
Protective Effects of Lycium barbarum Polysaccharide on 6-OHDA-Induced Apoptosis in PC12 Cells through the ROS-NO Pathway

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results and Discussion
2.1. Results
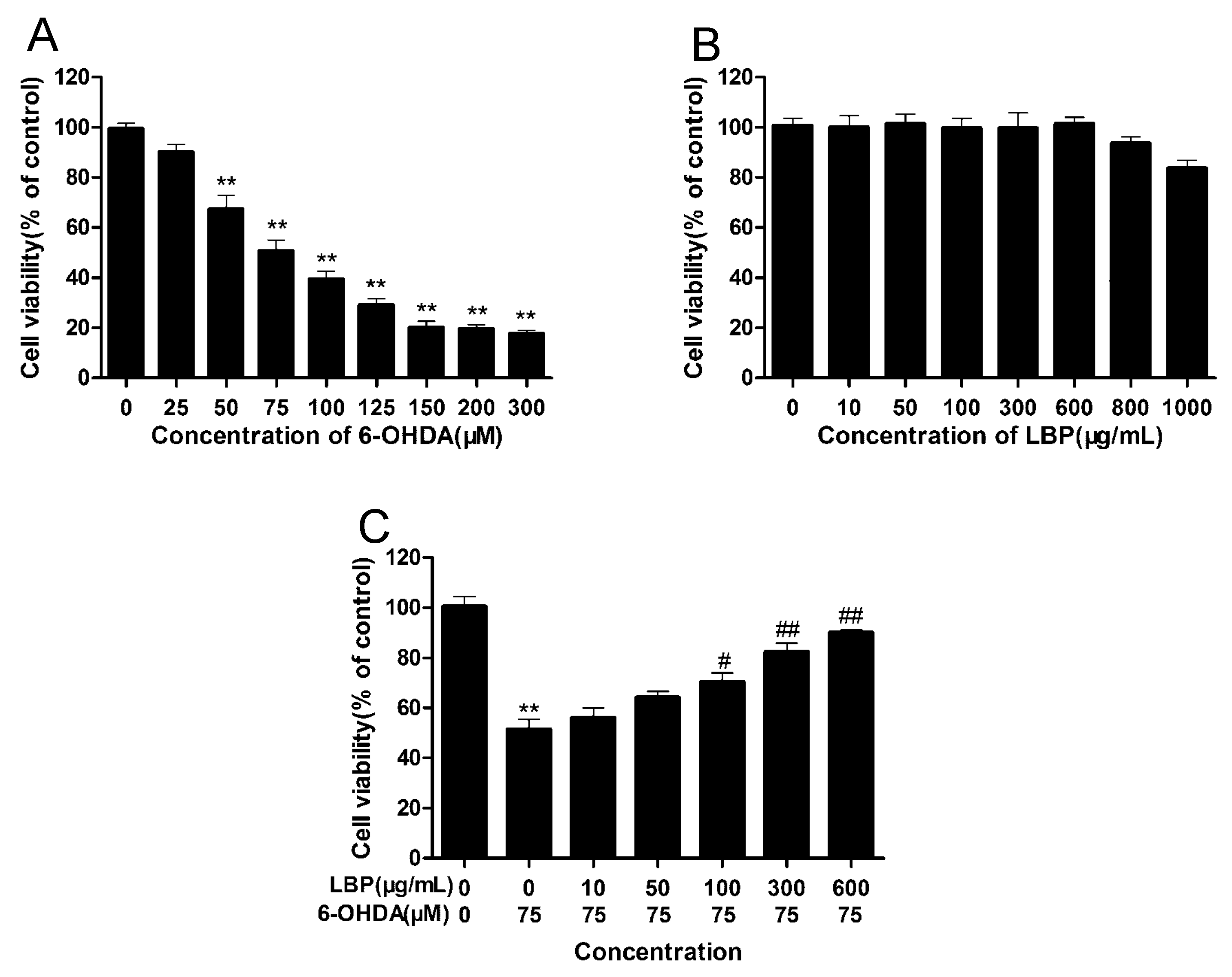
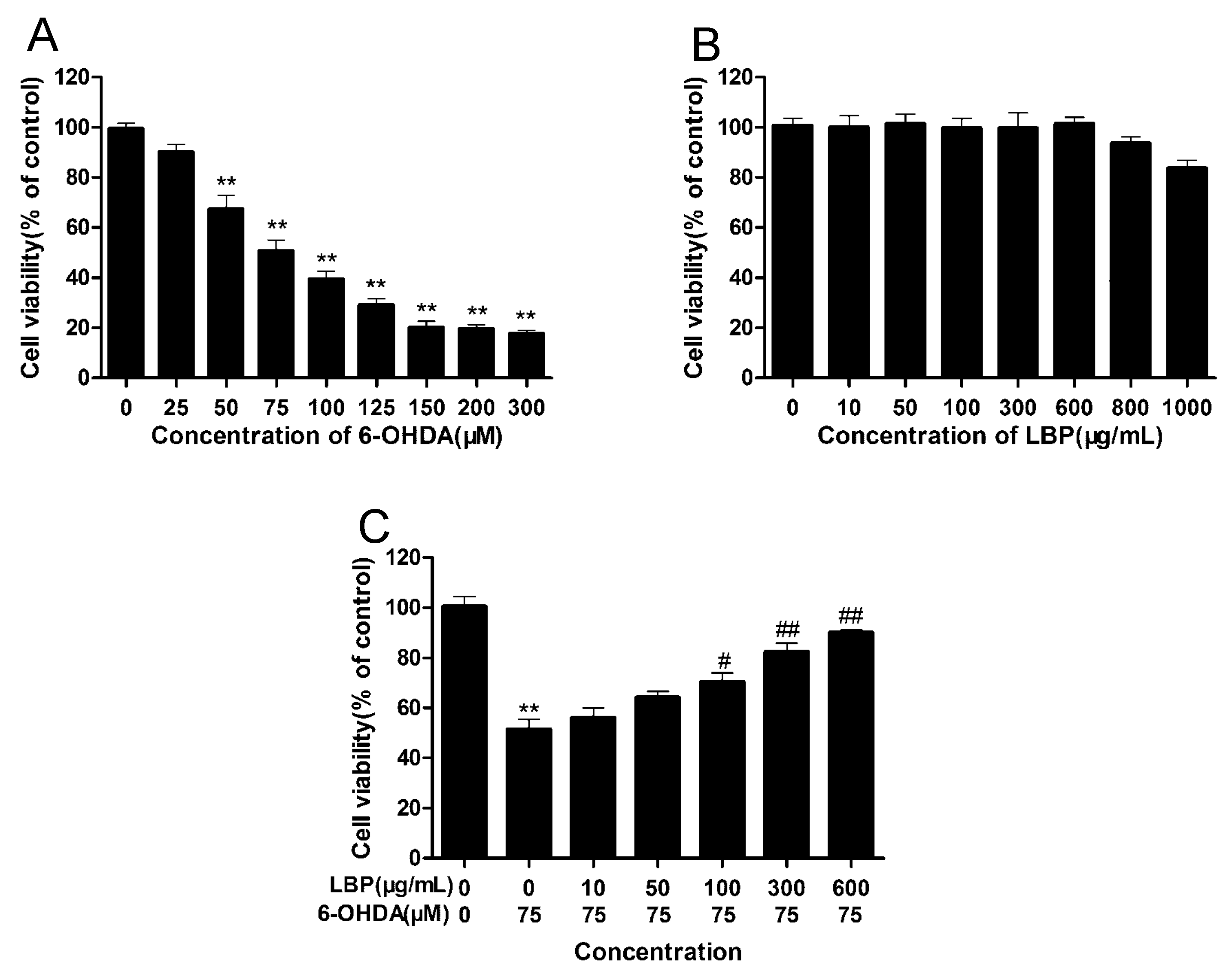
2.1.1. LBP Prevents 6-OHDA -Induced Apoptosis of pc12cells
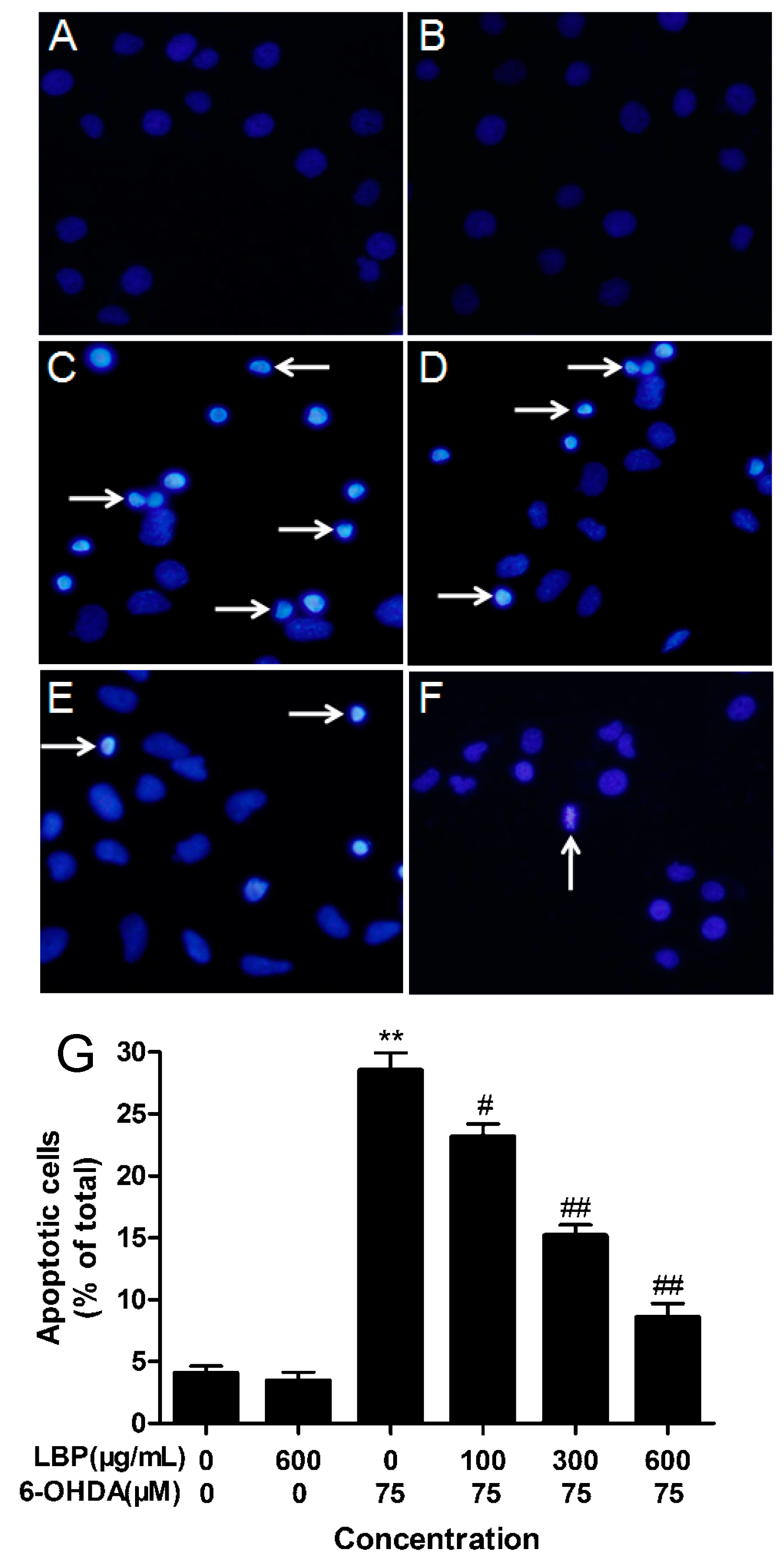
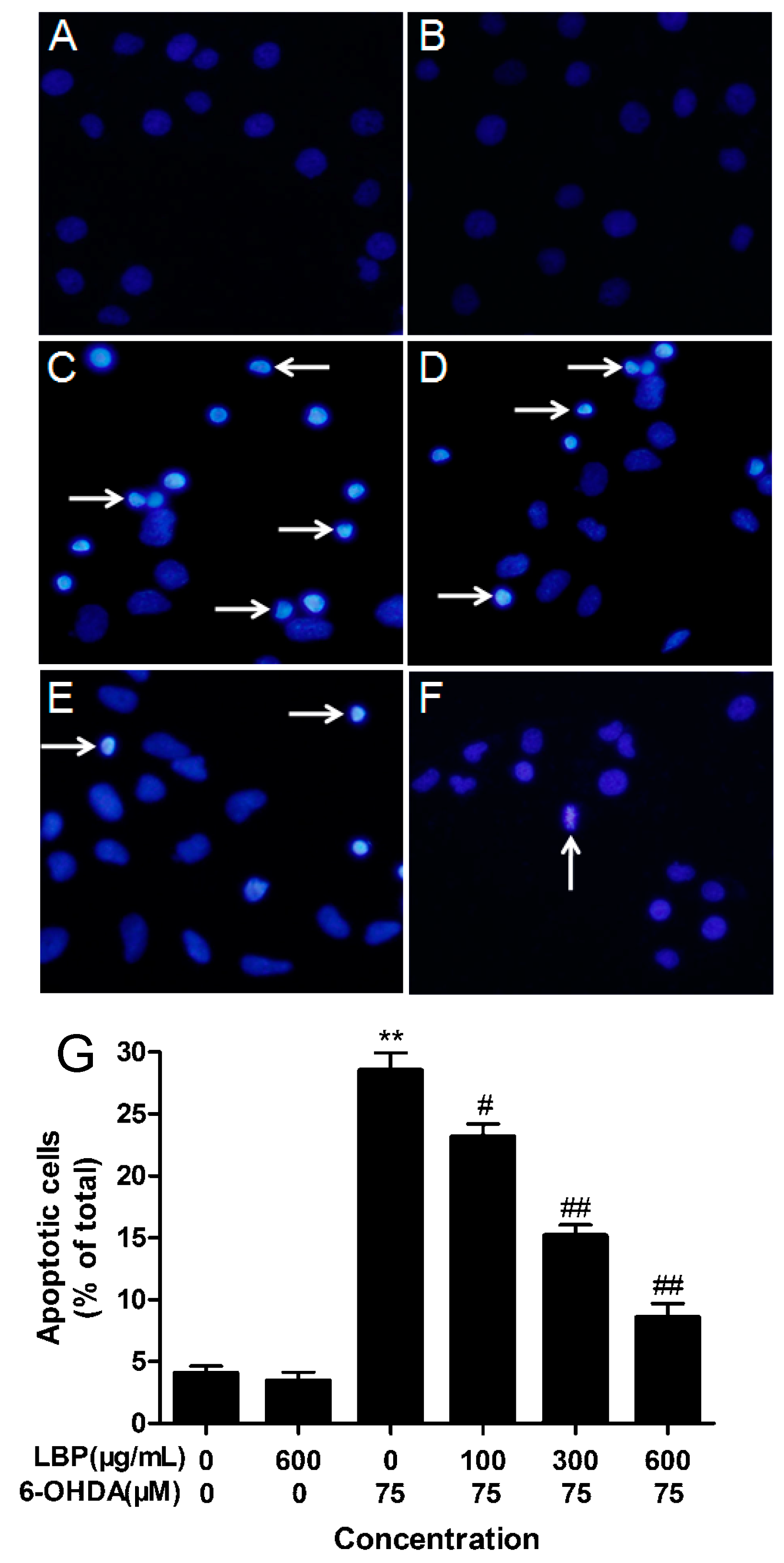
2.1.2. LBP Rescues 6-OHDA -Induced Changes in Nuclear Morphology
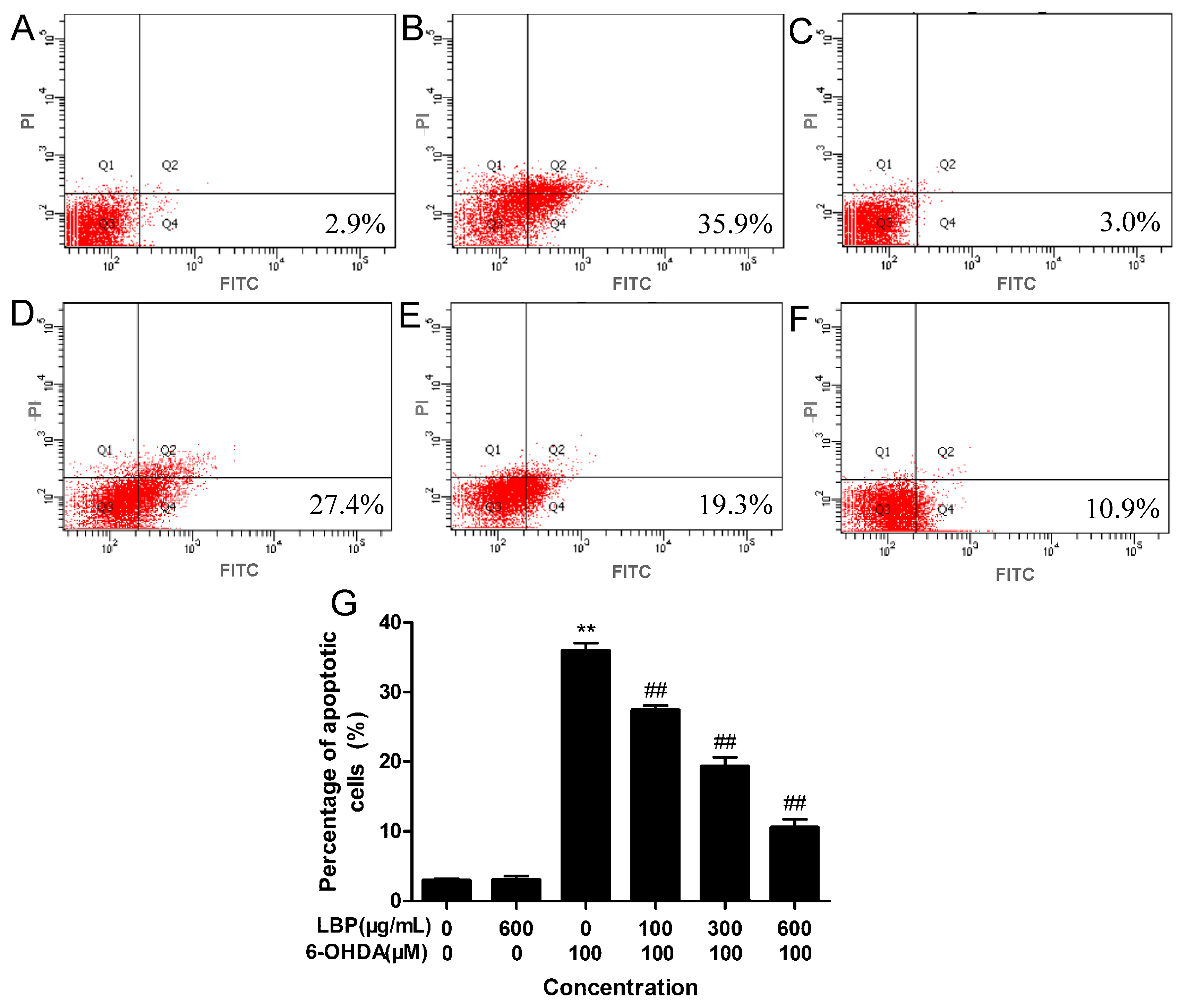
2.1.3. LBP Rescues 6-OHDA -Induced Apoptosis
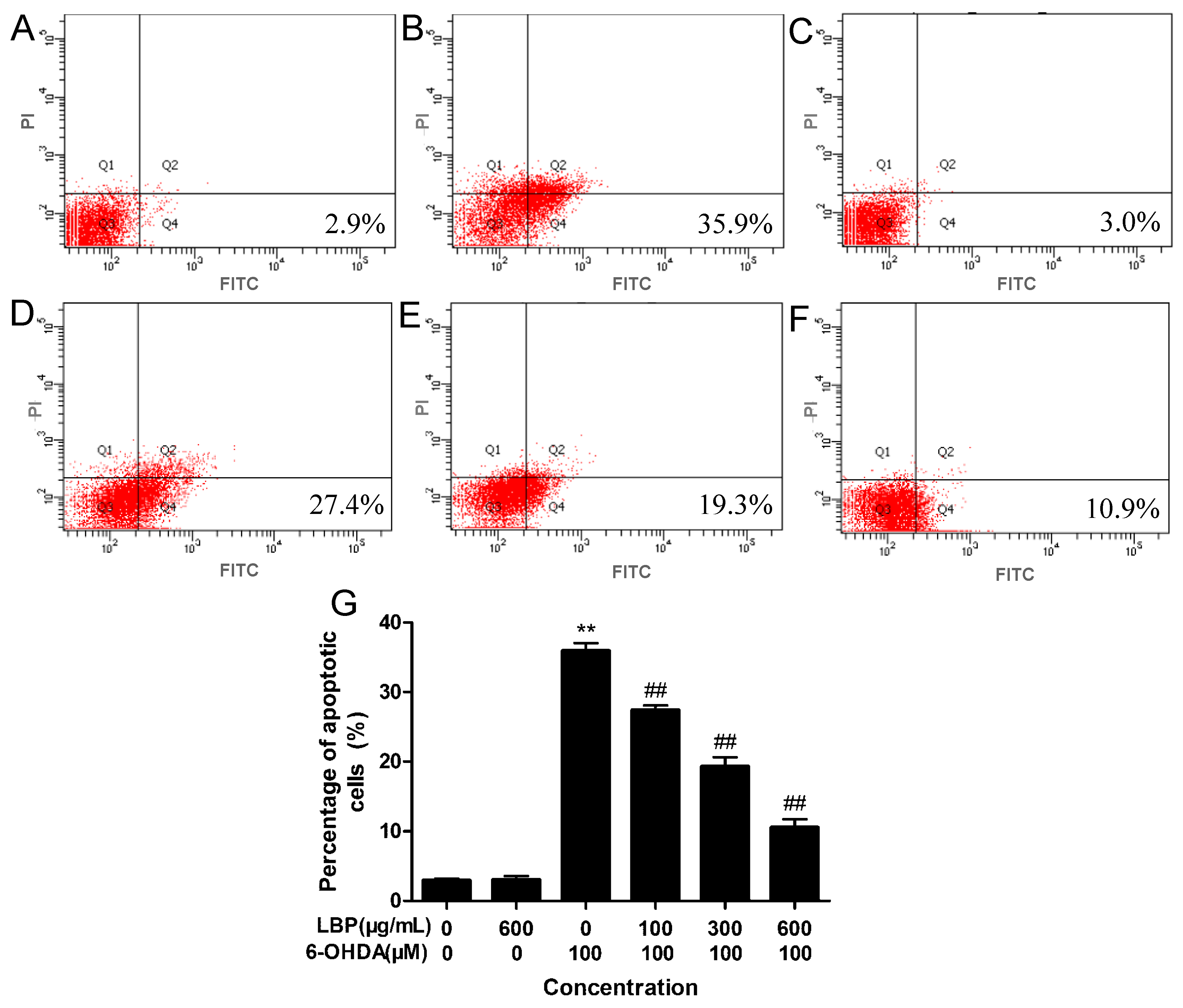
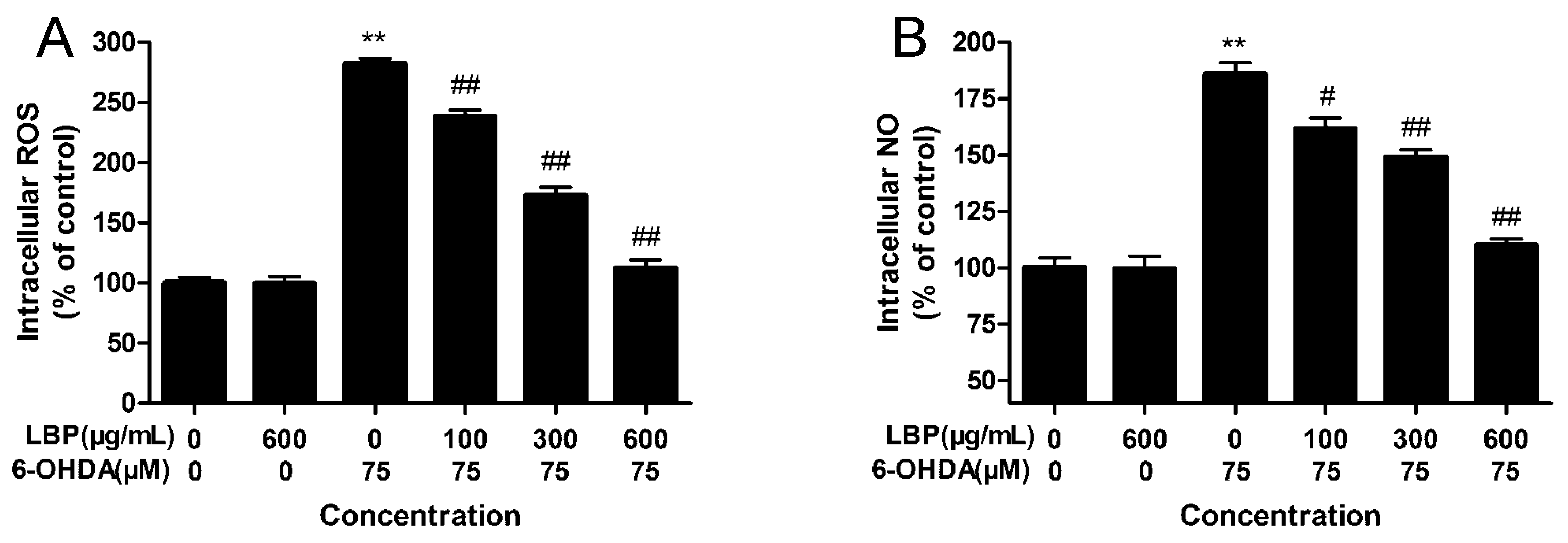
2.1.4. Effects of LBP on Intracellular ROS Levels
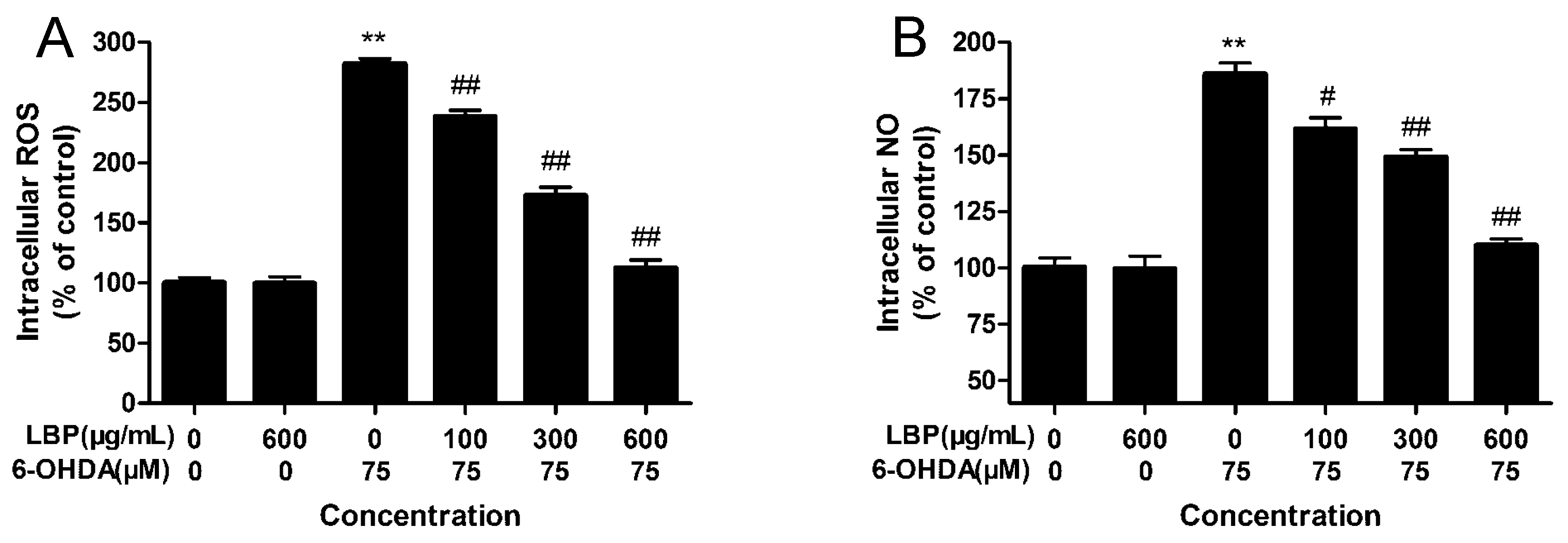
2.1.5. Effects of LBP on Intracellular NO Levels
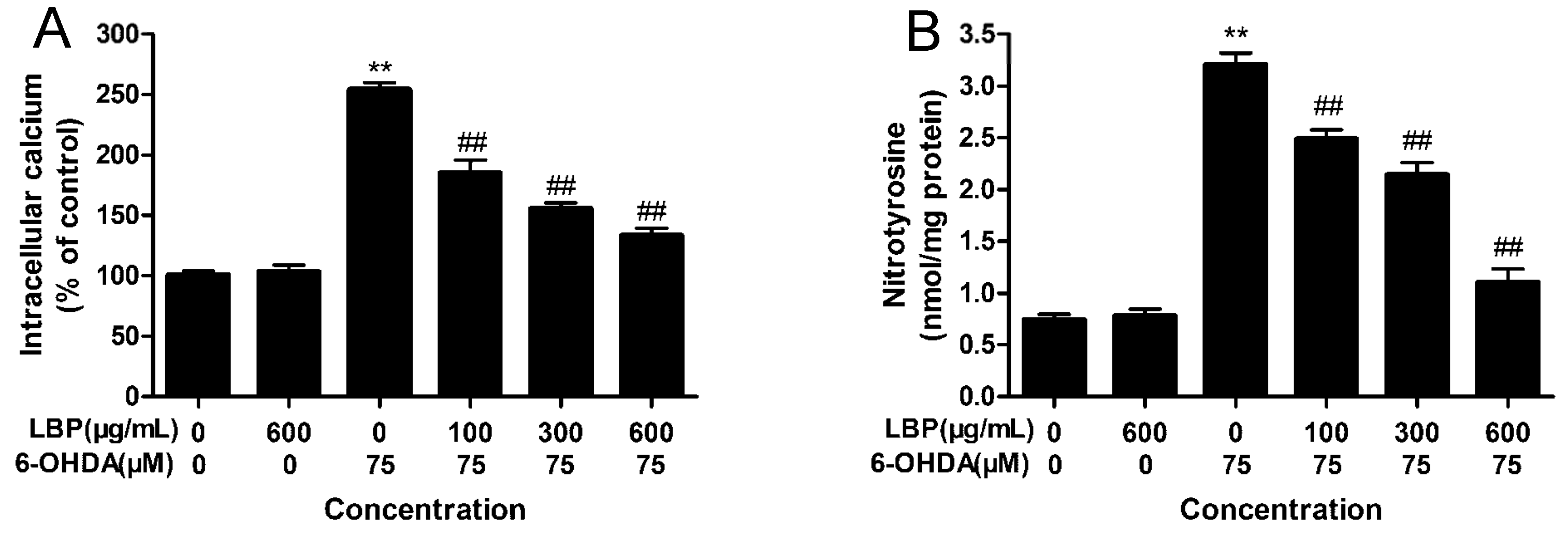
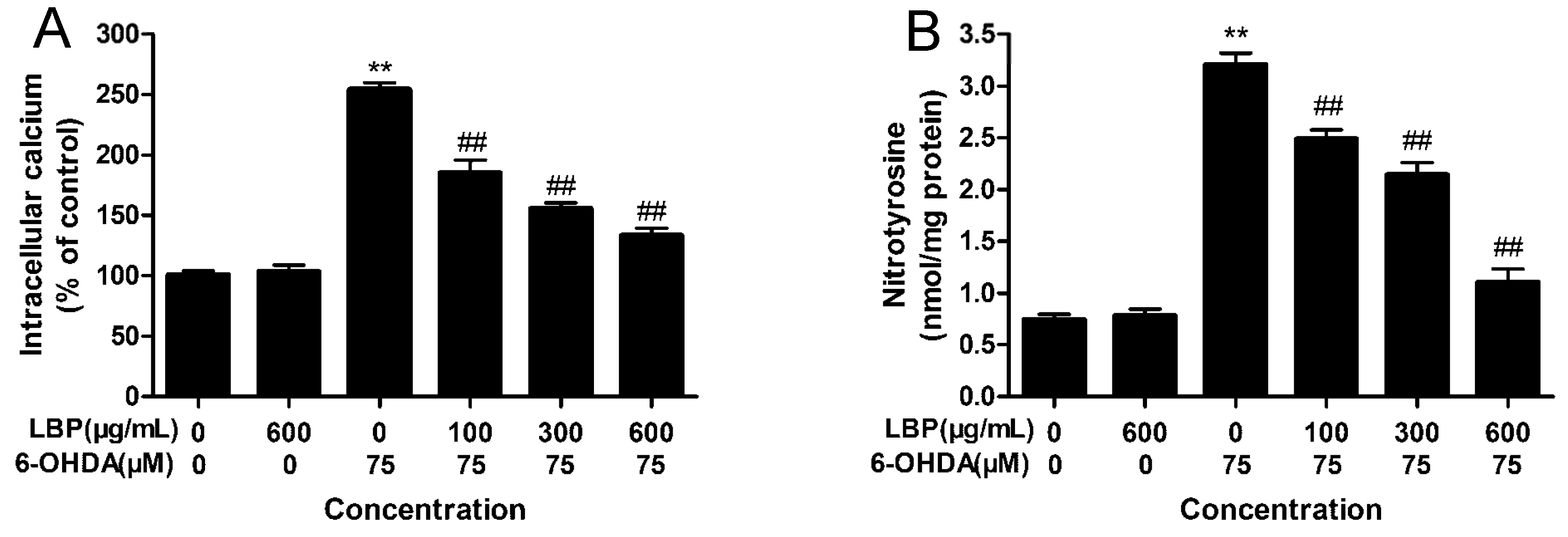
2.1.6. LBP Inhibits 6-OHDA -Induced Elevation in [Ca2+]i
2.1.7. LBP Inhibits the 6-OHDA -Induced Elevation in Protein-Bound 3-NT
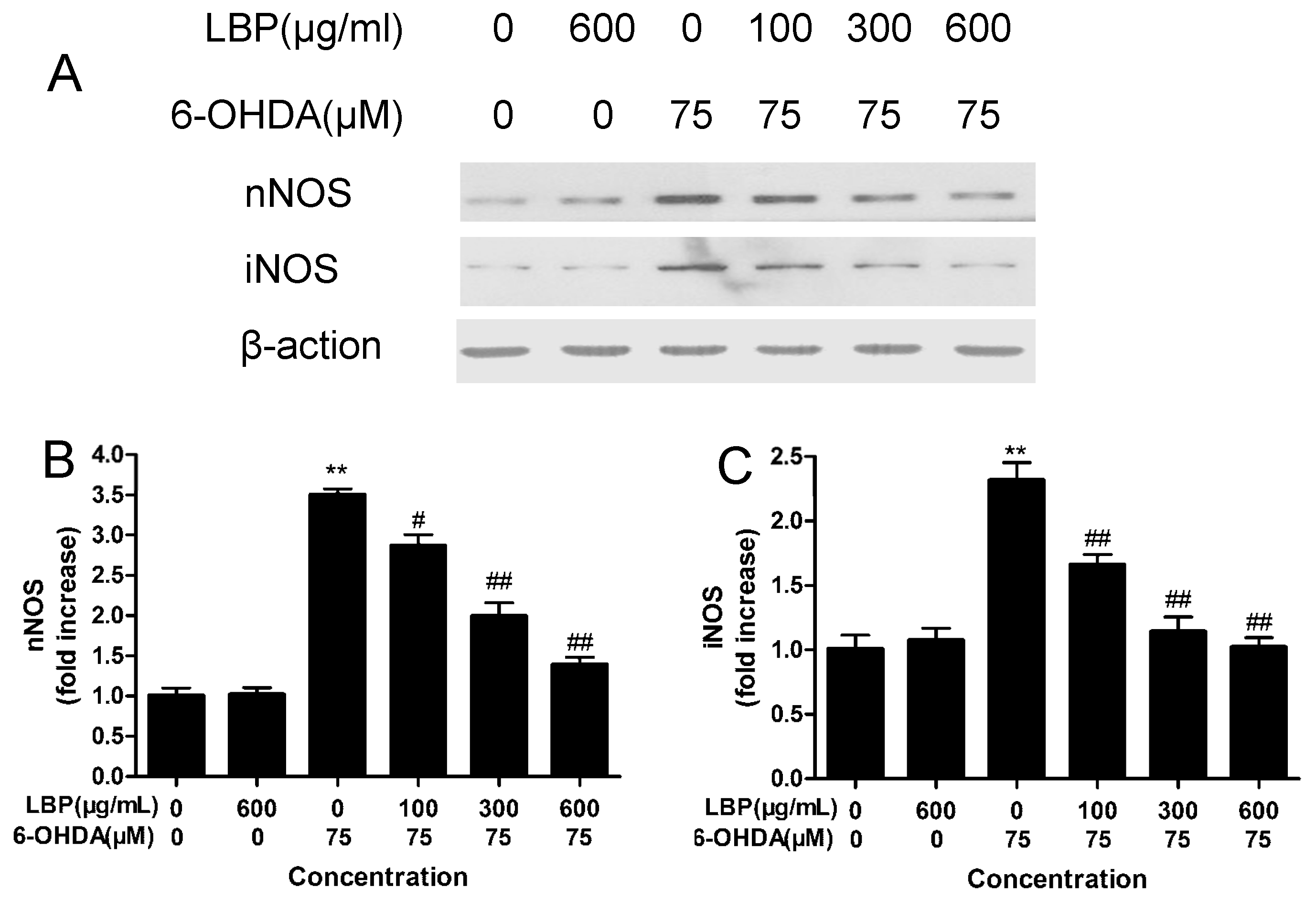
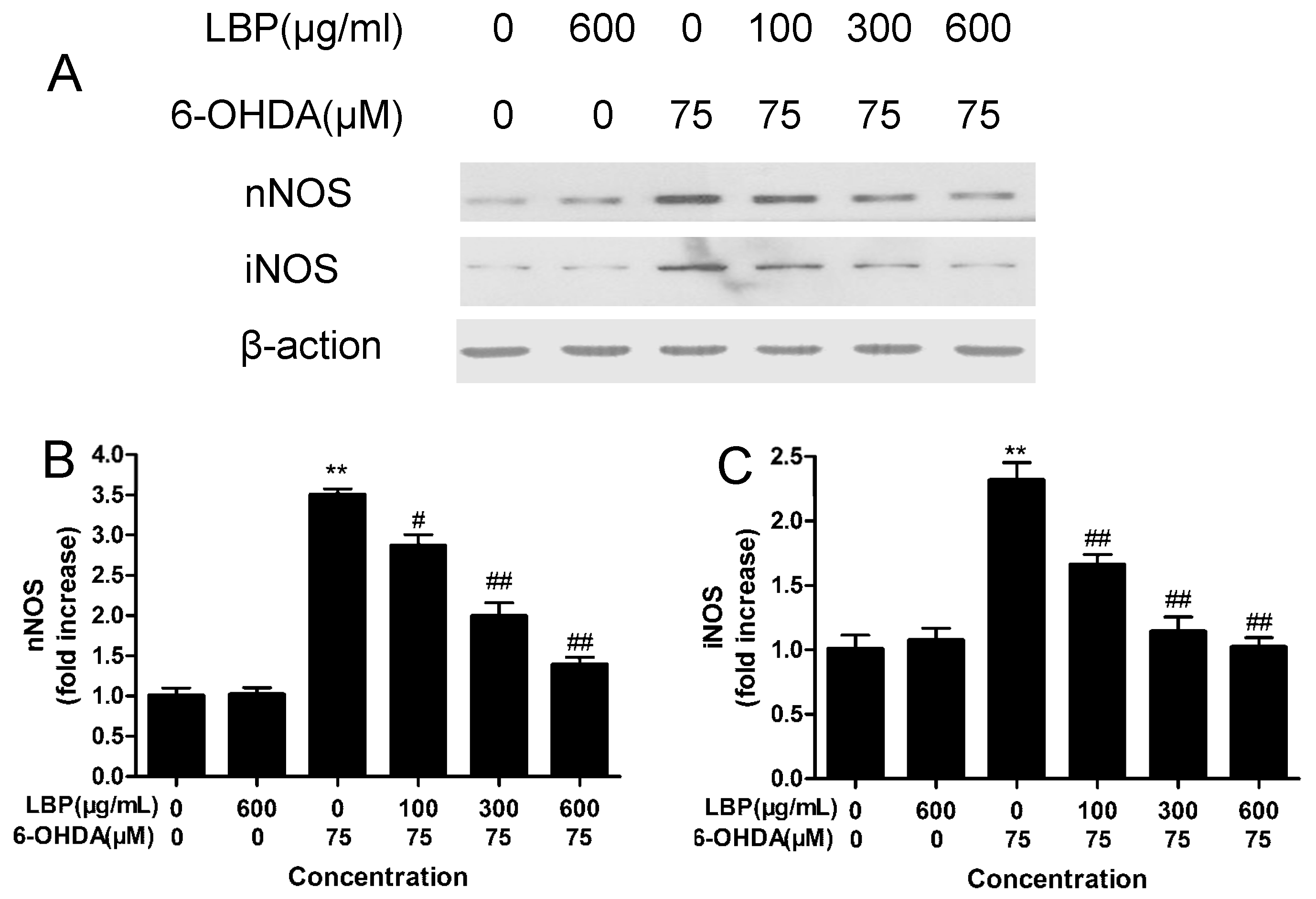
2.1.8. Effects of LBP on the Expression of nNOS and iNOS
2.1.9. Effects of LBP on Caspase 3 and Caspase 9 Activities
2.2. Discussion
3. Experimental Section
3.1. Preparation of Lycium Barbarum Polysaccharide
3.2. Reagents
3.3. Primary Culture of Rat Cortical Neurons
3.4. PC12 Cell Culture and Treatment
3.5. Assessment of Cell Viability
3.6. Morphological Changes
3.7. Flow Cytometric Analysis
3.8. Measurement of Intracellular ROS
3.9. Measurement of Intracellular NO
3.10. LBP Inhibits 6-OHDA -Induced Elevation in [Ca2+]i
3.11. Measurement of Protein-Bound 3-NT
3.12. Western Blot Analysis of nNOS and iNOS
3.13. Statistical Analysis
