Antioxidant and Antiproliferative Activities of Methanolic Extract from a Neglected Agricultural Product: Corn Cobs




Abstract
:1. Introduction
2. Results and Discussion
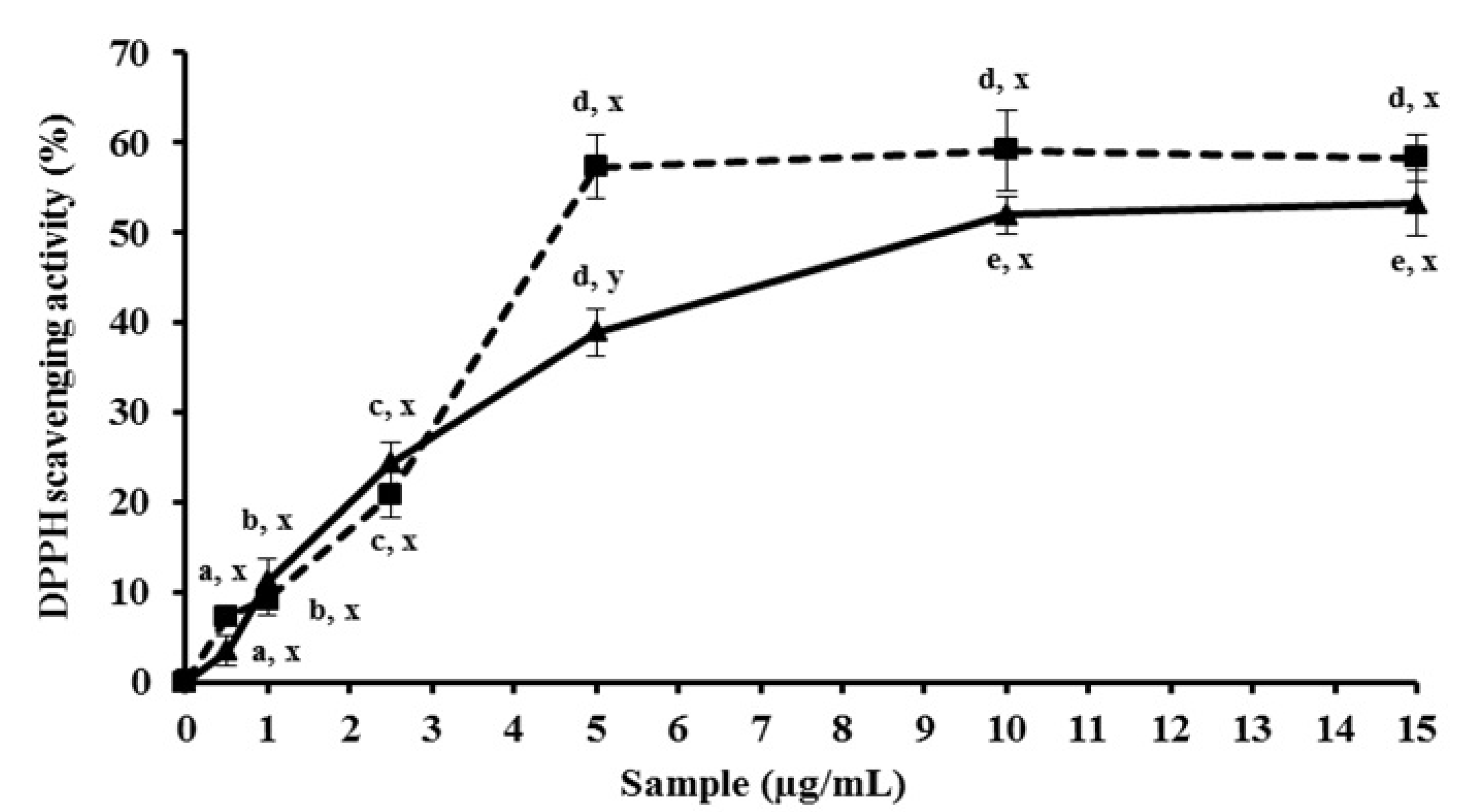
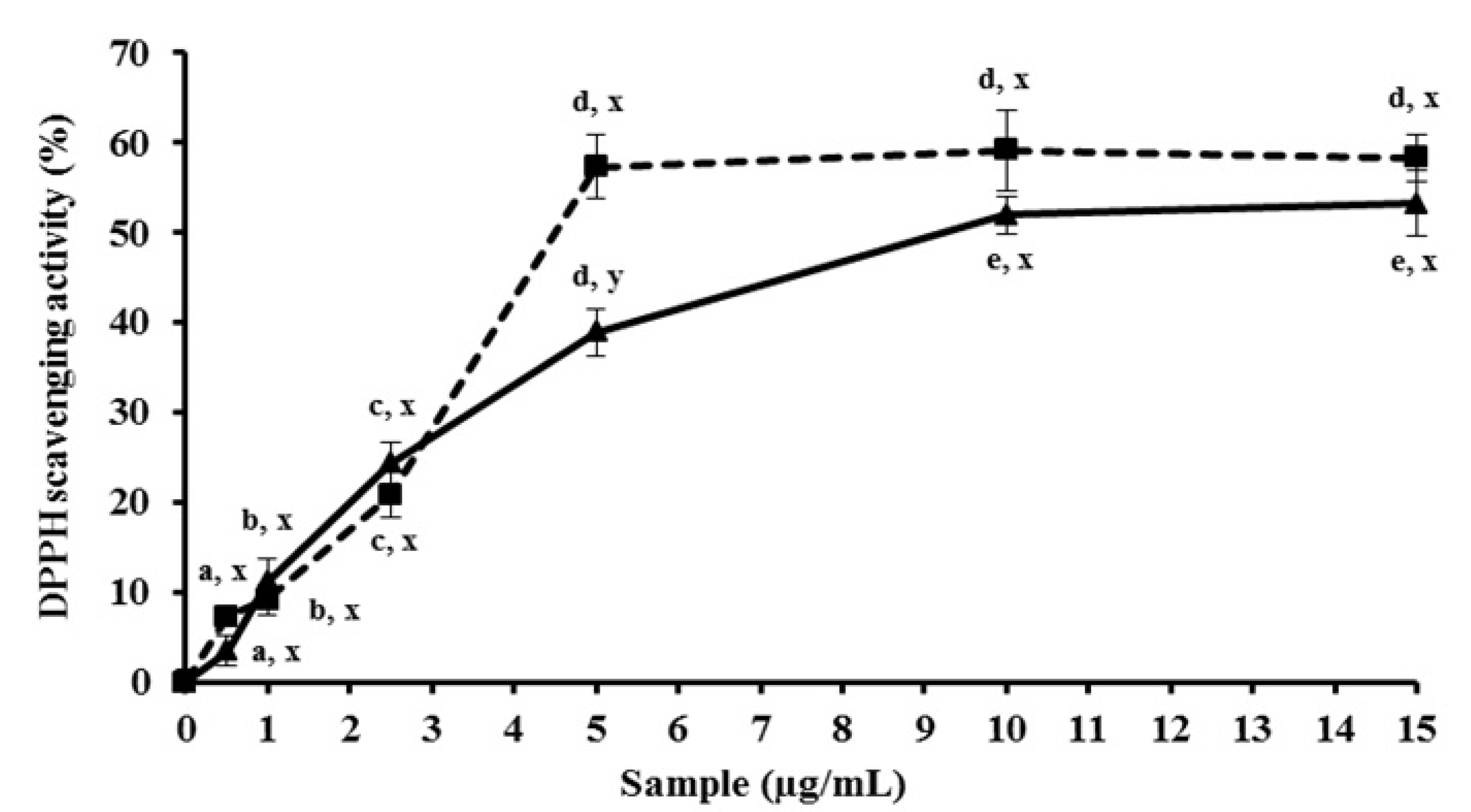
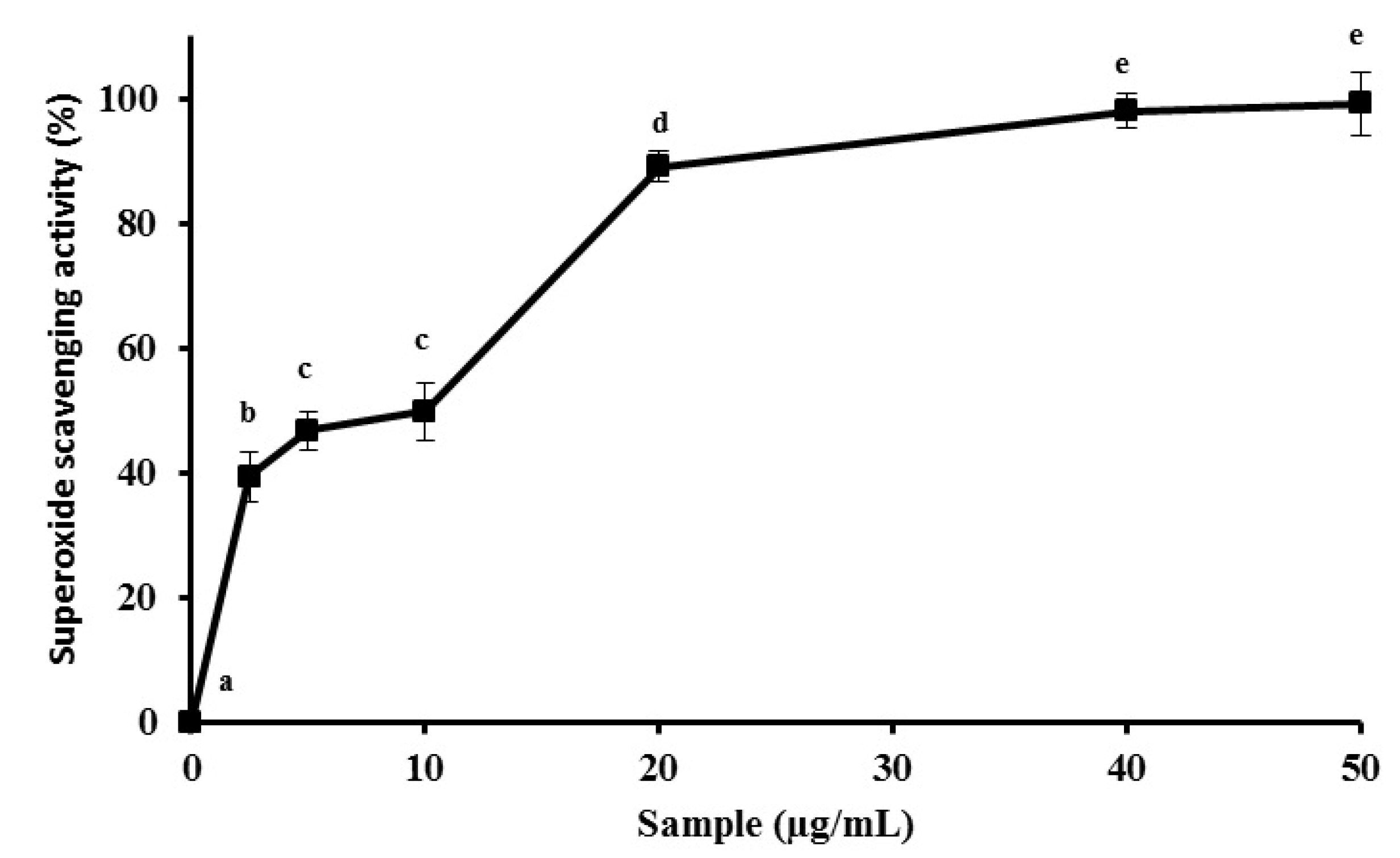
2.1. MEC up-Regulates Antioxidant Activity in Vitro


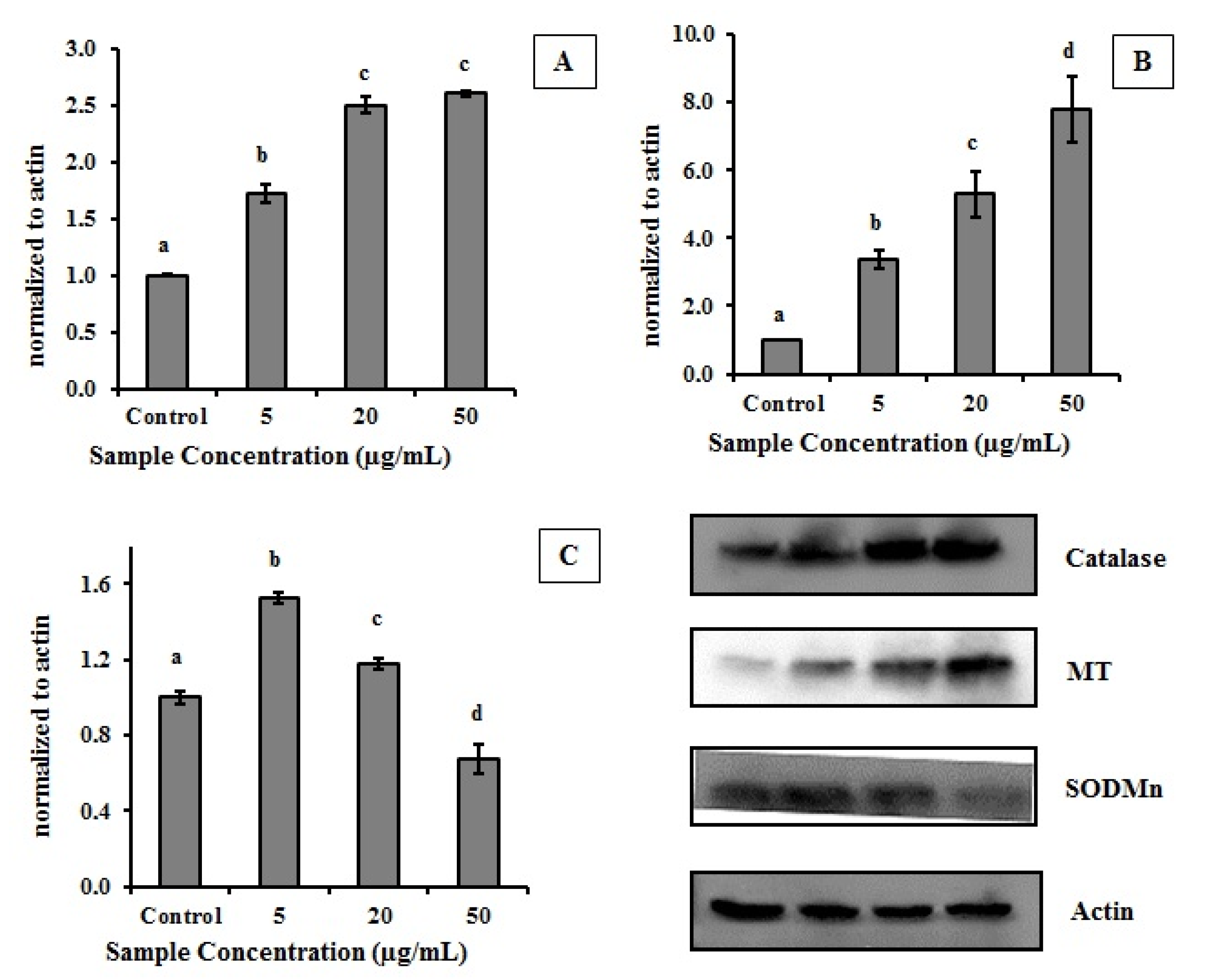
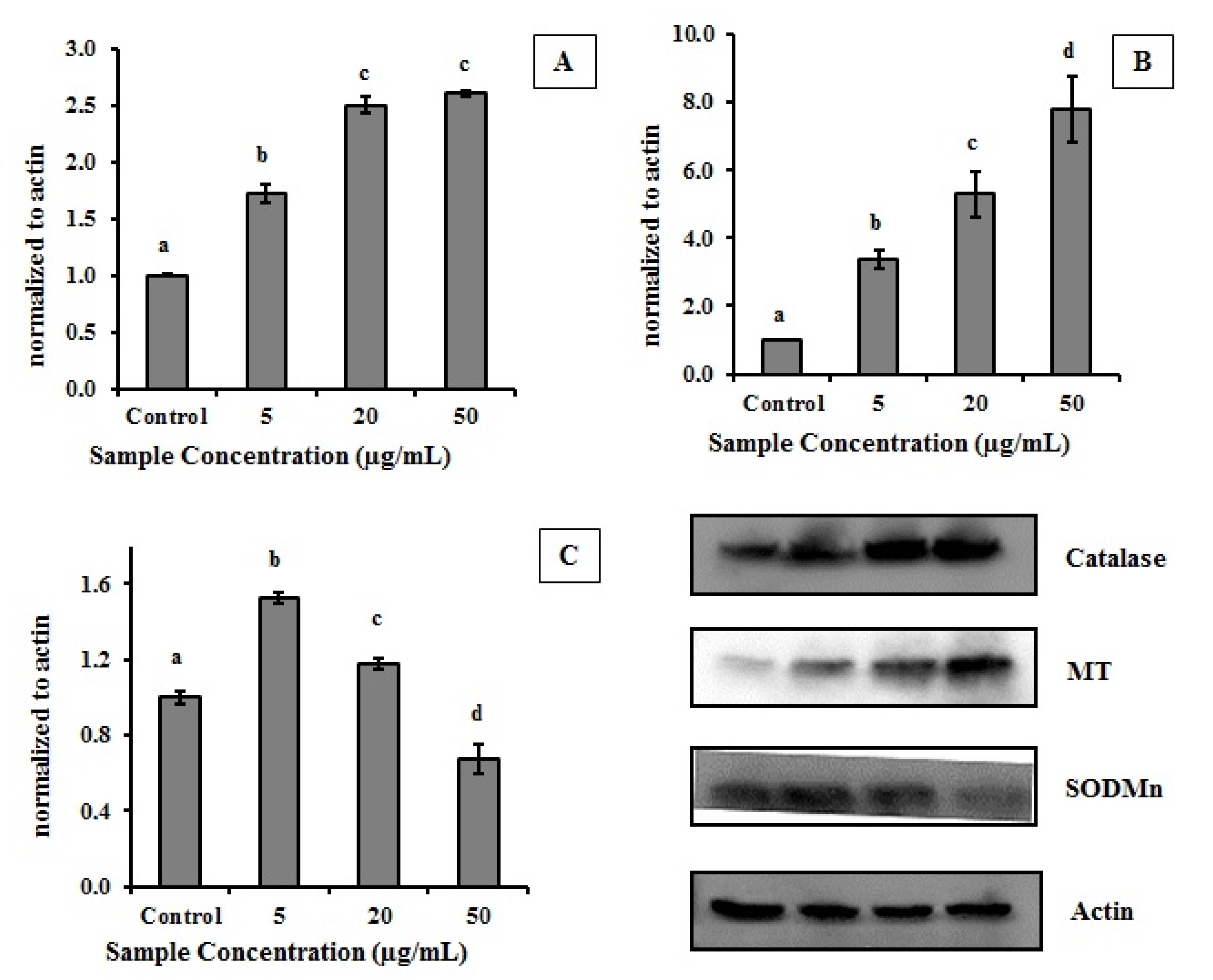
2.2. Effect of MEC on Cells Antioxidant Enzymes
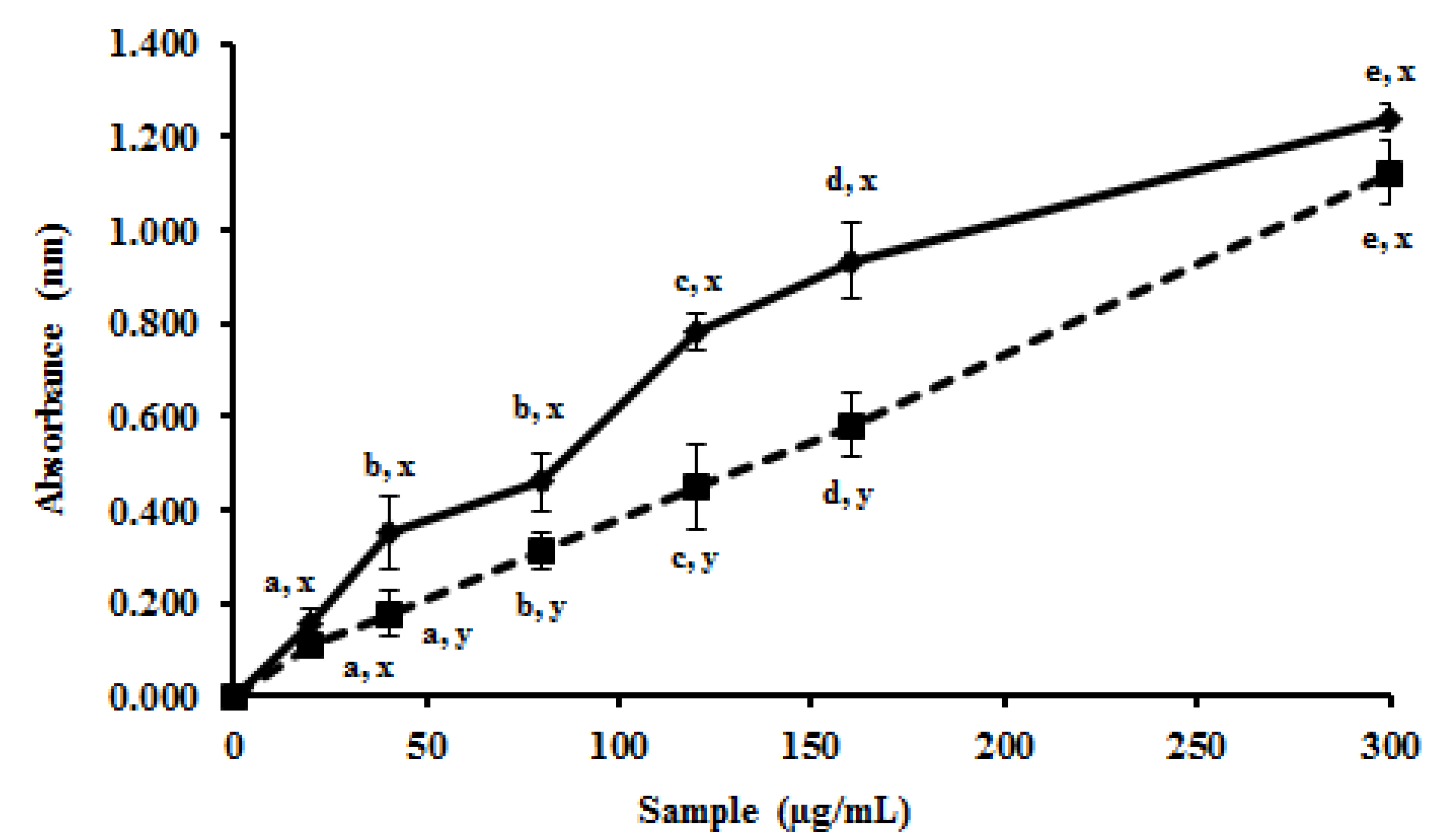
2.3. In Vivo Antioxidant Potential of MEC

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Group | Liver † | |||
|---|---|---|---|---|
| TEAC | MDA | CAT | SOD | |
| Control | 1.00 ± 0.04 | 1.00 ± 0.04 | 1.00 ± 0.18 | 1.00 ± 0.16 |
| CCl4 | a 0.65 ± 0.03 | a 2.85 ± 0.15 | a 0.50 ± 0.53 | a 0.70 ± 0.68 |
| CCl4 + Vit E | b 1.00 ± 0.05 | b 0.98 ± 0.07 | b 1.00 ± 0.18 | b 0.96 ± 0.13 |
| CCl4 + MEC | b 0.96 ± 0.04 | b 1.00 ± 0.07 | b 1.05 ± 0.13 | b 0.94 ± 0.93 |
| Group | Serum † | |||
| TEAC | MDA | CAT | SOD | |
| Control | 1.00 ± 0.47 | 1.00 ± 0.7 | 1.00 ± 0.16 | 1.00 ± 0.84 |
| CCl4 | a 0.60 ± 0.35 | a 2.26 ± 1.3 | a 0.78 ± 0.34 | a 0.60 ± 0.85 |
| CCl4 + Vit E | a 0.87 ± 0.55 | a,b 1.23 ± 0.9 | b 0.96 ± 0.39 | b 0.97 ± 0.76 |
| CCl4 + MEC | b 1.04 ± 0.36 | a,b 1.46 ± 0.7 | b 0.95 ± 0.40 | b 0.94 ± 0.11 |
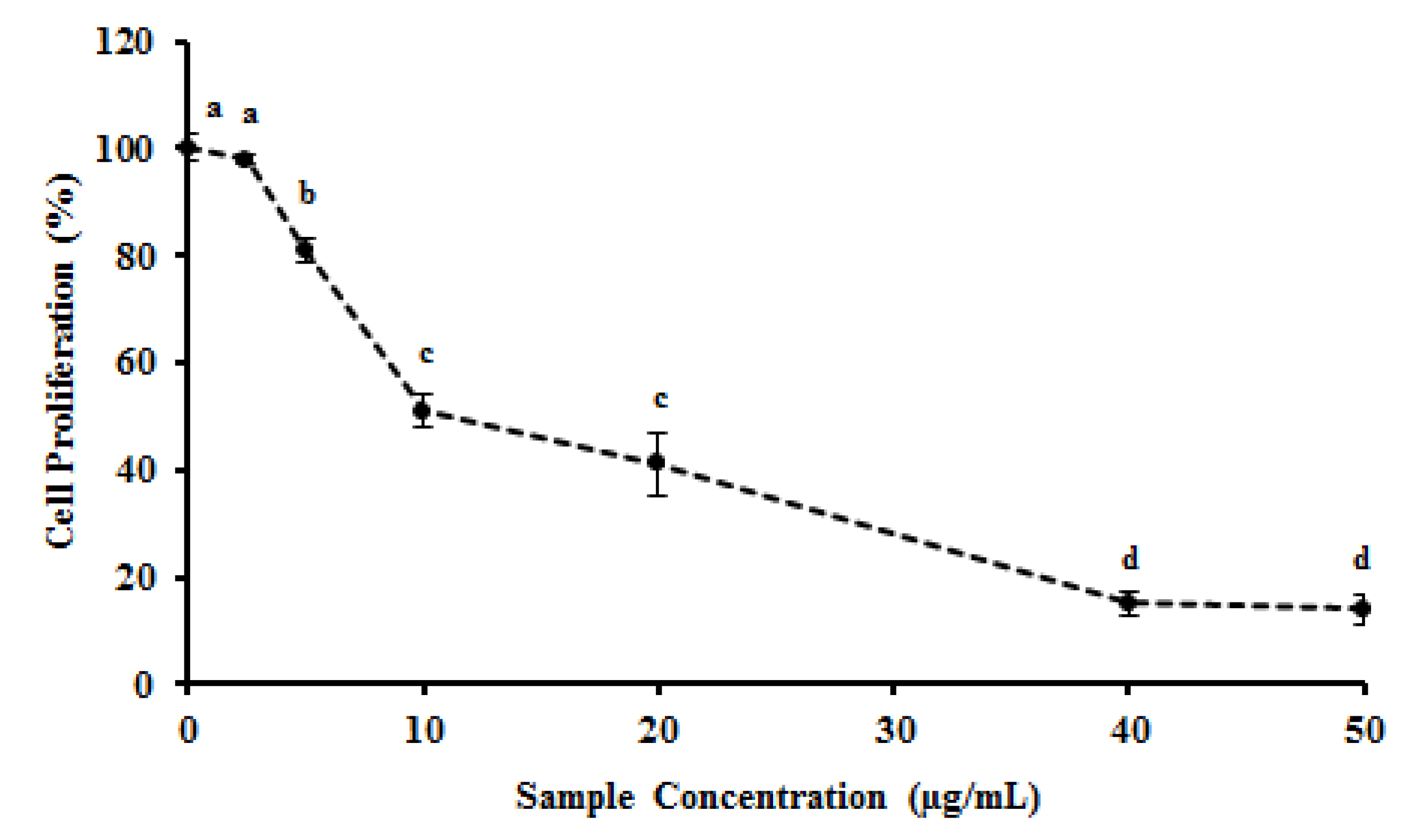
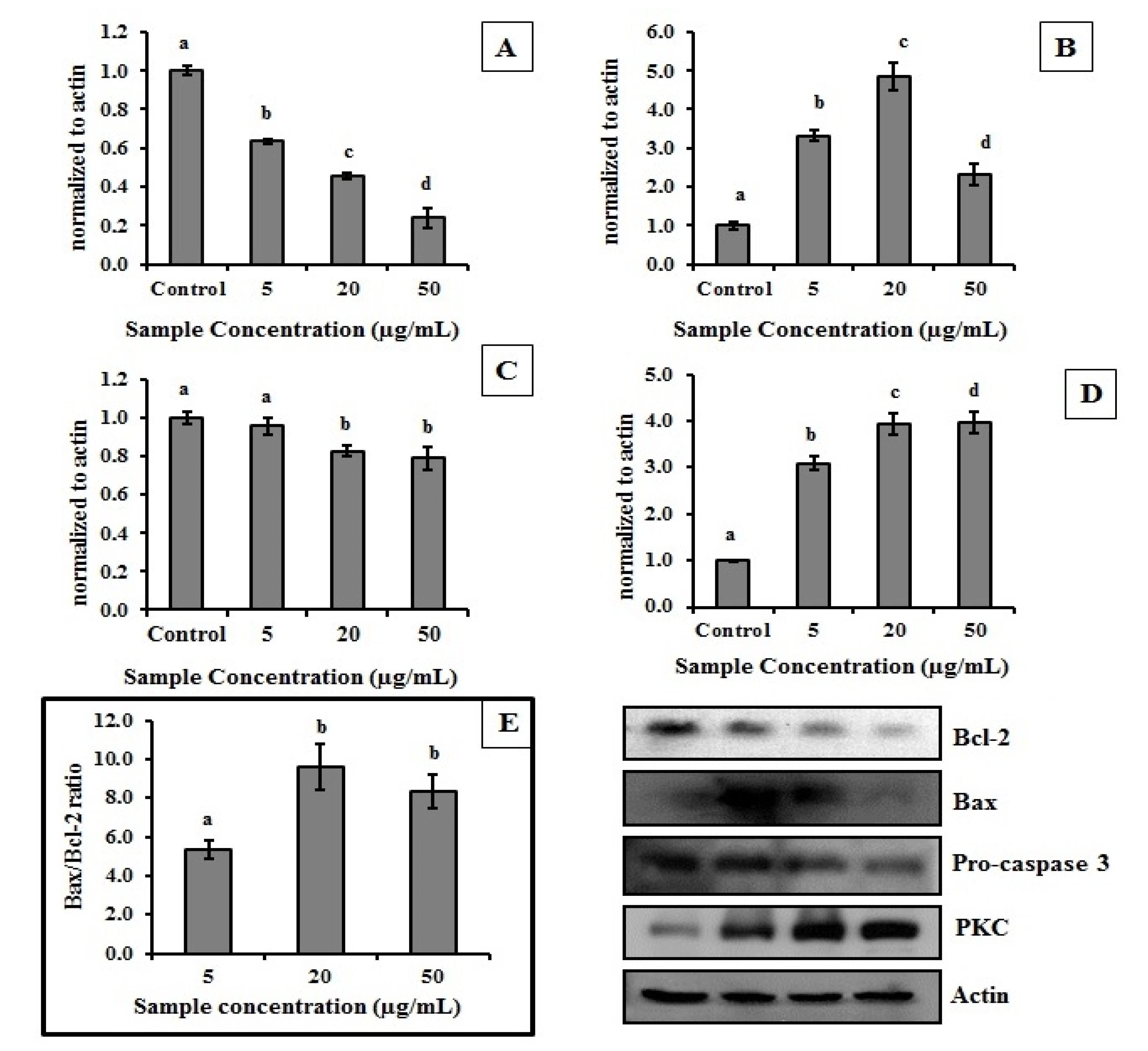
2.4. MEC Exhibits Anti-Proliferative and Pro-Apoptotic Activity
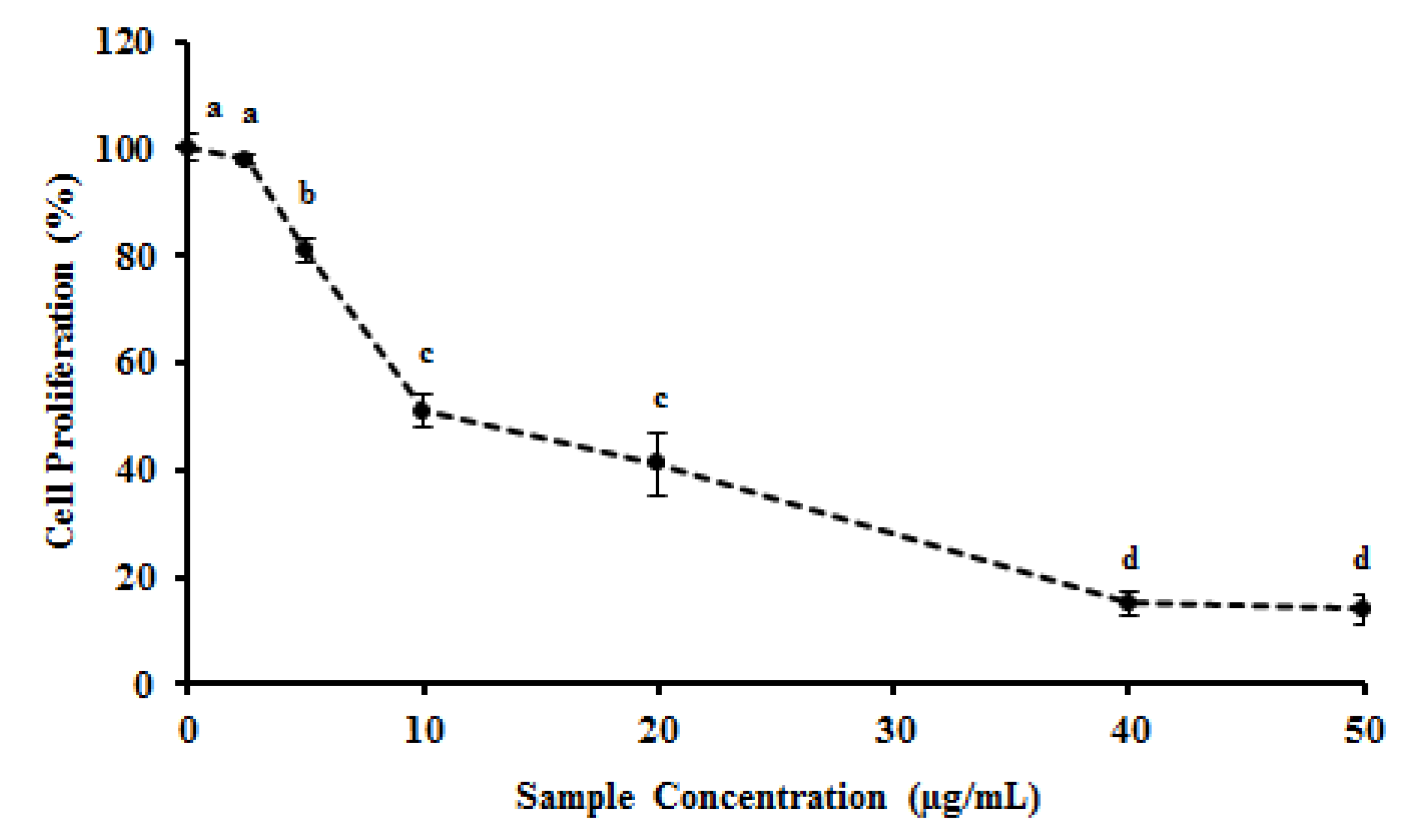
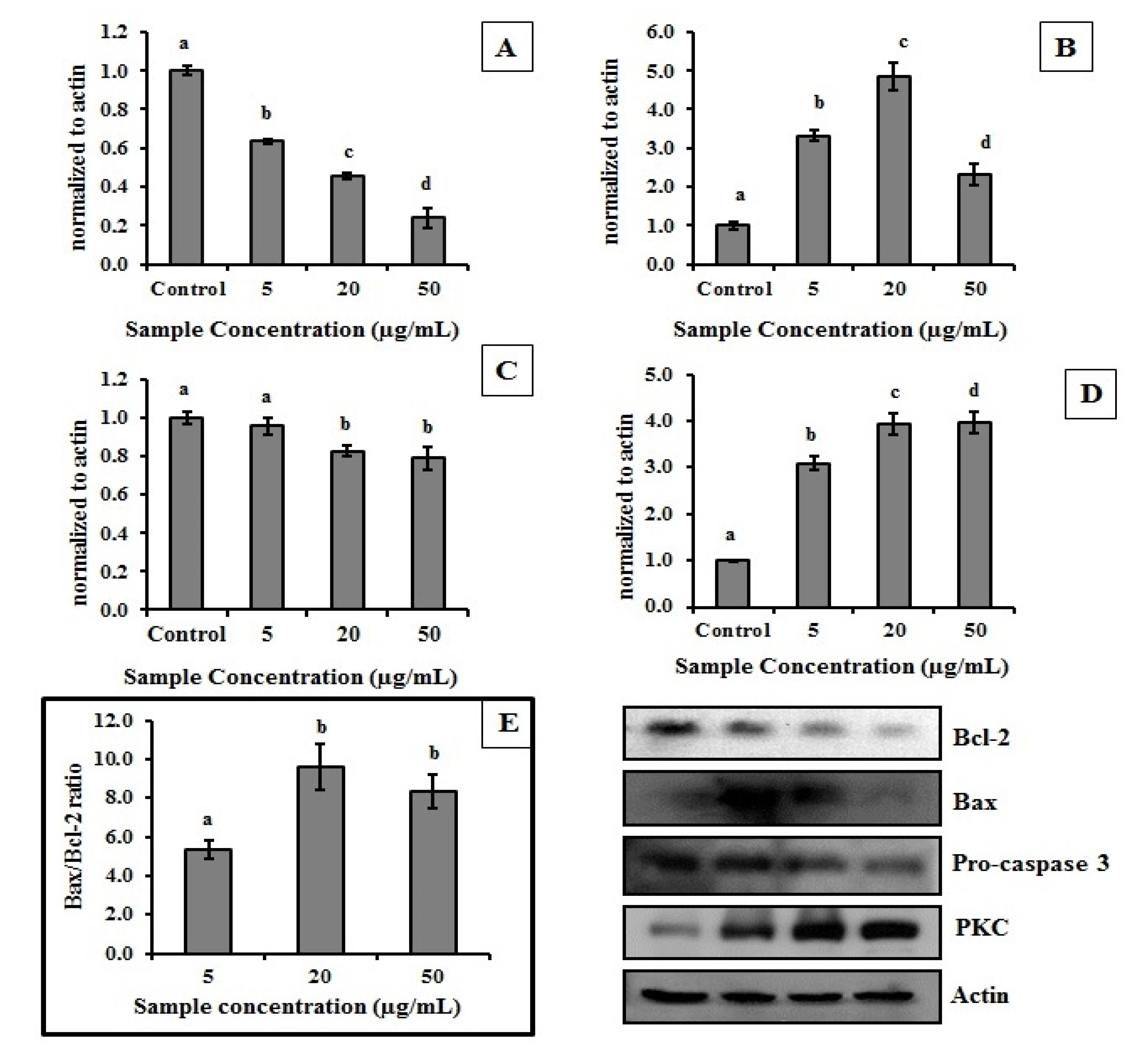
3. Experimental
3.1. Materials
3.2. Production of Methanolic Extract from Corn Cob (MEC)
3.3. In Vitro Antioxidant Tests
3.3.1. Total Antioxidant Capacity (TAC)
3.3.2. The Hydroxyl Radical Scavenging Activity
3.3.3. The Superoxide Radical Scavenging Activity
3.3.4. Ferric Chelating
3.3.5. Reducing Power
3.3.6. DPPH Assay
3.4. In Vivo Antioxidant Tests
