
The Content of Phenolic Compounds in Leaf Tissues of Aesculus glabra and Aesculus parviflora Walt.

Abstract
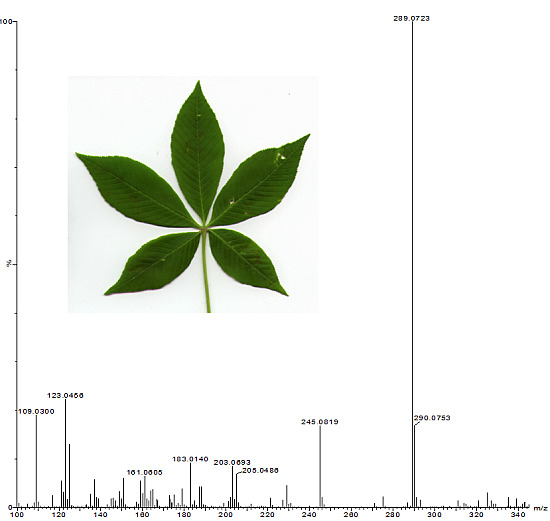
:
1. Introduction
2. Results and Discussion
2.1. Identification of Phenolics in Aesculus Leaves

{kind=link}
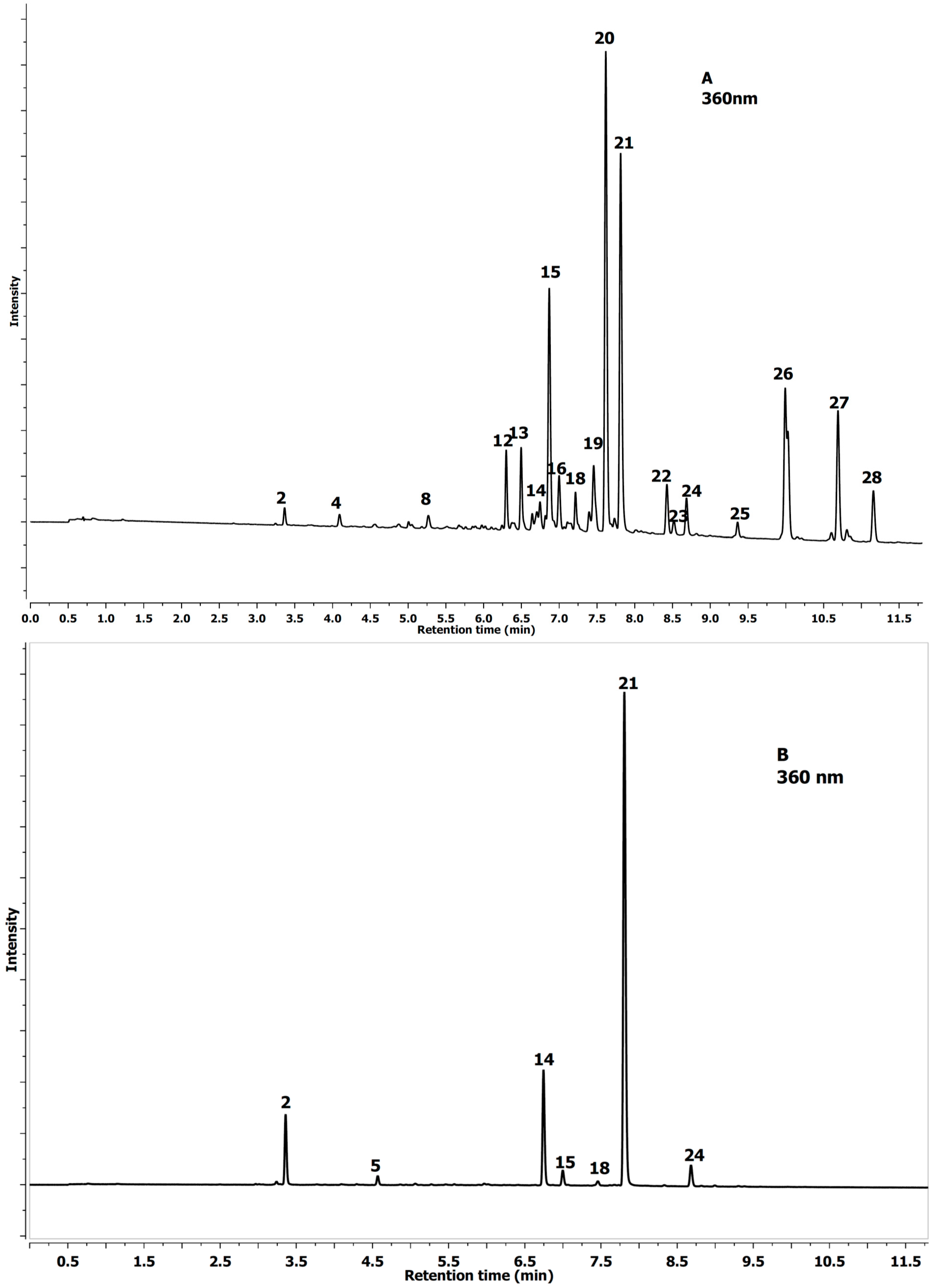{kind=link}
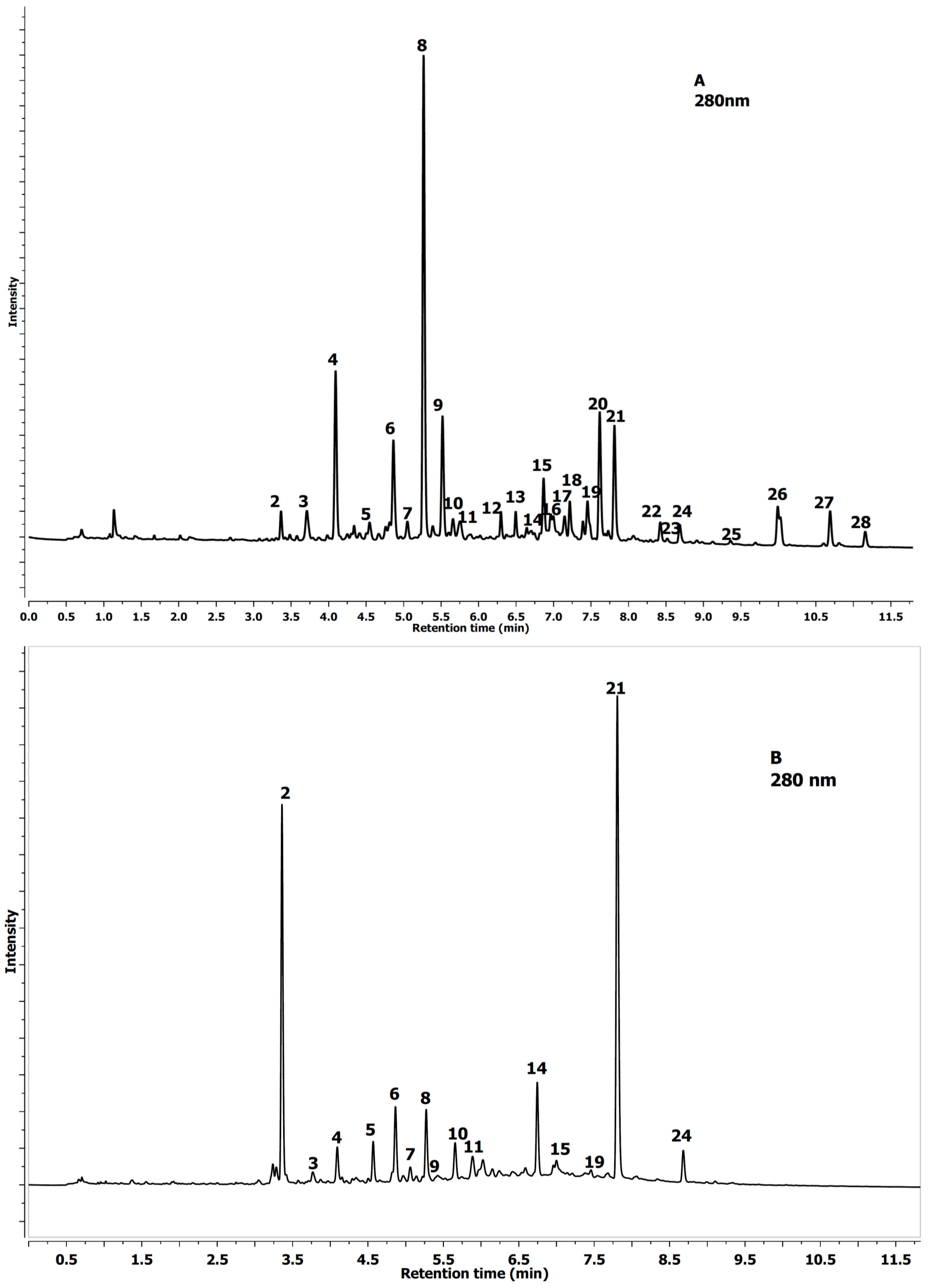{kind=link}
| No Peak | Tentative Identification | Rt (min) | λmax (nm) | [MS]− m/z | [MS−MS]− m/z |
|---|---|---|---|---|---|
| 1 | Protocatechuic-acid-4-glucoside | 2.69 | 292 | 315.0719 | 153.0180 |
| 2 | Neochlorogenic acid a | 3.37 | 324 | 353.0879 | 191.0552 |
| 3 | Procyanidin tetramer A-type | 3.71 | 280 | 1151.2501 | 289.0718 |
| 4 | 3-O-p-Coumaroylquinic acid | 4.09 | 309 | 337.0928 | 163.0394 |
| 5 | Chlorogenic acid a | 4.55 | 324 | 353.0888 | 191.0554 |
| 6 | Procyanidin B1 a | 4.86 | 280 | 577.1337 | 289.0708 |
| 7 | Procyanidin tetramer B-type | 5.05 | 280 | 1153.2748 | 289.0713 |
| 8 | (−)-Epicatechin a | 5.27 | 280 | 289.0719 | 245.0818 |
| 9 | A-type PA-trimer | 5.52 | 277 | 863.1826 | 289.0714 |
| 10 | B-type PA-trimer | 5.66 | 280 | 865.2001 | 289.0726 |
| 11 | Procyanidin tetramer A-type | 5.76 | 280 | 1151.2525 | 289.0726 |
| 12 | Quercetin-glucoside-rhamnoside-rhamnoside | 6.32 | 350 | 755.2052 | 301.0413 |
| 13 | Quercetin-O-deoxyhexose-O-deoxyhexoside | 6.50 | 350 | 593.1505 | 301.0708 |
| 14 | Quercetin-3-O-rutinoside a | 6.74 | 350 | 609.1461 | 301.0393 |
| 15 | Quercetin-3-O-galacoside a | 6.87 | 355 | 463.0873 | 301.0352 |
| 16 | Quercetin-3-O-glucoside | 7.00 | 355 | 463.0878 | 301.0355 |
| 17 | Procyanidin A-type dimer | 7.14 | 280 | 575.1195 | 289.0717 |
| 18 | Kaempferol-3-O-rutinoside | 7.20 | 350 | 593.1506 | 285.0409 |
| 19 | Quercetin-deoxyhexoside-hexoside | 7.46 | 350 | 609.1449 | 301.0355 |
