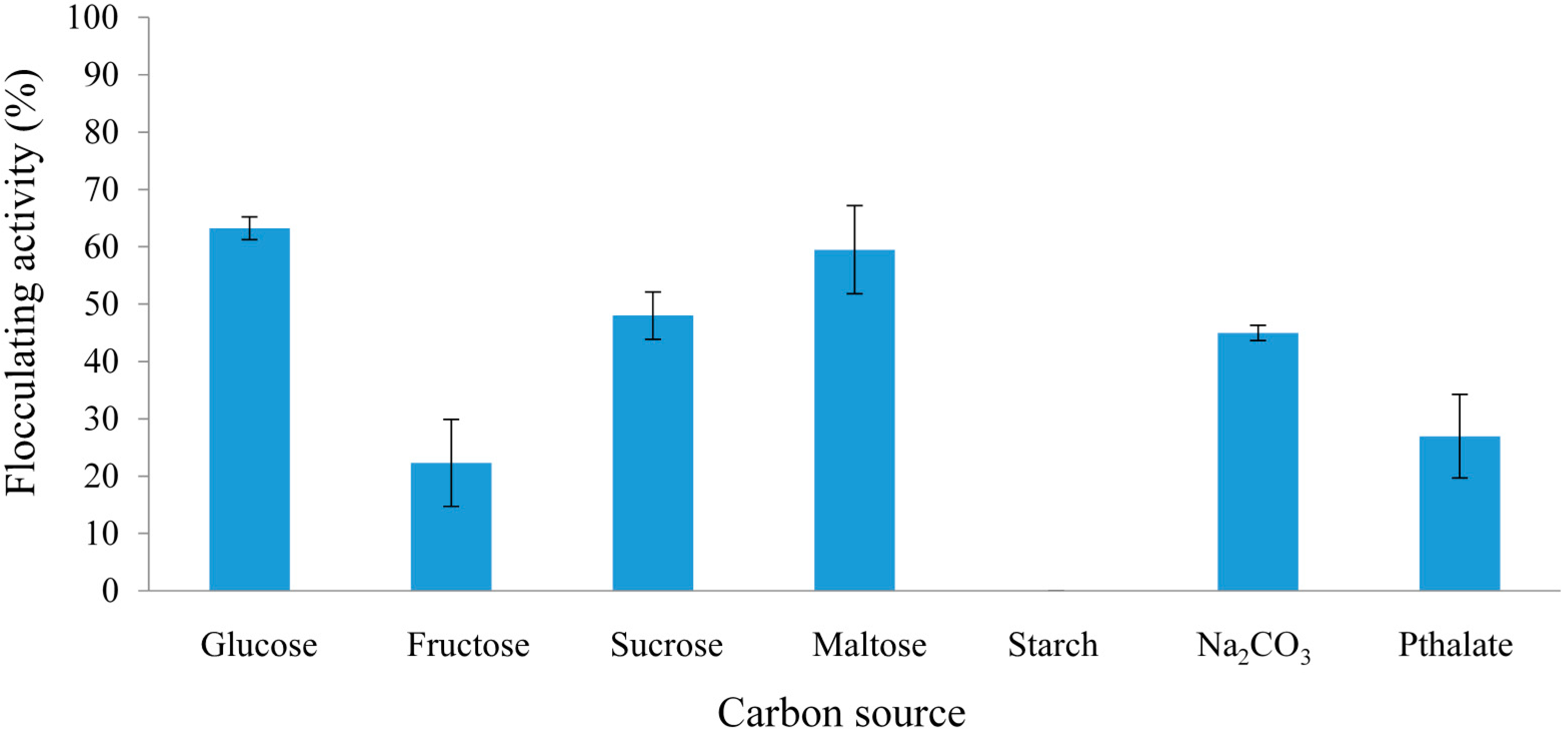
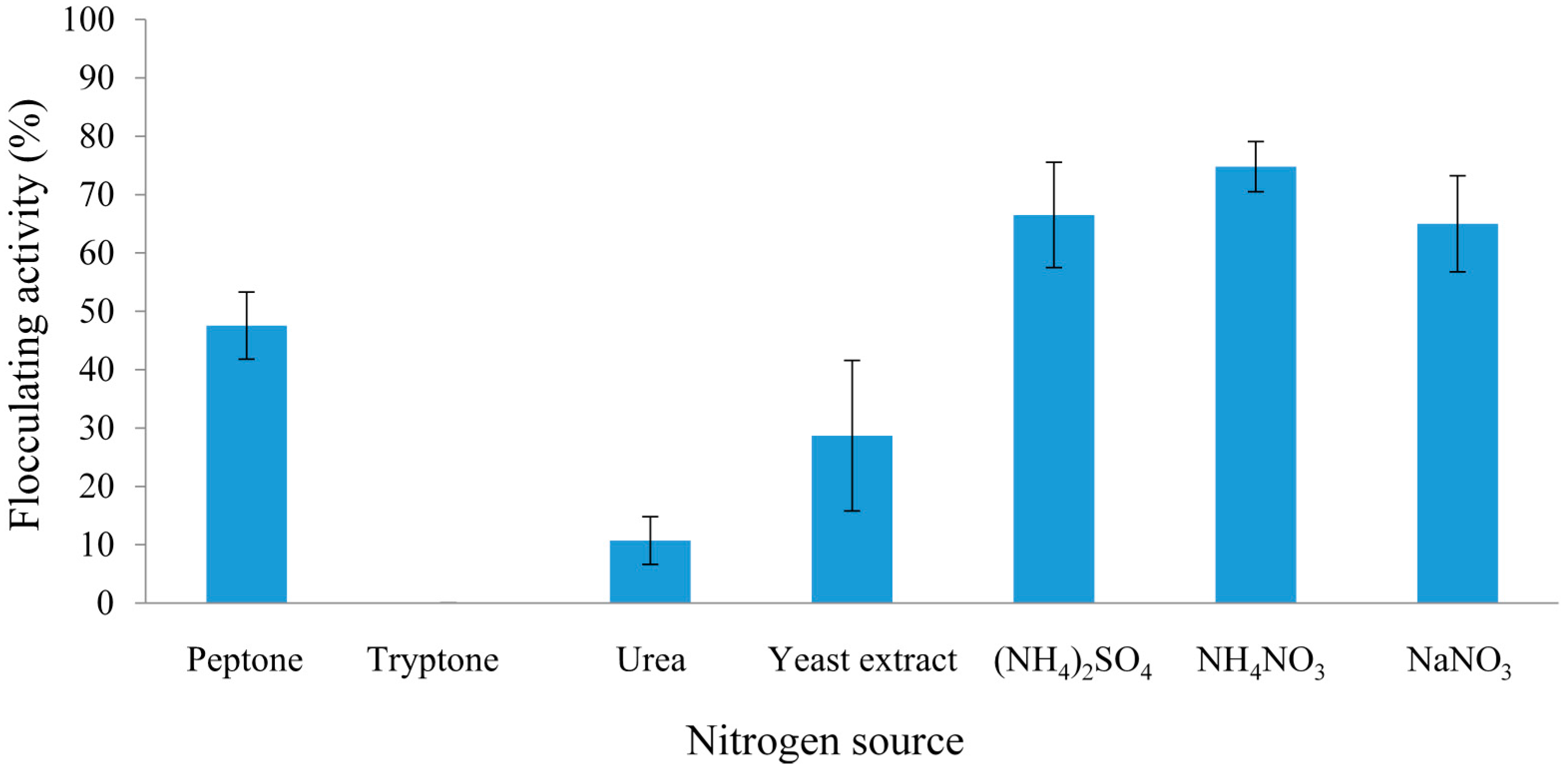
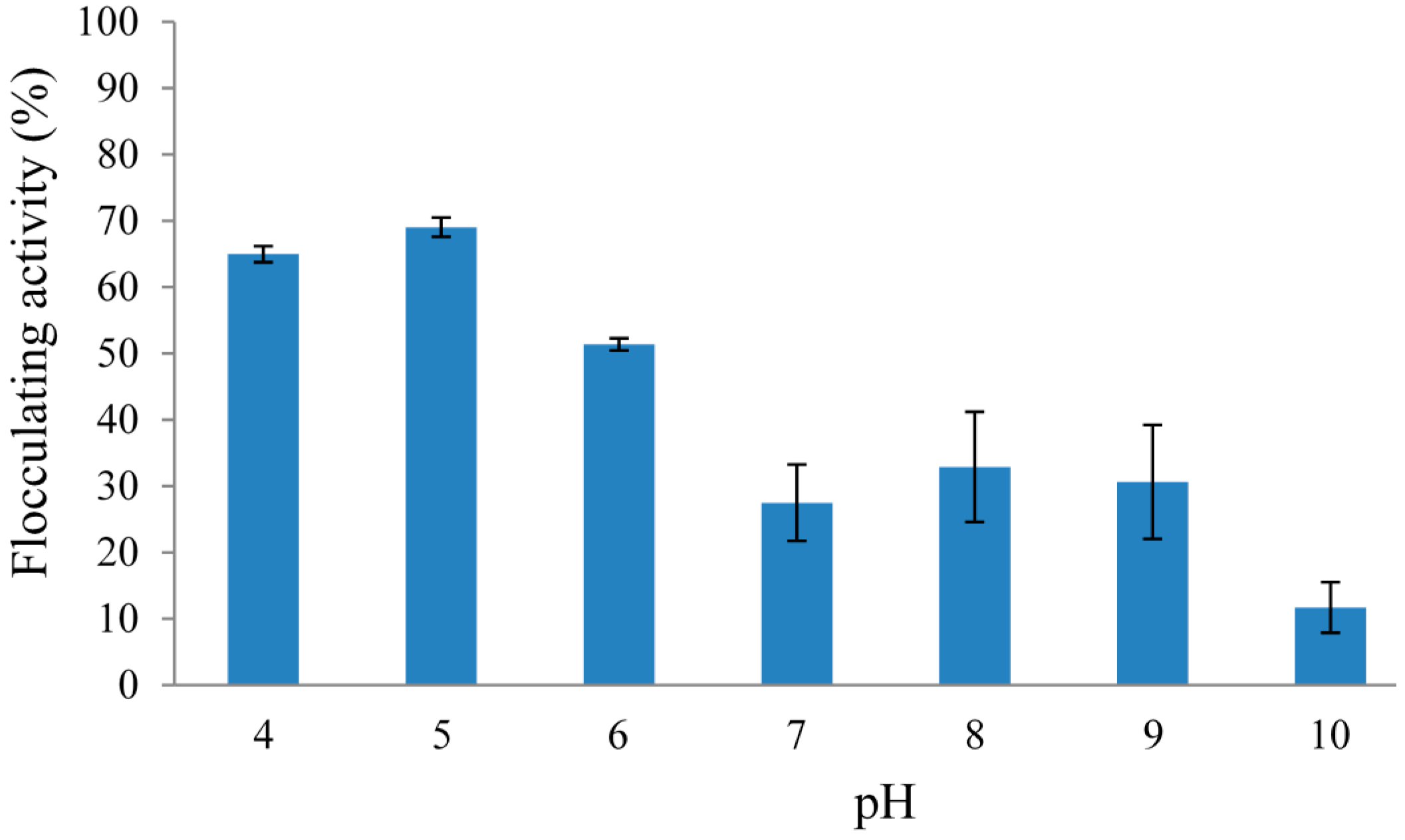
2.2.2. Effect of Initial pH of Growth Medium on REG-6 Production
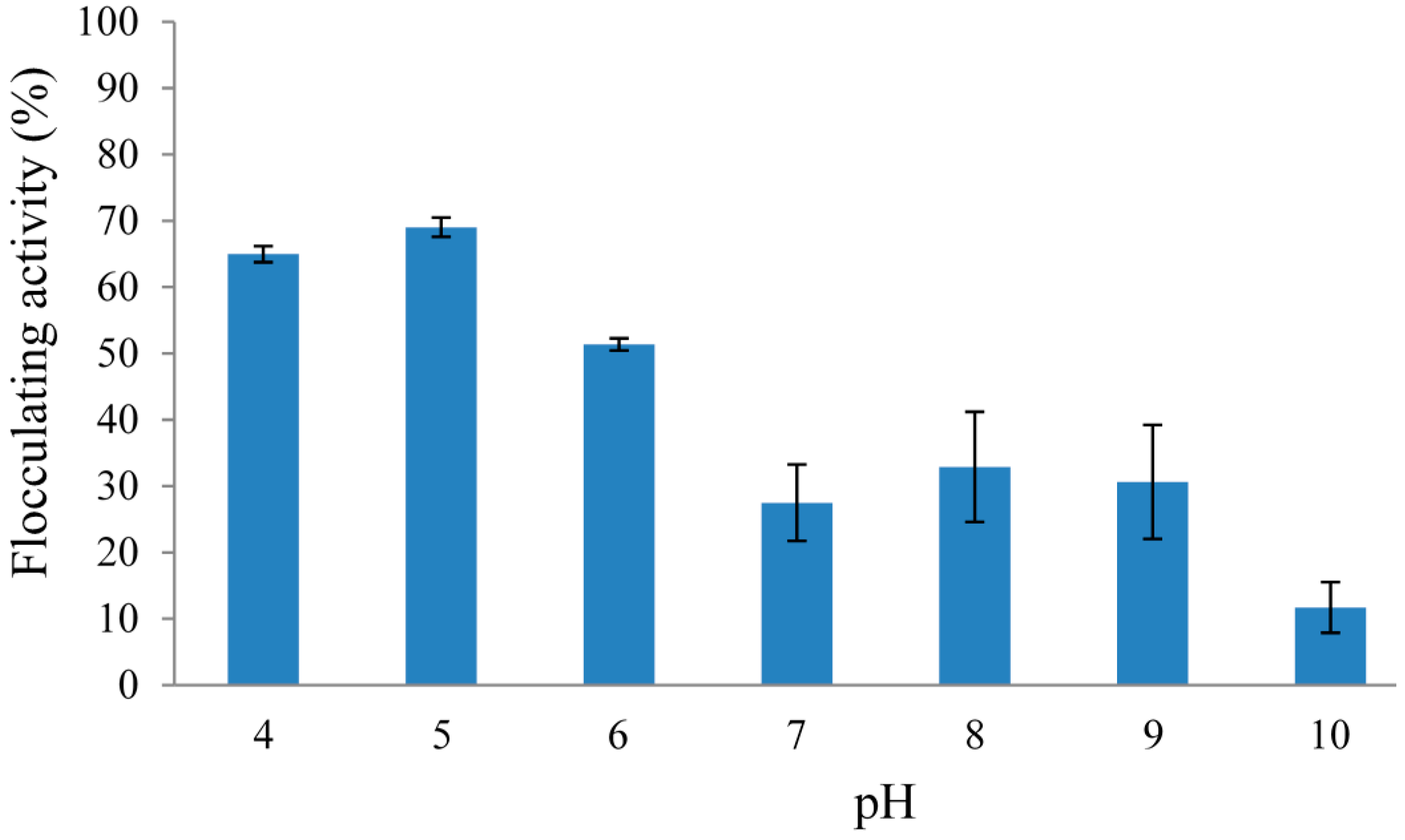
2.2.3. Effect of Inoculum Size on REG-6 Production

2.3. Optimization of the Flocculating Conditions of Crude REG-6
2.3.1. Effect of Cations on the Flocculating Activity of Crude REG-6

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Cations (w/v) | Flocculating Activity (%) |
|---|---|
| Li+ | - |
| K+ | - |
| Na+ | - |
| Mg2+ | 86.82 |
| Ca2+ | 84.96 |
| Mn2+ | 89.5 |
| Fe3+ | - |
| Al3+ | - |
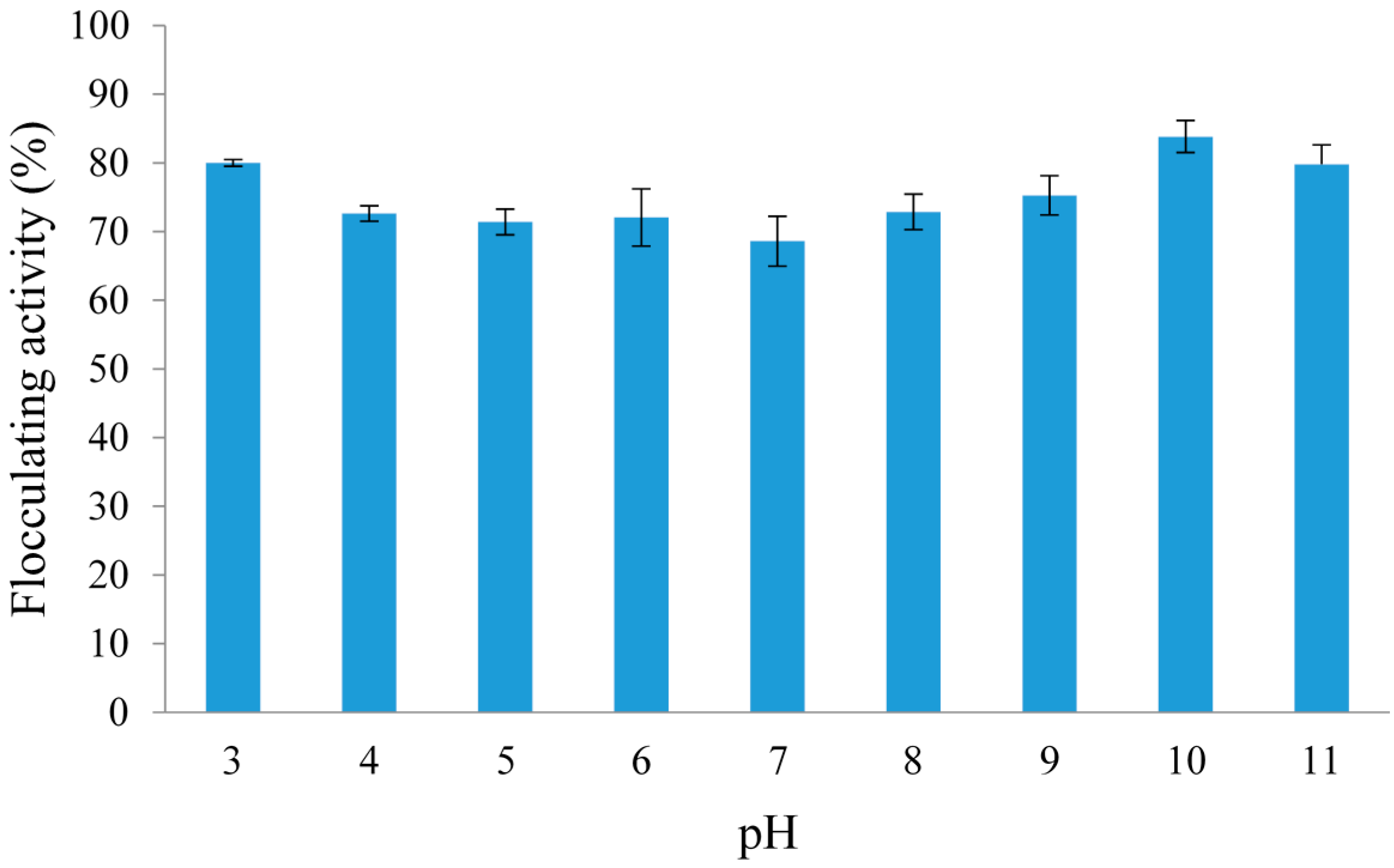
2.3.2. Effect of pH on the Flocculating Activity of Crude REG-6

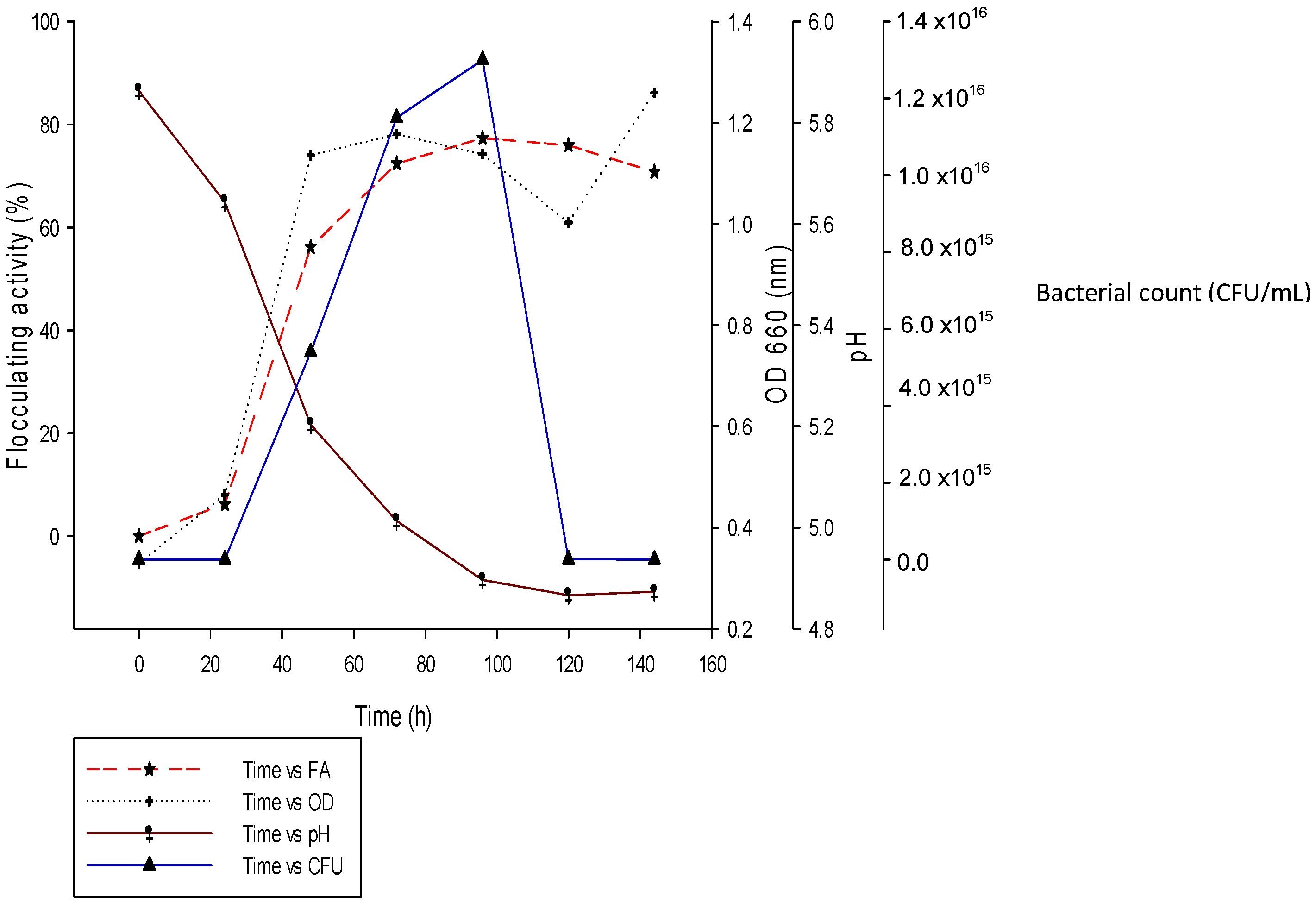
2.4. Time Course of REG-6 Production by Bacillus Toyonensis Strain AEMREG6

2.5. Characterization of the Purified REG-6
2.5.1. REG-6 Dosage
| Dosage (mg/mL) | FA (%) | pH | FA (%) | Cations | FA (%) | Temp (°C) | FA (%) |
|---|---|---|---|---|---|---|---|
| 0.05 | 48.23 | 3 | 80 | Li+ | - | 50 | 81.79 |
| 0.1 | 64.03 | 4 | 72.64 | K+ | - | 60 | 81.87 |
| 0.2 | 54.29 | 5 | 71.41 | Na+ | - | 70 | 82.74 |
| 0.3 | 55.37 | 6 | 72.05 | Mg2+ | 86.82 | 80 | 81.94 |
| 0.4 | 46.79 | 7 | 68.61 | Ca2+ | 84.96 | - | - |
| 0.5 | 45.51 | 8 | 72.86 | Mn2+ | 89.51 | - | - |
| - | - | 9 | 75.29 | Fe3+ | - | - | - |
| - | - | 10 | 83.84 | Al3+ | - | - | - |
| - | - | 11 | 79.80 | - | - | - | - |
2.5.2. Effect of pH on the Flocculating Activity of REG-6
2.5.3. Effect of Cations on the Flocculating Activity of REG-6
2.5.4. Thermal Stability of REG-6
2.5.5. REG-6 Yield and Chemical Analysis
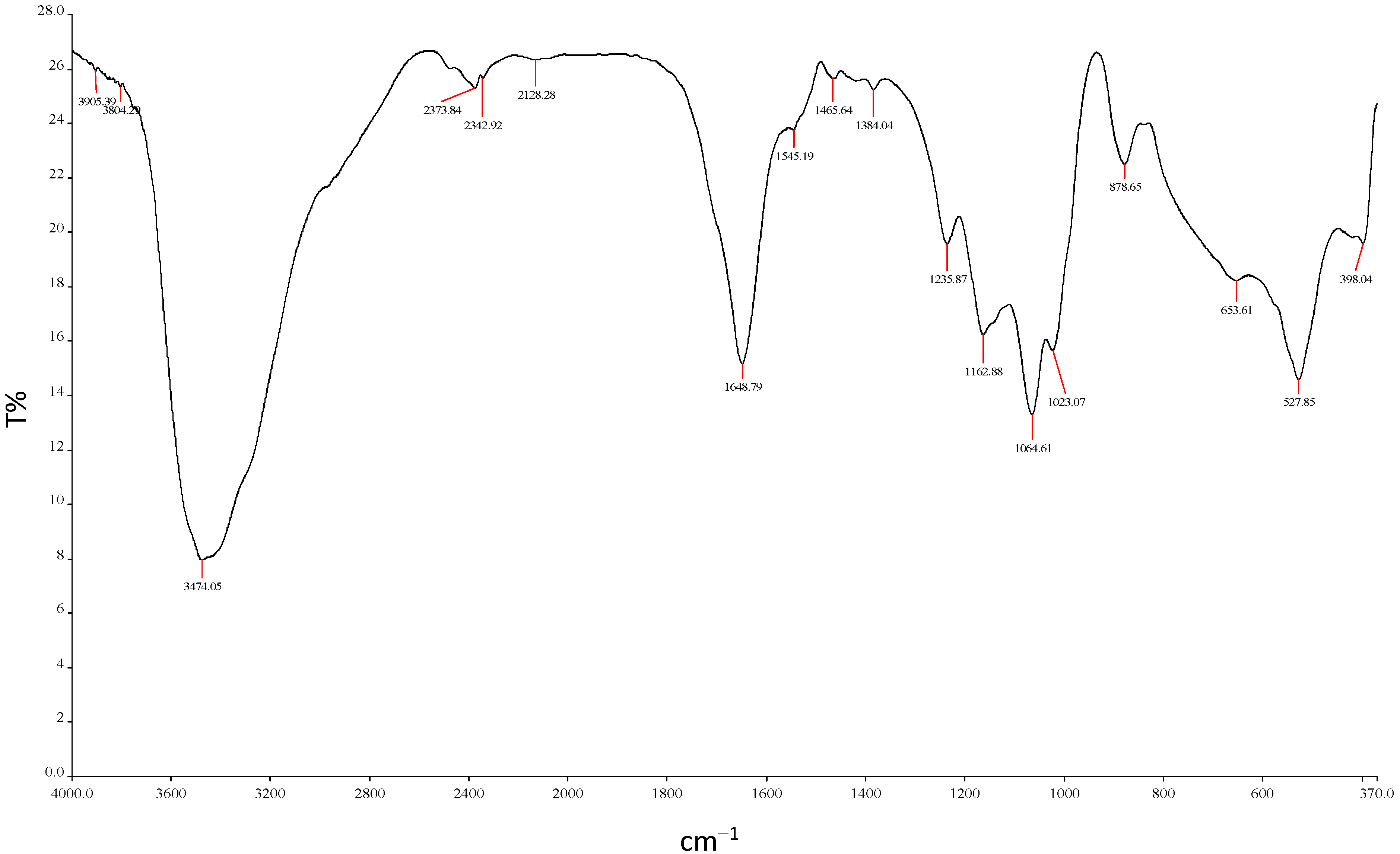
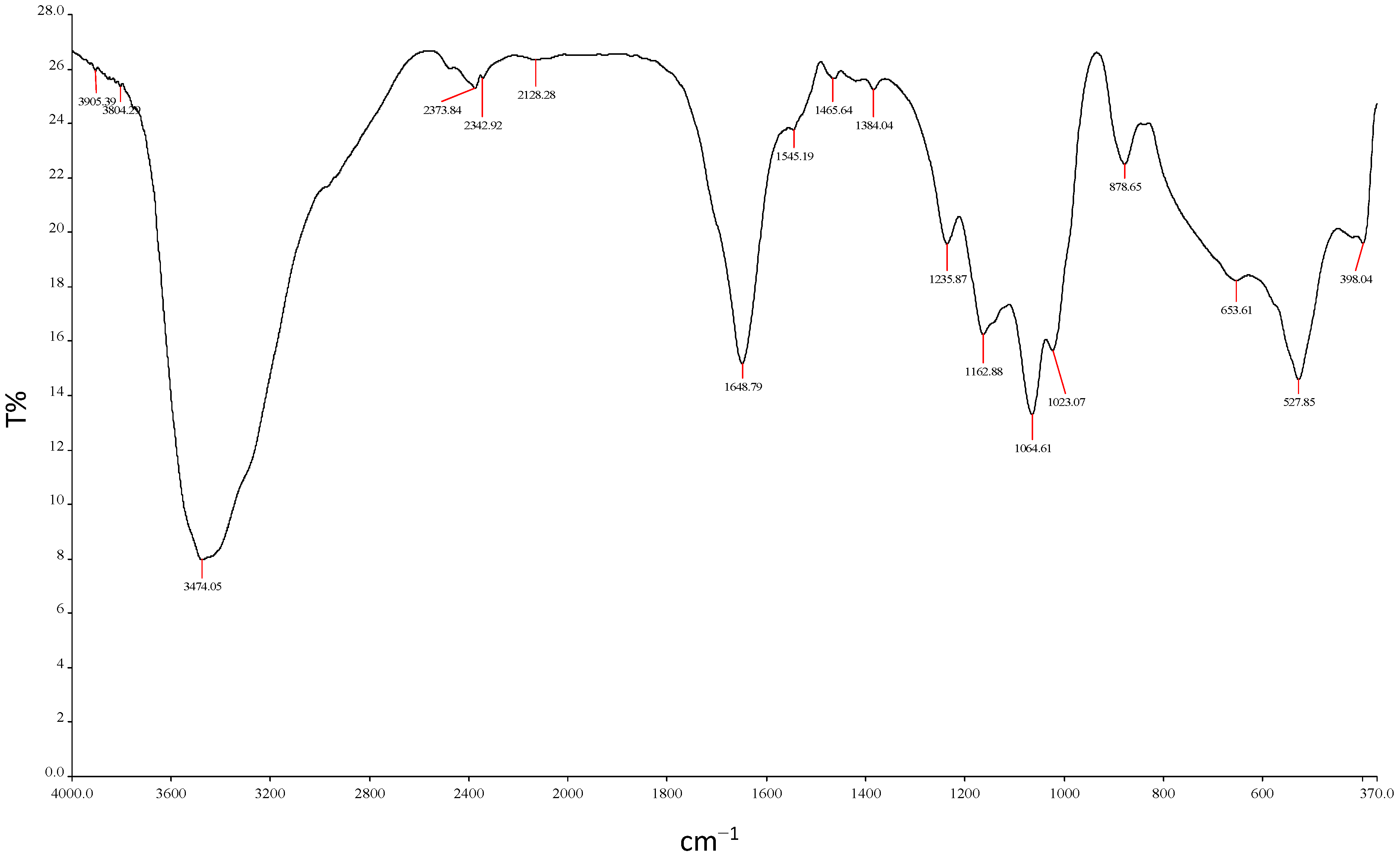
2.5.6. Functional Group Analysis by FTIR
2.5.7. Scanning Electron Microscopy (SEM) Images

3. Experimental Section
3.1. Sample Collection and Isolation of Bioflocculant-Producing Bacteria
3.2. Media and Cultivation Conditions
3.3. Screening of Bioflocculant-Producing Bacteria
3.4. Determination of Flocculating Activity of REG-6
3.5. Identification of the Bioflocculant-Producing Bacteria
3.6. Optimization of Culture Conditions for REG-6 Production
3.6.1. Effect of Carbon and Nitrogen Sources on REG-6 Production
3.6.2. Effect of Initial pH of Growth Medium on REG-6 Production
3.6.3. Effect of Inoculum Size on REG-6 Production
3.7. Optimization of Flocculating Conditions of Crude REG-6
3.7.1. Effect of Cations on Flocculating Activity of Crude REG-6
3.7.2. Effect of pH on the Flocculating Activity of Crude REG-6
3.8. Time Course of REG-6 Production by Bacillus Toyonensis Strain AEMREG6