4. Conclusions
Supplementary Materials
Acknowledgments
Author Contributions
Conflicts of Interest
References
- De Lau, L.M.; Breteler, M.M. Epidemiology of Parkinson’s disease. Lancet Neurol. 2006, 5, 525–535. [Google Scholar] [CrossRef]
- Hornykiewicz, O.; Kish, S.J. Biochemical pathophysiology of Parkinson’s disease. Adv. Neurol. 1987, 45, 19–34. [Google Scholar] [PubMed]
- Wang, T.; Gu, J.; Wu, P.F.; Wang, F.; Xiong, Z.; Yang, Y.J.; Wu, W.N.; Dong, L.D.; Chen, J.G. Protection by tetrahydroxystilbene glucoside against cerebral ischemia: Involvement of JNK, SIRT1, and NF-kappaB pathways and inhibition of intracellular ROS/RNS generation. Free Radic. Biol. Med. 2009, 47, 229–240. [Google Scholar] [CrossRef] [PubMed]
- Chung, C.Y.; Khurana, V.; Auluck, P.K.; Tardiff, D.F.; Mazzulli, J.R.; Soldner, F.; Baru, V.; Lou, Y.; Freyzon, Y.; Cho, S.; et al. Identification and rescue of alpha-synuclein toxicity in Parkinson patient-derived neurons. Science 2013, 342, 983–987. [Google Scholar] [CrossRef] [PubMed]
- Chaturvedi, R.K.; Beal, M.F. Mitochondria targeted therapeutic approaches in Parkinson’s and Huntington’s diseases. Mol. Cell. Neurosci. 2013, 55, 101–114. [Google Scholar] [CrossRef] [PubMed]
- Beal, M.F. Therapeutic approaches to mitochondrial dysfunction in Parkinson’s disease. Parkinsonism Relat. Disord. 2009, 15 (Suppl. 3), S189–S194. [Google Scholar]
- Guo, S.; Bezard, E.; Zhao, B. Protective effect of green tea polyphenols on the SH-SY5Y cells against 6-OHDA induced apoptosis through ROS-NO pathway. Free Radic. Biol. Med. 2005, 39, 682–695. [Google Scholar] [CrossRef] [PubMed]
- Jellinger, K.A.; Kienzl, E.; Rumpelmaier, G.; Paulus, W.; Riederer, P.; Stachelberger, H.; Youdim, M.B.; Ben-Shachar, D. Iron and ferritin in substantia nigra in Parkinson’s disease. Adv. Neurol. 1993, 60, 267–272. [Google Scholar] [PubMed]