| 20 | Quercetin-3-O-arabinoside | 7.62 | 350 | 433.0773 | 301.0351 |
| 21 | Quercetin-3-O-rhamnoside a | 7.83 | 350 | 447.0926 | 301.0353 |
| 22 | Keampferol-3-O-arabinoside | 8.43 | 350 | 417.0826 | 285.0401 |
| 23 | Kaempferol glucuronide | 8.53 | 350 | 461.0716 | 285.0386 |
| 24 | Keampferol-3-O-rhamnoside | 8.71 | 350 | 431.0978 | 285.0399 |
| 25 | Isorhamnetin-3-O-rutinoside | 9.39 | 350 | 632.1630 | 315.0505 |
| 26 | Isorhamnetin-3-O-glucoside | 9.99 | 350 | 477.1027 | 315.0497 |
| 27 | Isorhamnetin-3-O-pentoside | 10.69 | 350 | 447.0924 | 315.0504 |
| 28 | Isorhamnetin-3-O-rhamnoside | 11.16 | 350 | 461.1084 | 315.0493 |
| Compounds £ | Aesculus glabra | Aesculus parviflora Walt. |
|---|---|---|
| Protocatechuic-acid-4-glucoside | 0.42 ± 0.01 s * | 0.00 ± 0.00 v |
| Neochlorogenic acid | 1.53 ± 0.06 m | 9.62 ± 0.12 e |
| Procyanidin tetramer A-type | 3.08 ± 0.08 i | 0.50 ± 0.00 r |
| 3-O-p-Coumaroylquinic acid | 3.70 ± 0.03 i | 0.43 ± 0.00 s |
| Chlorogenic acid | 1.60 ± 0.01 l | 1.47 ± 0.01 m |
| Procyanidin B1 | 11.73 ± 0.15 d | 4.47 ± 0.12 h |
| Procyanidin tetramer B-type | 2.11 ± 0.00 j | 0.80 ± 0.00 p |
| (−)-Epicatechin | 51.54 ± 0.22 b | 3.84 ± 0.13 i |
| A-type PA-trimer | 8.90 ± 0.10 e | 0.00 ± 0.00 v |
| B-type PA-trimer | 1.88 ± 0.01 k | 2.07 ± 0.01 j |
| Procyanidin tetramer A-type | 1.91 ± 0.02 j | 0.08 ± 0.01 u |
| Q-glu-rha-rha | 0.73 ± 0.00 q | 0.00 ± 0.00 v |
| Quercetin-O-deoxyhexose-O-deoxyhexoside | 0.78 ± 0.00 pq | 0.00 ± 0.00 v |
| Quercetin-3-O-rutinoside | 0.57 ± 0.00 r | 4.07 ± 0.12 h |
| Quercetin-3-O-galacoside | 4.05 ± 0.11 h | 0.42 ± 0.08 s |
| Quercetin-3-O-glucoside | 1.31 ± 0.01 n | 0.00 ± 0.00 v |
| Procyanidin A-type dimer | 1.64 ± 0.10 l | 0.00 ± 0.00 v |