3.4.1. Determination of Total Antioxidant Capacity of Serum and Liver Homogenate
3.4.2. Malonaldehyde (MDA) Levels
3.4.3. Catalase Activity
3.4.4. Superoxide Dismutase (SOD) Activity
3.5. MTT Assay
 × 100
× 100
3.6. Western Blot (Immunoblot)
3.7. Statistical Analysis
4. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Nagmoti, D.M.; Khatri, D.K.; Juvekar, P.R.; Juvekar, A.R. Antioxidant activity free radical-scavenging potential of Pithecellobium dulce Benth seed extracts. Free Rad. Antioxid. 2012, 2, 37–43. [Google Scholar]
- Aktumsek, A.; Zengin, G.; Guler, G.O.; Cakmak, Y.S.; Duran, A. Antioxidant potentials and anticholinesterase activities of methanolic and aqueous extracts of three endemic Centaurea L. species. Food Chem. Toxicol. 2013, 55, 290–296. [Google Scholar]
- Dröge, W. Free radicals in the physiological control of cell function. Physiol. Rev. 2002, 82, 47–95. [Google Scholar]
- Hazra, B.; Biswas, S.; Mandal, N. Antioxidant and free radical scavenging activity of Spondias pinnata. BMC Complement. Altern. Med. 2008, 8, 63:1–63:10. [Google Scholar]
- Islam, S.; Nasrin, S.; Khan, M.A.; Hossain, A.S.; Islam, F.; Khandokhar, P.; Mollah, M.N.; Rashid, M.; Sadik, G.; Rahman, M.A.; et al. Evaluation of antioxidant and anticancer properties of the seed extracts of Syzygium fruticosum Roxb. growing in Rajshahi, Bangladesh. BMC Complement. Altern. Med. 2013, 13, 142:1–142:10. [Google Scholar]
- Wang, J.; Zhang, Q.; Zhang, Z.; Li, Z. Antioxidant activity of sulfated polysaccharide fractions extracted from Laminaria japonica. Int. J. Biol. Macromol. 2008, 42, 127–132. [Google Scholar] [CrossRef]
- Barlow, S.; Schlatter, J. Risk assessment of carcinogens in food. Toxicol. Appl. Pharmacol. 2010, 243, 180–190. [Google Scholar] [CrossRef]
- Zengin, G.; Aktumsek, A.; Guler, G.O.; Cakmak, Y.S.; Yildiztugay, E. Antioxidant properties of methanolic extract and fatty acid composition of Centaurea urvillei DC. subsp. hayekiana Wagenitz. Rec. Nat. Prod. 2011, 5, 123–132. [Google Scholar]