- Sian, J.; Dexter, D.T.; Lees, A.J.; Daniel, S.; Agid, Y.; Javoy-Aqid, F.; Jenner, P.; Marsden, C.D. Alterations in glutathione levels in Parkinson’s disease and other neurodegenerative disorders affecting basal ganglia. Ann. Neurol. 1994, 36, 348–355. [Google Scholar] [CrossRef] [PubMed]
- Smith, M.P.; Cass, W.A. GDNF reduces oxidative stress in a 6-hydroxydopamine model of Parkinson’s disease. Neurosci. Lett. 2007, 412, 259–263. [Google Scholar] [CrossRef] [PubMed]
- Ju, M.S.; Lee, P.; Kim, H.G.; Lee, K.Y.; Hur, J.; Cho, S.H.; Sung, S.H. Protective effects of standardized Thuja orientalis leaves against 6-hydroxyl-dopamine-induced neurotoxicity in SH-SY5Y cells. Toxicol. In Vitro 2010, 24, 759–765. [Google Scholar] [CrossRef] [PubMed]
- Tian, L.L.; Wang, X.J.; Sun, Y.N.; Li, C.R.; Xing, Y.L.; Zhao, H.B.; Duan, M.; Zhou, Z.; Wang, S.Q. Salvianolic acid B, an antioxidant from Salvia miltiorrhiza, prevents 6-hydroxydopamine-induced apoptosis in SH-SY5Y cells. Int. J. Biochem. Cell Biol. 2008, 40, 409–422. [Google Scholar] [CrossRef] [PubMed]
- Blum, D.; Torch, S.; Lambeng, N.; Nissou, M.; Benabid, A.L.; Sadoul, R.; Verma, J.M. Molecular pathways involved in the neurotoxicity of 6-OHAD, dopamine and MPTP: Contribution to the apoptotic theory in Parkinson’s disease. Prog. Neurobiol. 2001, 65, 135–172. [Google Scholar] [CrossRef] [PubMed]
- Feng, L.; Meng, H.; Wu, F.; Cheng, B.; He, X.; Wang, X.; Li, Z.; Liu, S. Olfactory ensheathing cells conditioned medium prevented apoptosis induced by 6-OHDA in PC12 cells through modulation of intrinsic apoptotic pathways. Int. J. Dev. Neurosci. 2008, 26, 323–329. [Google Scholar] [CrossRef] [PubMed]