| Kaempferol-3-O-rutinoside | 0.85 ± 0.01 p | 0.24 ± 0.09 t |
| Quercetin-deoxyhexoside-hexoside | 1.92 ± 0.13 jk | 0.00 ± 0.00 v |
| Quercetin-3-O-arabinoside | 9.14± 0.21 e | 0.00 ± 0.00 v |
| Quercetin-3-O-rhamnoside | 7.31 ± 0.10 f | 15.40 ± 0.15 d |
| Keampferol-3-O-arabinoside | 1.54 ± 0.03 m | 0.00 ± 0.00 v |
| Kaempferol glucuronide | 0.16 ± 0.01 t | 0.00 ± 0.00 v |
| Keampferol-3-O-rhamnoside | 1.19 ± 0.00 o | 1.06 ± 0.07 o |
| Isorhamnetin-3-O-rutinoside | 0.38 ± 0.00 s | 0.00 ± 0.00 v |
| Isorhamnetin-3-O-glucoside | 5.52 ± 0.10 g | 0.00 ± 0.00 v |
| Isorhamnetin-3-O-pentoside | 2.95 ± 0.02 ij | 0.00 ± 0.00 v |
| Isorhamnetin-3-O-rhamnoside | 1.14 ± 0.03 o | 0.00 ± 0.00 v |
| Procyanidin polymers | 37.04 ± 0.20 c | 81.87 ± 0.25 a |
| Total | 166.62 A | 126.34 B |
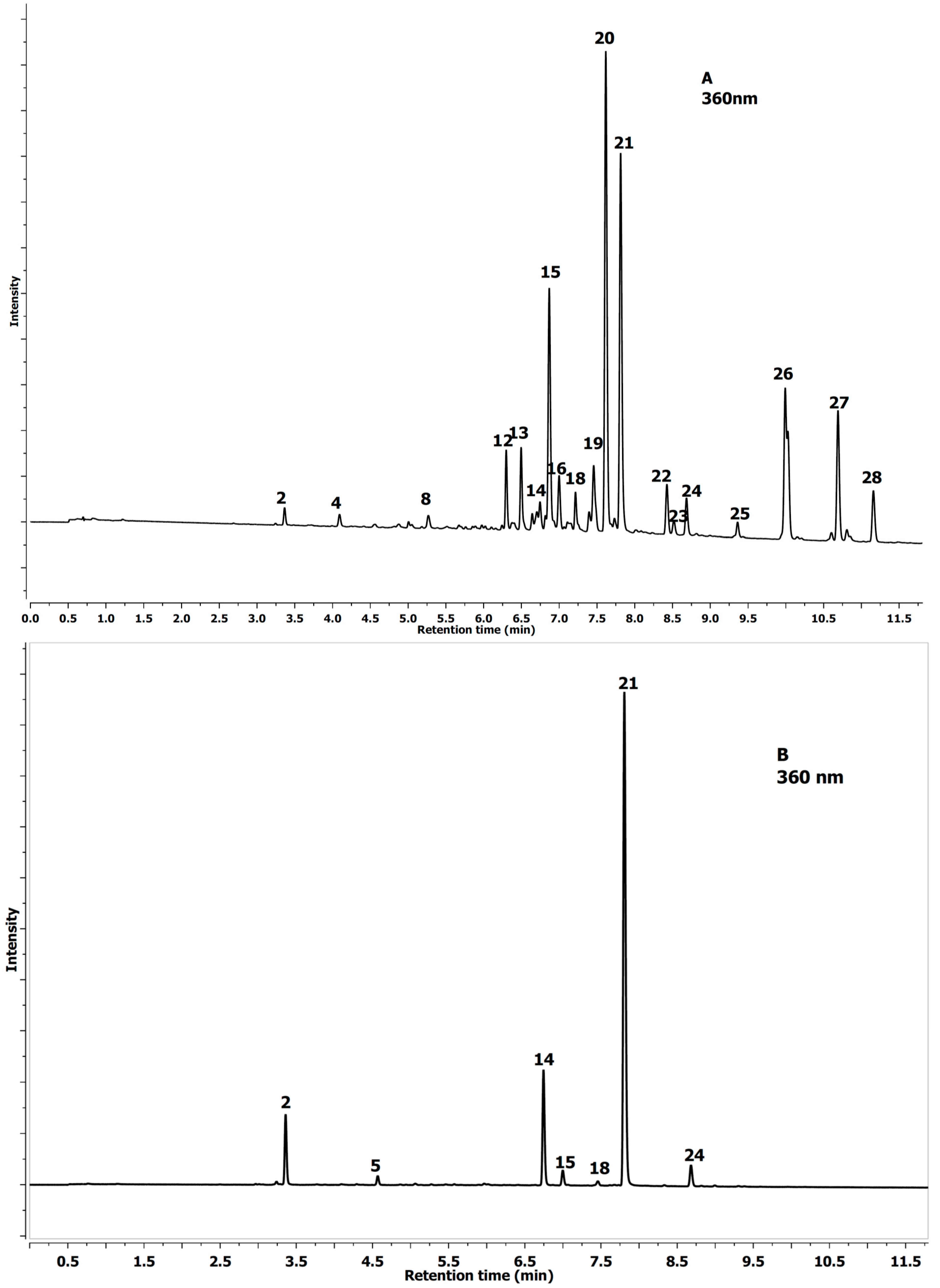
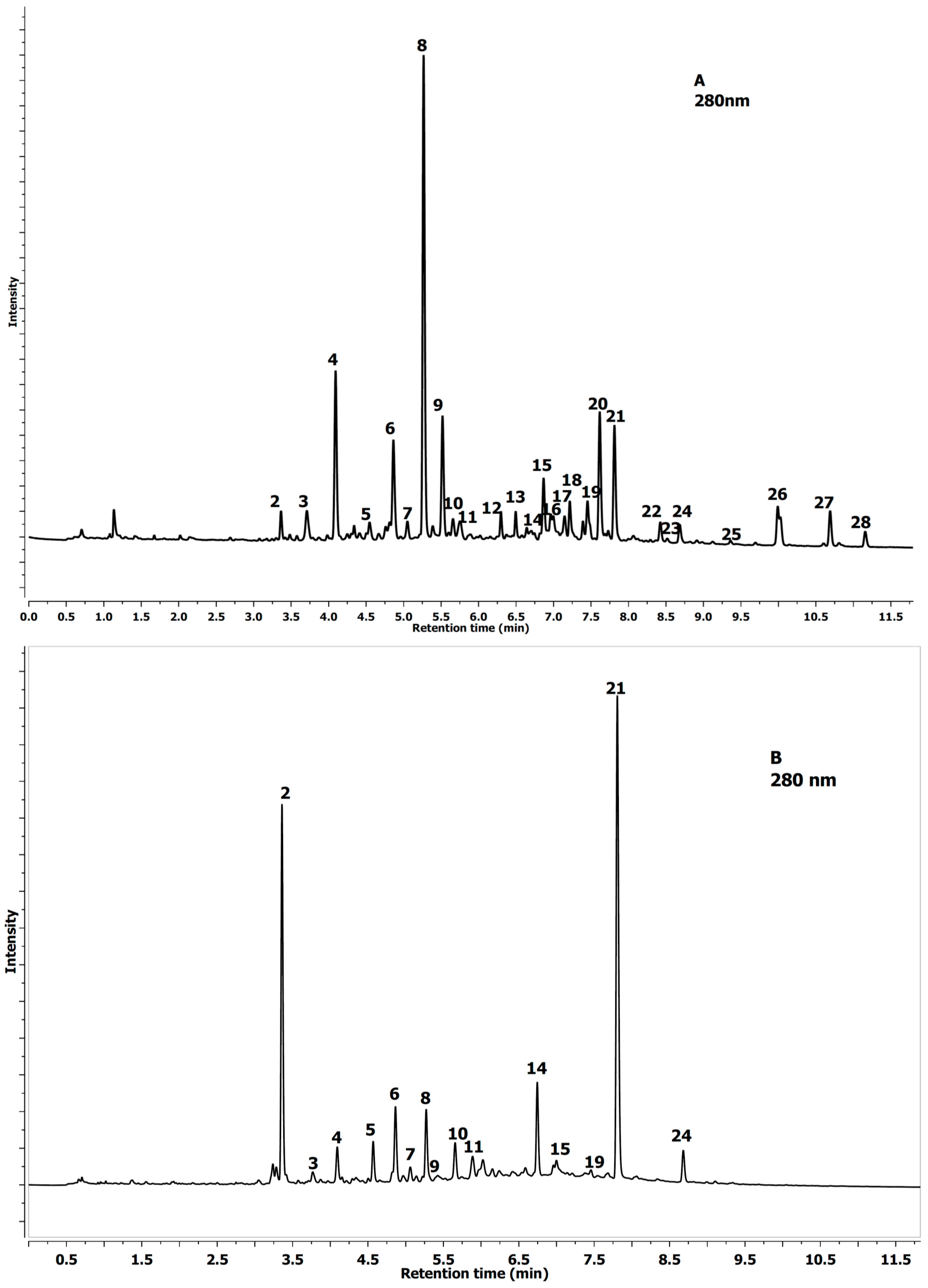
2.2. Quantification of Phenolics in Aesculus Leaves
3. Experimental Section
3.1. Reagent and Standards
3.2. Plant Material
3.3. Extraction Procedure
3.4. Identification of Polyphenols by the Liquid Chromatography-Mass Spectrometry (LC-MS) Method
3.5. Analysis of Proanthocyanidins by the Phloroglucinolysis Method
3.6. Statistical Analysis
4. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Koes, R.E.; Quattrocchio, F.; Mol, J.N.M. The flavonoid biosynthetic pathway in plants: Function and evolution. BioEssays 1994, 16, 123–132. [Google Scholar] [CrossRef]
- Elio, G.W.M.; Schijlen, C.H.; de Vos, R.; van Tunen, A.J.; Bovy, A.G. Modification of flavonoid biosynthesis in crop plants. Phytochemistry 2004, 65, 2631–2648. [Google Scholar] [CrossRef] [PubMed]
- Feucht, W. The localization of phenols at the cellular and tissue level. Acta Hort. 1994, 381, 803–815. [Google Scholar]