3.9. Extraction and Purification of REG-6
3.10. Characterization of the Purified REG-6
3.10.1. Chemical Composition Analysis of REG-6
3.10.2. Effect of REG-6 Dosage on Flocculating Activity
3.10.3. Effect of pH on the Flocculating Activity of REG-6
3.10.4. Effect of Cations on the Flocculating Activity of REG-6
3.10.5. Thermal Stability of REG-6
3.10.6. FTIR Analysis of REG-6
3.10.7. Scanning Electron Microscopy (SEM) Images
3.11. Statistical Analysis
4. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Tang, J.; Qi, S.; Li, Z.; An, Q.; Xie, M.; Yang, B.; Wang, Y. Production, purification and application of polysaccharide-based bioflocculant by Paenibacillus mucilaginosus. Carbohydr. Polym. 2014, 113, 463–470. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Ma, F.; Lee, D.J.; Wang, A.; Ren, N. Bioflocculants from hydrolysates of corn stover using isolated strain Ochrobactium ciceri W2. Bioresour. Technol. 2013, 145, 145259–145263. [Google Scholar]
- Liu, Z.Y.; Hu, Z.Q.; Wang, T.; Chen, Y.Y.; Zhang, J.; Yu, J.R.; Zhang, T.; Zhang, Y.F.; Li, Y.L. Production of novel microbial flocculants by Klebsiella sp. TG-1 using waste residue from the food industry and its use in defecating the trona suspension. Bioresour. Technol. 2013, 139, 265–271. [Google Scholar] [CrossRef] [PubMed]
- Flemming, H.; Wingender, J. The biofilm matrix. Nat. Rev. Microbiol. 2010, 8, 623–633. [Google Scholar] [PubMed]
- Lin, J.; Harichund, C. Isolation and characterization of heavy metal removing bacterial bioflocculant. Afr. J. Biotechnol. 2011, 5, 599–607. [Google Scholar]
- Patil, S.V.; Patil, C.D.; Salunke, B.K.; Salunkhe, R.B.; Bathe, G.A.; Patil, D.M. Studies on characterization of bioflocculant exopolysaccharide of Azotobacter indicus and its potential for wastewater treatment. Appl. Biochem. Biotechnol. 2011, 163, 463–472. [Google Scholar] [CrossRef] [PubMed]
- Zhao, H.J.; Liu, H.T.; Zhou, J.G. Characterization of a bioflocculant MBF5 by Klebsiella pneumoniae and its application in Acanthamoeba cysts removal. Bioresour. Technol. 2013, 137, 226–232. [Google Scholar] [CrossRef] [PubMed]
- Gao, J.; Bao, H.Y.; Xin, M.X.; Liu, Y.X.; Li, Q.; Zhang, Y.F. Characterization of a bioflocculant from a newly isolated Vagococcus sp. W31. J. Zhejiang Univ. Sci. B 2006, 7, 186–192. [Google Scholar] [CrossRef] [PubMed]
- Piyo, N.; Cosa, S.; Mabinya, L.V.; Okoh, A.I. Assessment of bioflocculant production by Bacillus sp. Gilbert, a marine bacteria isolated from the bottom sediment of Algoa. Bay. Mar. Drugs 2011, 9, 1232–1242. [Google Scholar] [CrossRef] [PubMed]