- Gorman, A.M.; Szegezdi, E.; Quigney, D.J.; Samali, A. Hsp27 inhibits 6-hydroxydopamine-induced cytochrome-c release and apoptosis in PC12 cells. Biochem. Biophys. Res. Commun. 2005, 327, 801–810. [Google Scholar] [CrossRef] [PubMed]
- Schober, A. Classic toxin-induced animal models of Parkinsonla disease: 6-OHDA and MPTP. Cell Tissue Res. 2004, 318, 215–224. [Google Scholar] [CrossRef] [PubMed]
- Mayo, J.C.; Sainz, R.M.; Antolin, I.; Rodriguez, C. Ultrastructural confirmation of neuronal protection by melatonin against the neurotoxin 6-hydroxydopamine cell damage. Brain Res. 1999, 818, 221–227. [Google Scholar] [CrossRef] [PubMed]
- Walkinshaw, G.; Waters, C.M. Neurotoxin-induced cell death in neuronal PC12 cells is mediated by induction of apoptosis. Neuroscience 1994, 63, 975–987. [Google Scholar] [CrossRef] [PubMed]
- Li, X.M. Protective effect of Lycium barbarum polysaccharides on streptozotocin-induced oxidative stress in rats. Int. J. Biol. Macromol. 2007, 40, 461–465. [Google Scholar] [CrossRef] [PubMed]
- Gan, L.; Zhang, S.H.; Yang, X.L.; Xu, H.B. Immunomodulation and antitumor activity by a polysaccharide-protein complex from Lycium barbarum. Int. Immunopharmacol. 2004, 4, 563–569. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.X.; Ng, T.B. Natural products with hypoglycemic, hypotensive, hypocholesterolemic, antiatherosclerotic and antithrombotic activities. Life Sci. 1999, 65, 2663–2677. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.B.; Li, Y.X.; Hao, Y.J.; Wang, T.F.; Lei, Z.; Wu, Y.; Zhao, Q.P.; Ang, H.; Ma, L.; Liu, J.; et al. Neuroprotective effects of LBP on brain ischemic reperfusion neurodegeneration. Eur. Rev. Med. Pharmacol. Sci. 2013, 17, 2760–2765. [Google Scholar] [PubMed]