- Costa, L.S.; Fidelis, G.P.; Telles, C.B.S.; Dantas-Santos, N.; Camara, R.B.G.; Cordeiro, S.L.; Costa, M.S.S.P.; Almeida-Lima, J.; Melo-Silveira, R.F.; Oliveira, R.M.; et al. Antioxidant and Antiproliferative Activities of Heterofucans from the Seaweed Sargassum filipendula. Mar. Drugs 2011, 9, 952–966. [Google Scholar] [CrossRef]
- Hazra, B.; Sarkar, R.; Biswas, S.; Mandal, N. Comparative study of the antioxidant and reactive oxygen species scavenging properties in the extracts of the fruits of Terminalia chebula, Terminalia belerica and Emblica officinalis. BMC Complement. Altern. Med. 2010, 10, 20:1–20:15. [Google Scholar]
- Gul, M.Z.; Bhakshu, L.M.; Ahmad, F.; Kondapi, A.K.; Qureshi, I.A.; Ghazi, I.A. Evaluation of Abelmoschus moschatus extracts for antioxidant, free radical scavenging, antimicrobial and antiproliferative activities using in vitro assays. BMC Complement. Altern. Med. 2011, 11, 64:1–64:12. [Google Scholar]
- Meyers, K.J.; Watkins, C.B.; Pritts, M.P.; Liu, R.H. Antioxidant and antiproliferative activities of strawberries. J. Agric. Food Chem. 2003, 51, 6887–6892. [Google Scholar] [CrossRef]
- Olsson, M.E.; Andersson, C.S.; Oredsson, S.; Berglund, R.H.; Gustavsson, K.E. Antioxidant levels and inhibition of cancer cell proliferation in vitro by extracts from organically and conventionally cultivated strawberries. J. Agric. Food Chem. 2006, 54, 1248–1255. [Google Scholar]
- Ema, A.; Kumar, M.S.; Rebecca, L.J.; Sindhu, S.; Anbarasi, P.; Sagadevan, E.; Arumugam, P. Evaluation of Antiproliferative effect of Grewia hirsuta on HepG2 cell lines. J. Acad. Ind. Res. 2013, 2, 1:1–1:5. [Google Scholar]
- Tutino, V.; Caruso, M.G.; Messa, C.; Perri, E.; Notarnicola, M. Antiproliferative, antioxidant and anti-inflammatory effects of hydroxytyrosol on human hepatoma HepG2 and Hep3B cell lines. Anticancer Res. 2012, 32, 5371–5377. [Google Scholar]