- Okaiyeto, K.; Nwodo, U.U.; Mabinya, L.V.; Okoh, A.I. Characterization of a bioflocculant produced by a consortium of Halomonas sp. Okoh and Micrococcus sp. Leo. Int. J. Environ. Res. Public Health 2013, 10, 5097–5110. [Google Scholar] [CrossRef] [PubMed]
- He, N.; Li, Y.; Chen, J. Production of a novel polygalacturonic acid bioflocculant REA-11 by Corynebacteria glutamicum. Bioresour. Technol. 2004, 94, 99–105. [Google Scholar] [CrossRef] [PubMed]
- Vijayalakshmi, S.P.; Raichur, A.M. The utility of Bacillus subtilis as a bioflocculant for fine coal. Colloids Surf. B. Biointerfaces 2003, 29, 265–275. [Google Scholar] [CrossRef]
- Adebayo-Tayo, B.; Adebami, G.E. Production and characterization of bioflocculant produced by Bacillus clausii NB2. Innov. Rom. Food Biotechnol. 2014, 14, 13–25. [Google Scholar]
- Elkady, M.F.; Farag, S.; Sahar, Z.; Gadallah, A.; Desouky, A. Bacillus mojavensis strain 32A, a bioflocculant-producing bacteria isolated from an Egyptian salt production pond. Bioresour. Technol. 2011, 102, 8143–8151. [Google Scholar] [CrossRef] [PubMed]
- Zaki, S.A.; Marwa, F.; Elkady, S.F.; Desouky, A. Characterization and flocculation properties of a carbohydrate bioflocculant from a newly isolated Bacillus velezensis 40B. J. Environ. Biol. 2013, 34, 51–58. [Google Scholar] [PubMed]
- Ugbenyen, A.M.; Cosa, S.; Mabinya, L.V.; Okoh, A.I. Bioflocculant production by Bacillus sp. Gilbert isolated from a marine environment in South Africa. Appl. Biochem. Microbiol. 2014, 50, 49–54. [Google Scholar] [CrossRef]
- Lauro, F.M.; McDougald, D.; Thomas, T.; Williams, T.J.; Egan, S.; Rice, S.; DeMaere, M.Z.; Ting, L.; Ertan, H.; Johnson, J.; et al. The genomic basis of trophic strategy in marine bacteria. Proc. Natl. Acad. Sci. USA 2009, 106, 15527–15533. [Google Scholar] [CrossRef] [PubMed]
- Dash, H.R.; Neelam, M.; Jaya, C.; Supriya, K.; Surajit, D. Marine bacteria: Potential candidates for enhanced bioremediation. Appl. Microbiol. Biotechnol. 2013, 97, 561–571. [Google Scholar] [CrossRef] [PubMed]
- Jiméneza, M.U.; Ana, C.; Aránzazu, L.; Anicet, R.B.; Javier, T.; Peter, K.; Anne-Brit, K.; Daniel, R.; Juan, F.; Martínezg, F.; et al. Description of Bacillus toyonensis sp. nov., a novel species of the Bacillus cereus group, and pairwise genome comparisons of the species of the group by means of ANI calculations Guillermo. Syst. Appl. Microbiol. 2013, 36, 383–391. [Google Scholar] [CrossRef] [PubMed]
- Abdel-Aziz, S.M.; Hoda, A.H.; Foukia, E.M.; Nayera, A.M. Extracellular metabolites produced by a novel strain, Bacillus alvei NRC-14: 3. Synthesis of a bioflocculant that has chitosan-like. Struct. Life Sci. J. 2011, 8, 883–890. [Google Scholar]