- Aver’yanov, A.A.; Lapikova, V.P. Participation of active forms of oxygen in the mechanism of ferulic acid toxicity. Biol. Bull. 1985, 4, 352–357. [Google Scholar]
- Satisha, J.; Doshi, P.; Adsule, P.G. Influence of rootstocks on changing the pattern of phenolic compounds in thompson seedless grapes and its relationship to the incidence of powdery mildew. Turk. J. Agric. For. 2008, 32, 1–9. [Google Scholar]
- Tahvannainen, J.; Helle, E.; Julkunen-Tinto, R.; Lavola, A. Phenolic glycosides govern the food selection pattern in willow feeding leaf beetles. Oecologia 1985, 67, 52–56. [Google Scholar] [CrossRef]
- Bennett, R.N.; Wallsgrove, R.M. Secondary metabolites in plant defence mechanisms. New Phytol. 1994, 127, 617–633. [Google Scholar] [CrossRef]
- Grayer, R.J.; Kimmins, F.M.; Padgham, D.E.; Harborne, J.B.; Ranga, D.V. Condensed tannins levels and resistance of groundnuts against Aphis craccivora. Phytochemistry 1992, 31, 3795–3780. [Google Scholar] [CrossRef]
- Michalek, S.; Mayr, U.; Treutter, D.; Lux-Endrich, A.; Gutmann, M.; Feucht, W.; Geibel, M. Role of flavan-3-ols in resistance of apple trees to Venturia inaequalis. Acta Hortic. 1999, 484, 535–539. [Google Scholar]
- Xia, N.H.; Thurland, N.J.; Gadek, P.A. Hippocastanaceae. Flora China 2010, 12, 1–4. [Google Scholar]
- Xiang, Q.Y.; Crawford, D.J.; Wolfe, A.D.; Tang, Y.C.; DePamphilis, C.W. Origin and biogeography of Aesculus L. (Hippocastanaceae): A molecular phylogenetic perspective. Evolution 1998, 52, 988–997. [Google Scholar] [CrossRef]
- Merz, R.W. Ohio buckeye (Aesculus glabra Willd.). In Silvics of Forest Trees of the United States; Fowells, H.A., Ed.; Comp. U.S. Department of Agriculture, Agriculture Handbook 271: Washington, DC, USA, 1965; pp. 75–77. [Google Scholar]