- Yong, Y.K.; Tan, J.J.; The, S.S.; Mah, S.H.; Lian Ee, G.C.; Chiong, H.S.; Ahmad, Z. Clinacanthus nutans extracts are antioxidant with antiproliferative effect on cultured human cancer cell lines. Evid. Based Complement. Alternat. Med. 2013, 2013, 462751:1–462751:8. [Google Scholar]
- Islam, S.; Shahid, M.; Mohammad, F. Perspectives for natural product based agents derived from industrial plants in textile applications—A review. J. Clean. Prod. 2013, 57, 2–8. [Google Scholar] [CrossRef]
- Tang, X.; Ren, N.; Xu, J. Evaluation of hydrogen production from corn cob with the mesophilic bacterium Clostridium hydrogeniproducens HR-1. Int. J. Hydrog. Energy 2013, 38, 9104–9110. [Google Scholar] [CrossRef]
- Ebringerová, A.; Hromádková, Z. The effect of ultrasound on the structure and properties of the watersoluble corn hull heteroxylan. Ultrason Sonochem. 1997, 4, 305–309. [Google Scholar] [CrossRef]
- Ebringerová, A.; Hromádková, Z.; Hřibalová, V. Structure and mitogenic activities of corn cob heteroxylans. Int. J. Biol. Macromol. 1995, 17, 327–331. [Google Scholar] [CrossRef]
- Melo-Silveira, R.F.; Fidelis, G.P.; Costa, M.S.; Telles, C.B.; Dantas-Santos, N.; de Oliveira Elias, S.; Ribeiro, V.B.; Barth, A.L.; Macedo, A.J.; Leite, E.L.; et al. In vitro antioxidant, anticoagulant and antimicrobial activity and in inhibition of cancer cell proliferation by xylan extracted from corn cobs. Int. J. Mol. Sci. 2012, 13, 409–426. [Google Scholar]
- Manzoor, M.; Anwa, F.; Mahmood, Z.; Rashid , U.; Ashraf, M. Variation in minerals, phenolics and antioxidant activity of peel and pulp of different varieties of peach (Prunus persica L.) fruit from Pakistan. Molecules 2012, 17, 6491–6506. [Google Scholar] [CrossRef]
- Pawar, C.R.; Surana, S.J. Antioxidant properties of the methanol extract of the wood and pericarp of Caesalpinia decapetala. J. Young Pharm. 2010, 2, 45–49. [Google Scholar] [CrossRef]