- Zheng, Y.; Ye, Z.L.; Fang, X.L.; Li, Y.H.; Cai, W.M. Production and characteristics of a bioflocculant produced by Bacillus sp. F19. Bioresour. Technol. 2008, 99, 7686–7691. [Google Scholar] [CrossRef] [PubMed]
- Li, Z.; Zhong, S.; Lei, H.; Chen, R.; Yu, Q.; Li, H. Production of a novel bioflocculant by Bacillus licheniformis X14 and its application to low temperature drinking water treatment. Bioresour. Technol. 2009, 100, 3650–3656. [Google Scholar] [CrossRef] [PubMed]
- Sathiyanarayanana, G.; Seghal, K.G.; Joseph, S. Synthesis of silver nanoparticles by polysaccharide bioflocculant produced from marine Bacillus subtilis MSBN17. Colloids Surf. B Biointerf. 2013, 102, 13–20. [Google Scholar] [CrossRef]
- Bereka, M.M. Improved exopolysaccharide production by Bacillus licheniformis strain-QS5 and application of statistical experimental design. Int. J. Curr. Microbiol. Appl. Sci. 2014, 3, 876–886. [Google Scholar]
- Cosa, S.; Ugbenyen, M.A.; Mabinya, L.V.; Okoh, I.A. Characterization of a thermostable polysaccharide bioflocculant produced by Virgibacillus species isolated from Algoa bay. Afr. J. Microbiol. Res. 2013, 7, 2925–2938. [Google Scholar]
- Nwodo, U.U.; Agunbiade, M.O.; Green, E.; Nwamadi, M.; Rumbold, K.; Okoh, A.I. Characterization of an exopolymeric flocculant produced by a Brachybacteria sp. Materials 2013, 6, 1237–1254. [Google Scholar] [CrossRef]
- Cosa, S.; Ugbenyen, M.A.; Mabinya, L.V.; Rumbold, K.; Olaniran, A.O.; Aghdasi, F.; Okoh, A.I. A marine bacteria, Oceanobacillus sp. Pinky, isolated from Algoa Bay sediment produces a thermostable glycoprotein flocculant. Afr. J. Biotechnol. 2013, 12, 4135–4146. [Google Scholar]
- Wan, C.; Zhao, X.Q.; Guo, S.L.; Alam, M.A.; Bai, F.W. Bioflocculant production from Solibacillus silvestris W01 and its application in cost-effective harvest of marine microalga Nannochloropsis oceanica by flocculation. Bioresour. Technol. 2013, 135, 207–212. [Google Scholar] [CrossRef] [PubMed]
- Deng, S.B.; Yu, G.; Ting, Y.P. Production of a bioflocculant by Aspergillus parasiticus and its application in dye removal. Colloids Surf. B Biointerfaces 2005, 44, 179–186. [Google Scholar] [CrossRef] [PubMed]
- He, J.; Zhen, Q.; Qiu, N.; Liu, Z.; Wang, B.; Shao, Z.; Yu, Z. Medium optimization for the production of a novel bioflocculant from Halomonas sp. V3a using response surface methodology. Bioresour. Technol. 2009, 100, 5922–5927. [Google Scholar] [CrossRef] [PubMed]
- Yan, D.; Yun, J. Screening of bioflocculant-producing strains and optimization of its nutritional conditions by using potato starch wastewater. Trans. Chin. Soc. Agric. Eng. 2013, 29, 198–206. [Google Scholar]