- Rudolf, P.O. Aesculus L. Buckeye, Horsechestnut. In Seeds of Woody Plants in the United States; U.S. Department of Agriculture, Agriculture Handbook 450: Washington, DC, USA, 1974; pp. 195–200. [Google Scholar]
- Zhang, Z.; Li, S.; Lian, X.Y. An overview of genus Aesculus L.: Ethnobotany, phytochemistry and pharmacological activities. Pharm. Crops 2010, 1, 24–51. [Google Scholar] [CrossRef]
- Simova-Tosic, D.; Filev, S. Contribution to the knowledge of leaf miners of horse chestnut. Zast. Bilja 1985, 36, 235–239. [Google Scholar]
- Freise, J.F.; Heitland, W. Bionomics of the horse chestnut leaf miner Cameraria ohridella DESCHKA & DIMIC 1986, a pest on Aesculus hippocastanum in Europe (Insecta, Lepidoptera, Gracillariidae). Senckenberg. Biol. 2004, 84, 61–80. [Google Scholar]
- Girardoz, S.; Quicke, D.L.J.; Kenis, M. Factors favouring the development and maintenance of outbreaks in an invasive leaf miner Cameraria ohridella (Lepidoptera: Gracillariidae): A life table study. Agric. For. Enomol. 2007, 9, 141–158. [Google Scholar] [CrossRef]
- Girardoz, S.; Tomov, R.; Eschen, R.; Quicke, D.L.J.; Kenis, M. Two methods of assessing the mortality factors affecting the larvae and pupae of Cameraria ohridella in the leaves of Aesculus hippocastanum in Switzerland and Bulgaria. Bull. Entomol. Res. 2007, 97, 445–453. [Google Scholar] [PubMed]
- Raimondo, F.; Ghirardella, L.A.; Nardini, A.; Salleo, S. Impact of the leaf miner Cameraria ohridella on photosynthesis, water relations and hydraulics of Aesculus hippocastanum leaves. Trees 2003, 17, 376–382. [Google Scholar]
- Thalmann, C.; Freise, J.; Heitland, W.; Bacher, S. Effects of defoliation by horse chestnut leaf miner Cameraria ohridella on reproduction in Aesculus hippocastanum. Trees 2003, 17, 383–388. [Google Scholar] [CrossRef]