- Silva, J.M.C.; Dantas-Santos, N.; Gomes, D.L.; Costa, L.S.; Cordeiro, S.L.; Costa, M.S.S.P.; Silva, N.B.; Freitas, M.L.; Scortecci, K.C.; Leite, E.L.; et al. Biological activities of the sulfated polysaccharide from the vascular plant Halodule wrightii. Braz. J. Pharmacogn. 2012, 22, 94–101. [Google Scholar]
- Costa, L.S.; Fidelis, G.P.; Cordeiro, S.L.; Oliveira, R.M.; Sabry, D.A.; Camara, R.B.; Nobre, L.T.; Costa, M.S.; Almeida-Lima, J.; Farias, E.H.; et al. Biological activities of sulfated polysaccharides from tropical seaweeds. Biomed. Pharmacother. 2010, 64, 21–28. [Google Scholar] [CrossRef]
- Chaouche, T.M.; Haddouchi, F.; Ksouri, R.; Medini, F.; Atik-Bekara, F. In vitro evaluation of antioxidant activity of the hydro-methanolic extracts of Juniperus oxycedrus subsp. oxycedrus. Phytothérapie 2013, 11, 244–249. [Google Scholar]
- Li, C.; Han, W.; Wang, M.H. Antioxidant Activity of Hawthorn Fruit in vitro. J. Appl. Biol. Chem. 2010, 53, 8–12. [Google Scholar] [CrossRef]
- Sultana, B.; Anwar, F.; Przybylski, R. Antioxidant potential of corncob extracts for stabilization of corn oil subjected to microwave heating. Food Chem. 2007, 104, 997–1005. [Google Scholar] [CrossRef]
- Yu, L.; Perret, J.; Harris, M.; Wilson, J.; Haley, S. Antioxidant properties of bran extracts from “Akron” wheat grown at different locations. J. Agric. Food Chem. 2003, 51, 1566–1570. [Google Scholar] [CrossRef]
- Iqbal, S.; Bhanger, M.I.; Anwar, F. Antioxidant properties and components of bran extracts from selected wheat varieties commercially available in Pakistan. LWT Food Sci. Technol. 2007, 40, 361–367. [Google Scholar] [CrossRef]
- Lung, M.Y.; Chang, Y.C. Antioxidant Properties of the Edible Basidiomycete Armillaria mellea in Submerged Cultures. Int. J. Mol. Sci. 2011, 12, 6367–6384. [Google Scholar] [CrossRef]
- Kim, S.G.; Ahn, H.R.; Lee, K.H. Pervaporation characteristics of polyelectrolyte complex gel membranes based on two anionic polysaccharides having a chelating structure. Curr. Appl. Phys. 2009, 9, 42–46. [Google Scholar] [CrossRef]