- Yim, J.H.; Kim, S.J.; Se, H.A.; Hong, K.L. Characterization of a novel bioflocculant, p-KG03, from a marine dinoflagellate, Gyrodinium impudicum KG03. Bioresour. Technol. 2007, 98, 361–367. [Google Scholar] [CrossRef] [PubMed]
- Aljuboori, A.H.R.; Yoshimitsu, U.; Noridah, B.O.; Suzana, Y. Production of a bioflocculant from Aspergillus niger using palm oil mill effluent as carbon source. Bioresour. Technol. 2014, 171, 66–70. [Google Scholar] [CrossRef] [PubMed]
- Abu-Elreesh, G.; Zaki, S.; Farag, S.; Elkady, M.F.; Abd-El-Haleem, D. Exobiopolymer from polyhydroxyalkanoate-producing transgenic yeast. Afr. J. Biotechnol. 2011, 10, 6558–6563. [Google Scholar]
- Liu, L.; Cheng, W. Characteristics and culture conditions of a bioflocculant produced by Penicillium sp. Biomed. Environ. Sci. 2010, 23, 213–218. [Google Scholar] [CrossRef] [PubMed]
- Ugbenyen, A.; Cosa, S.; Mabinya, L.V.; Babalola, O.O.; Aghdasi, F.; Okoh, A.I. Thermostable bacterial bioflocculant produced by Cobetia spp. isolated from Algoa Bay (South Africa). Int. J. Environ. Res. Public Health 2012, 9, 2108–2120. [Google Scholar] [CrossRef] [PubMed]
- Xia, S.; Zhang, Z.; Wang, X.; Yang, A.; Chen, L.; Zhao, J.; Leonard, D.; Jaffrezic-Renault, N. Production and characterization of bioflocculant by Proteus mirabilis TJ-1. Bioresour. Technol. 2008, 99, 6520–6527. [Google Scholar] [CrossRef] [PubMed]
- Liu, W.J.; Kai, W.; Li, B.Z.; Hong, L.Y.; Jin, S.Y. Production and characterization of an intracellular bioflocculant by Chryseobacteria daeguense W6 cultured in low nutrition medium. Bioresour. Technol. 2010, 101, 1044–1048. [Google Scholar] [CrossRef] [PubMed]
- Zulkeflee, Z.; Aris, A.Z.; Shamsuddin, Z.H.; Yusoff, M.K. Cation dependence, pH tolerance, and dosage requirement of a bioflocculant produced by Bacillus spp. UPMB13: Flocculation performance optimization through kaolin assays. Sci. World J. 2012, 7. [Google Scholar] [CrossRef]
- Salehizadeh, H.; Shojaosadati, S.A. Extracellular biopolymeric flocculants: Recent trends and biotechnological importance. Biotechnol. Adv. 2001, 19, 371–385. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.; Gong, W.; Liu, X.; Lin, T.; Yue, Y.; Gao, B. Production of a novel bioflocculant by culture of Klebsiella mobilis using dairy wastewater. Biochem. Eng. J. 2007, 36, 81–86. [Google Scholar] [CrossRef]
- Wu, J.; Ye, H.F. Characterization and flocculating properties of an extracellular biopolymer produced from a Bacillus subtillis DYU1 isolate. Process Biochem. 2007, 42, 1114–1123. [Google Scholar] [CrossRef]