- Freise, J.F.; Heitland, W.; Sturm, A. Das physiologische Wirtspflanzenspektrum der Rosskastanien-Miniermotte, Cameraria ohridella Deschka & Dimic (Lepidoptera: Gracillariidae). Nachrichtenbl. Deut. Pflanzenschutzd. 2003, 55, 209–211. [Google Scholar]
- Péré, C.; Augustin, S.; Turlings, T.C.J.; Kenis, M. The invasive alien leaf miner, Cameraria ohridella and the native tree, Acer pseudoplatanus: A fatal attraction? Agric. For. Entomol. 2010, 12, 151–159. [Google Scholar] [CrossRef]
- Straw, N.A.; Tilbury, C. Host plants of the horsechestnut leaf-miner (Cameraria ohridella), and the rapid spread of the moth in the UK 2002–2005. Arboric. J. 2006, 29, 83–99. [Google Scholar] [CrossRef]
- Dudek-Makuch, M.; Matławska, I. Flavonoids from the flowers of Aesculus hippocastanum. Acta Pol. Pharm. 2011, 68, 403–408. [Google Scholar] [PubMed]
- Kapusta, I.; Janda, B.; Szajwaj, B. Flavonoids in horse chestnut (Aesculus hippocastanum) seeds and powdered waste water byproducts. J. Agric. Food Chem. 2007, 55, 8485–8490. [Google Scholar] [CrossRef] [PubMed]
- Hübner, G.; Wray, V.; Nahrstedt, A. Flavonol oligosaccharides from the seeds of Aesculus hippocastanum. Planta Med. 1999, 65, 636–642. [Google Scholar] [CrossRef] [PubMed]
- Oszmiański, J.; Kalisz, S.; Wojdyło, A. The content of phenolic compounds in leaf tissues of white (Aesculus hippocastanum L.) and red horse chestnut (Aesculus carea H.) colonized by the horse chestnut leaf miner (Cameraria ohridella Deschka & Dimić). Molecules 2014, 19, 14625–14636. [Google Scholar] [CrossRef]
- Wei, F.; Ma, S.C.; Ma, L.Y.; But, P.P.; Lin, R.C.; Khan, I.A. Antiviral flavonoids from the seeds of Aesculus chinensis. J. Nat. Prod. 2004, 67, 650–653. [Google Scholar] [CrossRef]