- Manso, Y.; Carrasco, J.; Comes, G.; Adlard, P.A.; Bush, A.I.; Hidalgo, J. Characterization of the role of the antioxidant proteins metallothioneins 1 and 2 in an animal model of Alzheimer’s disease. Cell. Mol. Life Sci. 2012, 69, 3665–3681. [Google Scholar] [CrossRef]
- Serbetci, T.; Özsoy, N.; Demirci, B.; Can, A.; Kültür, S.; Baser, K.H.C. Chemical composition of the essential oil and antioxidant activity of methanolic extracts from fruits and flowers of Hypericum lydium Boiss. Ind. Crops Prod. 2012, 36, 599–606. [Google Scholar]
- Wang, J.; Yuan, X.; Jin, Z.; Tian, Y.; Song, H. Free radical and reactive oxygen species scavenging activities of peanut skins extract. Food Chem. 2007, 104, 242–250. [Google Scholar] [CrossRef]
- Khan, A.; Tania, M.; Zhang, D.Z.; Chen, H.C. Antioxidant enzymes and cancer. Chin. J. Cancer Res. 2010, 22, 87–92. [Google Scholar] [CrossRef]
- Gorrini, C.; Baniasadi, P.S.; Harris, I.S.; Silvester, J.; Inoue, S.; Snow, B.; Joshi, P.A.; Wakeham, A.; Molyneux, S.D.; Martin, B.; et al. BRCA1 interacts with Nrf2 to regulate antioxidant signaling and cell survival. J. Exp. Med. 2013, 210, 1529–1544. [Google Scholar] [CrossRef]
- Chelikani, P.; Fita, I.; Loewen, P.C. Diversity of structures and properties among catalases. Cell. Mol. Life Sci. 2004, 61, 192–208. [Google Scholar] [CrossRef]
- Peroza, E.A.; Schmucki, R.; Güntert, P.; Freisinger, E.; Zerbe, O. The beta(E)-domain of wheat E(c)-1 metallothionein: A metal-binding domain with a distinctive structure. J. Mol. Biol. 2009, 387, 207–218. [Google Scholar] [CrossRef]
- Barondeau, D.P.; Kassmann, C.J.; Bruns, C.K.; Tainer, J.A.; Getzoff, E.D. Nickel superoxide dismutase structure and mechanism. Biochemistry 2004, 43, 8038–8047. [Google Scholar]
- Liu, C.Y.; Lin, Y.C.; Deng, J.S.; Liao, J.C.; Peng, W.H.; Huang, G.J. Antioxidant, anti-inflammatory, and antiproliferative activities of Taxillus sutchuenensi. Am. J. Chin. Med. 2012, 40, 335–348. [Google Scholar] [CrossRef]