- Yang, Y.; Li, W.; Li, Y.; Wang, Q.; Gao, L.; Zhao, J.J. Dietary Lycium barbarum Polysaccharide Induces Nrf2/ARE Pathway and Ameliorates Insulin Resistance Induced by High-Fat via Activation of PI3K/AKT Signaling. Oxid. Med. Cell. Longev. 2014, 2014, 145641. [Google Scholar] [PubMed]
- Soto-Otero, R.; Mendez-Alvarez, E.; Hermida-Ameijeiras, A.; Munoz-Patino, A.M.; Labandeira-Garcia, J.L. Autoxidation and neurotoxicity of 6-hydroxydopamine in the presence of some antioxidants: Potential implication in relation to the pathogenesis of Parkinsonki disease. J. Neurochem. 2000, 74, 1605–1612. [Google Scholar] [CrossRef] [PubMed]
- Saito, Y.; Nishio, K.; Ogawa, Y.; Kinumi, T.; Yoshida, Y.; Masuo, Y.; Niki, E. Molecular mechanisms of 6-hydroxydopamine-induced cytotoxicity in PC12 cells: Involvement of hydrogen peroxide-dependent and -independent action. Free Radic. Biol. Med. 2007, 42, 675–685. [Google Scholar] [CrossRef] [PubMed]
- Fujita, H.; Ogino, T.; Kobuchi, H.; Fujiwara, T.; Yano, H.; Akiyama, J.; Utsumi, K.; Sasaki, J. Cell-permeable cAMP analog suppresses 6-hydroxydopamine-induced apoptosis in PC12 cells through the activation of the Akt pathway. Brain Res. 2006, 1113, 10–23. [Google Scholar] [CrossRef] [PubMed]
- Guix, F.X.; Uribesalgo, I.; Coma, M.; Muomasa, F.J. The physiology and pathophysiology of nitric oxide in the brain. Prog. Neurobiol. 2005, 76, 126–152. [Google Scholar] [CrossRef] [PubMed]
- Calabrese, V.; Cornelius, C.; Rizzarelli, E.; Owen, J.B.; Dinkova-Kostova, A.T.; Butterfield, D.A. Nitric oxide in cell survival: A Janus molecule. Antioxid. Redox Signal 2009, 11, 2717–2739. [Google Scholar] [CrossRef] [PubMed]