- Curir, P.; Galeotti, F.; Dolci, M.; Barile, E.; Lanzotti, V. Pavietin, a coumarin from Aesculus pavia with antifungal activity. J. Nat. Prod. 2007, 10, 1668–1671. [Google Scholar] [CrossRef]
- Niederleitner, S.; Zinkernagel, V.; Treutter, D.; Feucht, W. Accumulation of flavonols in cherry leaves after infection by the fungus Blumeriella jaapi. Acta Hortic. 1994, 381, 767–770. [Google Scholar]
- Andreotti, C.; Costa, G.; Treutter, D. Composition of phenolic compounds in pear leaves as affected by genetics, ontogenesis and the environment. Sci. Hortic. 2006, 109, 130–137. [Google Scholar] [CrossRef]
- Schultz, J.C. Impact of Variable Plant Defensive Chemistry on Susceptibility of Insects to Natural Enemies; Hein, P.A., Ed.; American Chemistry Society: Washington, DC, USA, 1983; pp. 37–54. [Google Scholar]
- Agrawal, A.A.; Hastings, A.P.; Johnson, M.T.J.; Maron, J.L.; Salminen, J.P. Insect herbivores drive real-time ecological and evolutionary change in plant populations. Science 2012, 338, 113–116. [Google Scholar] [CrossRef] [PubMed]
- Feeny, P.P. Seasonal changes in oak leave tannins and nutrients as a cause of spring feeding by winter moth caterpillars. Ecology 1970, 51, 565–581. [Google Scholar] [CrossRef]
- Forkner, R.E.; Marquis, R.J.; Lill, J.T. Feeny revisited: Condensed tannins as anti-herbivore defences in leaf-chewing herbivore communities of Quercus. Ecol. Entomol. 2004, 29, 174–187. [Google Scholar] [CrossRef]
- Haslam, E. Vegetable tannins—Lessons of a phytochemical lifetime. Phytochemistry 2007, 68, 2713–2721. [Google Scholar] [CrossRef] [PubMed]
- Feeny, P.P. Inhibitory effect of oak leaf tannins on the hydrolysis of proteins by trypsin. Phytochemistry 1969, 8, 2119–2126. [Google Scholar] [CrossRef]
- Robbins, C.T.; Hanley, T.A.; Hagerman, A.E.; Hjeljord, O.; Baker, D.L.; Schwartz, C.C.; Mautz, W.W. Role of tannins in defending plants against ruminants: Reduction in protein availability. Ecology 1987, 68, 98–107. [Google Scholar] [CrossRef]