- Lin, E.S.; Chou, H.J.; Kuo, P.L.; Huang, Y.C. Antioxidant and antiproliferative activities of methanolic extracts of Perilla frutescens. J. Med. Plants Res. 2010, 4, 477–483. [Google Scholar]
- Mojžišová, G.; Mojžiš, J.; Pilátová, M.; Varinská, L.; Ivanová, L.; Strojný, L.; Richnavský, J. Antiproliferative and antiangiogenic properties of horse chestnut extract. Phytother. Res. 2012, 27, 159–165. [Google Scholar]
- Ren, G.; Zhao, Y.P.; Yang, L.; Fu, C.X. Anti-proliferative effect of clitocine from the mushroom Leucopaxillus giganteus on human cervical cancer HeLa cells by inducing apoptosis. Cancer Lett. 2008, 262, 190–200. [Google Scholar] [CrossRef]
- Ramos, P.; Santos, S.; Guerra, A.; Guerreiro, O.; Felício, L.; Jerónimo, E.; Silvestre, A.J.D.; Neto, C.P.; Duarte, M. Valorization of olive mill residues: Antioxidant and breast cancer antiproliferative activities of hydroxytyrosol-rich extracts derived from olive oil by-products. Ind. Crops Prod. 2013, 46, 359–368. [Google Scholar] [CrossRef]
- Elmore, S. Apoptosis: A review of programmed cell death. Toxicol. Pathol. 2007, 35, 495–516. [Google Scholar] [CrossRef]
- Colitti, M. BCL-2 Family of proteins and mammary cellular fate. Anat. Histol. Embryol. 2012, 41, 237–247. [Google Scholar]
- Tian, H.; Zhang, B.; Di, J.; Jiang, G.; Chen, F.; Li, H.; Li, L.; Pei, D.; Zheng, J. Keap1: One stone kills three birds Nrf2, IKKβ and Bcl-2/Bcl-xL. Cancer Lett. 2012, 325, 26–34. [Google Scholar] [CrossRef]
- Salvesen, G.S. Caspases: Opening the boxes and interpreting the arrows. Cell Death Differ. 2002, 9, 3–5. [Google Scholar] [CrossRef]
- Ghavami, S.; Hashemi, M.; Ande, S.R.; Yeganeh, B.; Xiao, W.; Eshraghi, M.; Bus, C.J.; Kadkhoda, K.; Wiechec, E.; Halayko, A.J.; et al. Apoptosis and cancer: Mutations within caspase genes. J. Med. Genet. 2009, 46, 497–510. [Google Scholar] [CrossRef]