- Fujita, M.; Ike, M.; Tachibana, S.; Kitada, G.; Kim, S.M.; Inoue, Z. Characterization of bioflocculant produced by Citrobacter sp. TKF04 from acetic and propionic acids. J. Biosci. Bioeng. 2000, 98, 40–46. [Google Scholar] [CrossRef]
- Yong, P.; Bo, S.; Yu, Z. Research on flocculation property of bioflocculant PGa21 Ca. Mod. Appl. Sci. 2009, 3, 106–112. [Google Scholar]
- Wang, L.; Ma, F.; Qu, Y.; Sun, D.; Li, A.; Guo, J.; Yu, B. Characterization of a compound bioflocculant produced by mixed culture of Rhizobium radiobacter F2 and Bacillus sphaeicus F6. World J. Microbiol. Biotechnol. 2011, 27, 2559–2565. [Google Scholar] [CrossRef]
- Gao, Q.; Zhu, X.H.; Mu, J.; Zhang, Y.; Dong, X.W. Using Ruditapes philippinarum conglutination mud to produce bioflocculant and its applications in wastewater treatment Bioresour. Technol. 2009, 100, 4996–5001. [Google Scholar]
- Ji, B.; Zhang, X.Y.; Li, Z.; Xie, H.Q.; Xiao, X.M.; Fan, G.J. Flocculation properties of a bioflocculant produced by Bacillus licheniformis. Water Sci. Technol. 2010, 62, 1907–1913. [Google Scholar] [CrossRef] [PubMed]
- Salehizadeh, H.; Yan, N. Recent advances in extracellular biopolymer flocculants. Biotechnol. Adv. 2014, 32, 1506–1522. [Google Scholar] [CrossRef] [PubMed]
- Lu, W.Y.; Zhang, T.; Zhang, D.Y.; Li, C.H.; Wen, J.P.; Du, L.X. A novel bioflocculant produced by Enterobacter aerogenes and its use in defaecating the trona suspension. Biochem. Eng. J. 2005, 27, 1–7. [Google Scholar] [CrossRef]
- Zhang, Z.Q.; Xia, S.Q.; Zhao, S.Q.; Zhang, J.F. Characterization and flocculation mechanism of high efficiency microbial flocculant TJ-F1 from Proteus mirabilis. Colloids Surf. B Biointerfaces 2010, 75, 247–251. [Google Scholar] [CrossRef] [PubMed]
- Luo, Z.; Li, C.; Changhong, C.; Wei, Z.; Ming, L.; Ye, H.; Jiangang, Z. Production and characteristics of a bioflocculant by Klebsiella pneumonia YZ-6 isolated from human saliva. Appl. Biochem. Biotechnol. 2014, 172, 1282–1292. [Google Scholar] [CrossRef] [PubMed]
- Pan, Y.Z.; Shi, B.; Zhang, Y. Research on flocculation property of bioflocculant PG.a21 Ca. Mod. Appl. Sci. 2009, 3, 106–112. [Google Scholar] [CrossRef]
- Zhu, C.; Chen, C.; Zhao, L.; Zhang, Y.; Yang, J.; Song, L.; Yang, S. Bioflocculant produced by Chlamydomonas reinhardtii. J. Appl. Phycol. 2012, 24, 1245–1251. [Google Scholar] [CrossRef]
- Li, X.M.; Yang, Q.; Huang, K.; Zeng, G.M.; Xiao, D.X.; Liu, J.J.; Long, W.F. Screening and characterization of a bioflocculant produced by Aeromonas sp. Biomed. Environ. Sci. 2007, 20, 274–278. [Google Scholar] [PubMed]