- Isman, M.B.; Duffey, S.S. Toxicity of tomato phenolic compounds to the fruit form Heliothis zea. Entomol. Exp. App. 1982, 31, 370–376. [Google Scholar] [CrossRef]
- Oszmiań>ski, J.; Kolniak-Ostek, J.; Wojdyło, A. Application of ultra performance liquid chromatography-photodiode detector-quadrupole/time of flight-mass spectrometry (UPLC-PDA-Q/TOF-MS) method for the characterization of phenolic compounds of Lepidium sativum L. sprouts. Eur. Food Res. Technol. 2013, 236, 699–706. [Google Scholar] [CrossRef]
- Kolniak-Ostek, J.; Oszmiański, J.; Rutkowski, K.P.; Wojdyło, A. Effect of 1-methylcyclopropene postharvest treatment apple and storage on the cloudy juices properties. LWT—Food Sci. Technol. 2014, 59, 1166–1176. [Google Scholar] [CrossRef]
- Sample Availability: Samples of the analysed leaves are available from the authors.
© 2015 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license ( http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Oszmiański, J.; Kolniak-Ostek, J.; Biernat, A. The Content of Phenolic Compounds in Leaf Tissues of Aesculus glabra and Aesculus parviflora Walt. Molecules 2015, 20, 2176-2189. https://doi.org/10.3390/molecules20022176
Oszmiański J, Kolniak-Ostek J, Biernat A. The Content of Phenolic Compounds in Leaf Tissues of Aesculus glabra and Aesculus parviflora Walt. Molecules. 2015; 20(2):2176-2189. https://doi.org/10.3390/molecules20022176
Chicago/Turabian StyleOszmiański, Jan, Joanna Kolniak-Ostek, and Agata Biernat. 2015. "The Content of Phenolic Compounds in Leaf Tissues of Aesculus glabra and Aesculus parviflora Walt." Molecules 20, no. 2: 2176-2189. https://doi.org/10.3390/molecules20022176