- Del-Bel, E.; Padovan-Neto, F.E.; Raisman-Vozari, R.; Lazzarini, M. Role of nitric oxide in motor control: Implications for Parkinson’s disease pathophysiology and treatment. Curr. Pharm. Des. 2011, 17, 471–488. [Google Scholar] [CrossRef] [PubMed]
- Brown, G.C. Nitric oxide and mitochondrial respiration. Biochim. Biophys. Acta 1999, 1411, 351–369. [Google Scholar] [CrossRef] [PubMed]
- Li, X.F.; Ye, X.L.; Li, X.B.; Sun, X.; Liang, Q.; Tao, L.Z.; Kang, X.G.; Chen, J.Z. Salidroside protects against MPP+-induced apoptosis in PC12 cells by inhibiting the NO pathway. Brain Res. 2011, 1382, 9–18. [Google Scholar] [CrossRef] [PubMed]
- Barker, J.E.; Heales, S.J.; Cassidy, A.; Bolanos, J.P.; Land, J.M.; Clark, J.B. Depletion of brain glutathione results in a decrease of glutathione reductase activity: An enzyme susceptible to oxidative damage. Brain Res. 1996, 716, 118–122. [Google Scholar] [CrossRef] [PubMed]
- Naoi, M.; Maruyama, W. Future of neuroprotection in Parkinson’s disease. Parkinsonism Relat. Disord. 2001, 8, 139–145. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Dawson, V.L.; Dawson, T.M. Role of nitric oxide in Parkinson’s disease. Pharmacol. Ther. 2006, 109, 33–41. [Google Scholar] [CrossRef] [PubMed]
- Kavya, R.; Saluja, R.; Singh, S.; Dikshit, M. Nitric oxide synthase regulation and diversity: Implications in Parkinson’s disease. Indian Acad. Sci. 2006, 15, 280–294. [Google Scholar]
- Dehmer, T.; Lindenau, J.; Haid, S.; Dichgans, J.; Schulz, J.B. Deficiency of inducible nitric oxide synthase protects against MPTP toxicity in vivo. J. Neurochem. 2000, 74, 2213–2216. [Google Scholar] [CrossRef] [PubMed]