- Gong, W.; Wang, S.; Sun, X.; Liu, X.; Yue, Q.; Gao, B. Bioflocculant production by culture of Serratia ficaria and its application in wastewater treatment. Bioresour. Technol. 2008, 99, 4668–4674. [Google Scholar] [CrossRef] [PubMed]
- Li, W.W.; Zhou, W.Z.; Zhang, W.Z.; Wang, J.; Zhu, X.B. Flocculation behaviour and mechanism of an exopolysaccharide from the deep-sea psychrophilic bacteria Pseudoalteromonas sp. SM9913. Bioresour. Technol. 2008, 99, 6893–6899. [Google Scholar] [CrossRef] [PubMed]
- Cosa, S.; Mabinya, L.V.; Olaniran, A.O.; Okoh, O.O.; Okoh, A.I. Bioflocculant production by Virgibacillus sp. Rob isolated from the bottom sediment of Algoa Bay in the Eastern Cape, South Africa. Molecules 2011, 16, 2431–2442. [Google Scholar] [CrossRef] [PubMed]
- Liu, C.; Wang, K.; Jiang, J.-H.; Liu, W.-J.; Wang, J.-Y. A novel bioflocculant produced by a salt-tolerant, alkaliphilic and biofilm-forming strain Bacillus agaradhaerens C9 and its application in harvesting Chlorella minutissima UTEX2341. Biochem. Eng. J. 2015, 93, 166–172. [Google Scholar] [CrossRef]
- Gomaa, E.Z. Production and characteristics of a heavy metals removing bioflocculant produced by Pseudomonas aeruginosa. Pol. J. Microbiol. 2012, 61, 281–289. [Google Scholar] [PubMed]
- Zhang, D.; Hou, Z.; Liu, Z.; Wang, T. Experimental research on Phanerochaete chrysosporium as coal microbial flocculant. Int. J. Min. Sci. Technol. 2013, 23, 521–524. [Google Scholar] [CrossRef]
- Kavita, K.; Avinash, M.; Bhavanath, J. Extracellular polymeric substances from two biofilm forming Vibrio species: Characterization and applications. Carbohydr. Polym. 2013, 94, 882–888. [Google Scholar] [CrossRef] [PubMed]
- Nwodo, U.U.; Okoh, A.I. Characterization and flocculation properties of biopolymeric flocculant (glycosaminoglycan) produced by Cellulomonas sp. Okoh. J. Appl. Microbiol. 2012, 114, 1325–1337. [Google Scholar] [CrossRef]
- Mabrouk, M.E.M. Production of bioflocculant by the marine actinomycete Nocardiopsis aegyptia sp. nov. Life Sci. J. 2014, 11, 27–35. [Google Scholar]
- Jensen, P.R.; Erin, G.; Chrisy, M.; Tracy, J.; Mincer, W.F. Culturable marine actinomycete diversity from tropical Pacific Ocean sediments. Environ. Microbiol. 2005, 7, 1039–1048. [Google Scholar]
- Ntsaluba, L.; Nwodo, U.U.; Mabinya, L.V.; Okoh, A.I. Studies on bioflocculant production by a mixed culture of Methylobacteria sp. Obi and Actinobacterium sp. Mayor. BMC Biotechnol. 2013, 13. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Z.Q.; Lin, B.; Xia, S.Q.; Wang, X.J.; Yang, A.M. Production and application of a novel bioflocculant by multiple-microorganism consortia using brewery wastewater as carbon source. Environ. Sci. 2007, 19, 667–673. [Google Scholar] [CrossRef]