- Singh, S.; Das, T.; Ravindran, A.; Chaturvedi, R.K.; Shukla, Y.; Agarwal, A.K.; Dikshit, M. Involvement of nitric oxide in neurodegeneration: A study on the experimental models of Parkinson’s disease. Redox Rep. 2005, 10, 103–109. [Google Scholar] [CrossRef] [PubMed]
- Watanabe, H.; Muramatsu, Y.; Kurosaki, R.; Michimata, M.; Matsubara, M.; Imai, Y.; Araki, T. Protective effects of neuronal nitric oxide synthase inhibitor in mouse brain against MPTP neurotoxicity: an immunohistological study. Eur. Neuropsychopharmacol. 2004, 14, 93–104. [Google Scholar] [CrossRef] [PubMed]
- Atton, W.G.; Chalmers-Redman, R.M.E.; Ju, W.J.H.; Mammen, M.; Carlile, G.W.; Pong, A.W.; Tatton, N.A. Propargylamines induce antiapoptotic new protein synthesis in serum- and nerve growth factor (NGF)-withdrawn, NGF-differentiated PC-12 Cells. J. Pharmacol. Exp. Ther. 2002, 301, 753–764. [Google Scholar] [CrossRef] [PubMed]
- Denizot, F.; Lang, R. Rapid colorimetric assay for cell growth and survival. Modifications to the tetrazolium dye procedure giving improved sensitivity and reliability. J. Immunol. Meth. 1986, 89, 271–277. [Google Scholar] [CrossRef]
- Lee, K.E.; Kim, E.Y.; Kim, C.S.; Choi, J.S.; Bae, E.H.; Ma, S.K.; Park, J.S.; Jung, Y.D.; Kim, S.H.; Lee, J.U.; et al. Macrophage-stimulating protein attenuates hydrogen peroxide-induced apoptosis in human renal HK-2 cells. Eur. J. Pharmacol. 2013, 715, 304–311. [Google Scholar] [CrossRef] [PubMed]
- Ormerod, M.G.; Collins, M.K.L.; Rodriguez-Tarduchy, G.; Robertson, D. Apoptosis in interleukin-3-dependent haemopoietic cells: Quantification by two flow cytometric methods. J. Immunol. Meth. 1992, 153, 57–65. [Google Scholar] [CrossRef]
- Myhre, O.; Andersen, J.M.; Aarnes, H.; Fonnum, F. Evaluation of the probes 2',7'-dichlorofluorescin diacetate, luminol, and lucigenin as indicators of reactive species formation. Biochem. Pharmacol. 2003, 65, 1575–1582. [Google Scholar] [CrossRef] [PubMed]
- Kojima, H.; Urano, Y.; Kikuchi, K.; Higuchi, T.; Hirata, Y.; Nagano, T. Fluorescent Indicators for Imaging Nitric Oxide Production. Angew. Chem. Int. Ed. 1999, 38, 3209–3212. [Google Scholar] [CrossRef]
- Tao, L.Z.; Li, X.F.; Zhang, L.L.; Tian, J.Y.; Li, X.B.; Sun, X.; Li, X.F.; Jiang, L.; Zhang, X.J.; Chen, J.Z. Protective Effect of Tetrahydroxystilbene Glucoside on 6-OHDA-Induced Apoptosis in PC12 Cells through the ROS-NO Pathway. PLoS One 2011, 6, e26055. [Google Scholar] [CrossRef] [PubMed]
- Bradford, M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef] [PubMed]
- Sample Availability: Samples of LBP are available from the authors.
© 2014 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license ( http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gao, K.; Liu, M.; Cao, J.; Yao, M.; Lu, Y.; Li, J.; Zhu, X.; Yang, Z.; Wen, A. Protective Effects of Lycium barbarum Polysaccharide on 6-OHDA-Induced Apoptosis in PC12 Cells through the ROS-NO Pathway. Molecules 2015, 20, 293-308. https://doi.org/10.3390/molecules20010293
Gao K, Liu M, Cao J, Yao M, Lu Y, Li J, Zhu X, Yang Z, Wen A. Protective Effects of Lycium barbarum Polysaccharide on 6-OHDA-Induced Apoptosis in PC12 Cells through the ROS-NO Pathway. Molecules. 2015; 20(1):293-308. https://doi.org/10.3390/molecules20010293
Chicago/Turabian StyleGao, Kai, Meiyou Liu, Jinyi Cao, Minna Yao, Yunyang Lu, Jiankang Li, Xiaohe Zhu, Zhifu Yang, and Aidong Wen. 2015. "Protective Effects of Lycium barbarum Polysaccharide on 6-OHDA-Induced Apoptosis in PC12 Cells through the ROS-NO Pathway" Molecules 20, no. 1: 293-308. https://doi.org/10.3390/molecules20010293