- Kurane, R.; Hatamochi, K.; Kakuno, T.; Kiyohara, M.; Hirano, M.; Taniguchi, Y. Production of a bioflocculant by Rhodococcus erythropolis S-l grown on alcohols. Biosci. Biotech. Biochem. 1994, 2, 428–429. [Google Scholar] [CrossRef]
- Abdel-Aziz, S.M.; Hamed, H.A.; Mouafi, F.E. Acidic exopolysaccharide flocculant produced by the fungus Mucor rouxii using Beet-Molasses. Res. Biotechnol. 2012, 3, 1–13. [Google Scholar]
- He, P.; Geng, L.; Wang, Z.; Mao, D.; Wang, J.; Xu, C. Fermentation optimization, characterization and bioactivity of exopolysaccharides from Funalia trogii. Carbohydr. Polym. 2012, 89, 17–23. [Google Scholar] [CrossRef] [PubMed]
- Yang, Q.; Luo, K.; Liao, D.; Li, X.; Wang, D.; Liu, X.; Zeng, G.; Li, X. A novel bioflocculant produced by Klebsiella sp. and its application to sludge dewatering. Water Environ. J. 2012, 26, 560–566. [Google Scholar] [CrossRef]
- Wong, Y.-S.; Ong, S.-A.; Teng, T.-T.; Aminah, L.N.; Kumaran, K. Production of bioflocculant by Staphylococcus cohnii ssp. from palm oil mill effluent (POME). Water Air Soil Pollut. 2012, 223, 3775–3781. [Google Scholar] [CrossRef]
- Chaplin, M.F.; Kennedy, J.F. Carbohydrate Analysis, 2nd ed.; Oxford University Press: New York, NY, USA, 1994. [Google Scholar]
- Bradford, M.M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef] [PubMed]
- Wan, Y.X.; Wu, L.P.; Tang, P.J.; Zhang, J.Y. Composition of extracellular polymeric substances produced by Geotrichum sp. J-5. Asian J. Chem. 2014, 26, 592–596. [Google Scholar]
- Sample Availability: Samples of the compounds are available from the authors.
© 2015 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license ( http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Okaiyeto, K.; Nwodo, U.U.; Mabinya, L.V.; Okoh, A.I. Bacillus toyonensis Strain AEMREG6, a Bacterium Isolated from South African Marine Environment Sediment Samples Produces a Glycoprotein Bioflocculant. Molecules 2015, 20, 5239-5259. https://doi.org/10.3390/molecules20035239
Okaiyeto K, Nwodo UU, Mabinya LV, Okoh AI. Bacillus toyonensis Strain AEMREG6, a Bacterium Isolated from South African Marine Environment Sediment Samples Produces a Glycoprotein Bioflocculant. Molecules. 2015; 20(3):5239-5259. https://doi.org/10.3390/molecules20035239
Chicago/Turabian StyleOkaiyeto, Kunle, Uchechukwu U. Nwodo, Leonard V. Mabinya, and Anthony I. Okoh. 2015. "Bacillus toyonensis Strain AEMREG6, a Bacterium Isolated from South African Marine Environment Sediment Samples Produces a Glycoprotein Bioflocculant" Molecules 20, no. 3: 5239-5259. https://doi.org/10.3390/molecules20035239