- Telles, C.B.S.; Sabry, D.A.; Almeida-Lima, J.; Costa, M.S.S.P.; Melo-Silveira, R.F.; Trindade, E.S.; Sassaki, G.L.; Wisbeck, E.; Furlan, S.A.; Leite, E.L.; et al. Sulfation of the extracellular polysaccharide produced by the edible mushroom Pleurotus sajor-caju alters its antioxidant, anticoagulant and antiproliferative properties in vitro. Carbohydr. Polym. 2011, 85, 514–521. [Google Scholar] [CrossRef]
- Hvattum, E.; Stenstrøm, Y.; Ekeberg, D. Study of the reaction products of flavonols with 2,2-diphenyl-1-picrylhydrazyl using liquid chromatography coupled with negative electrospray ionization tandem mass spectrometry. J. Mass Spectrom. 2004, 39, 1570–1581. [Google Scholar] [CrossRef]
- Aebi, H. Catalase in vitro. Meth. Enzimol. 1984, 105, 121–126. [Google Scholar] [CrossRef]
- Beyer, W.E.; Fridovich, I. Assaying for superoxide dismutase activity: Some large consequence of minor changes in conditions. Anal. Biochem. 1987, 161, 559–556. [Google Scholar] [CrossRef]
- Yang, X.; Yang, S.; Guo, Y.; Jiao, Y.; Zhao, Y. Compositional characterisation of soluble apple polysaccharides, and their antioxidant and hepatoprotective effects on acute CCl4-caused liver damage in mice. Food Chem. 2013, 138, 1256–1264. [Google Scholar]
- Sample Availability: Samples of the compounds are available from the authors.
© 2014 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license ( http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Melo-Silveira, R.F.; Fidelis, G.P.; Viana, R.L.S.; Soeiro, V.C.; Silva, R.A.d.; Machado, D.; Costa, L.S.; Ferreira, C.V.; Oliveira Rocha, H.A. Antioxidant and Antiproliferative Activities of Methanolic Extract from a Neglected Agricultural Product: Corn Cobs. Molecules 2014, 19, 5360-5378. https://doi.org/10.3390/molecules19045360
Melo-Silveira RF, Fidelis GP, Viana RLS, Soeiro VC, Silva RAd, Machado D, Costa LS, Ferreira CV, Oliveira Rocha HA. Antioxidant and Antiproliferative Activities of Methanolic Extract from a Neglected Agricultural Product: Corn Cobs. Molecules. 2014; 19(4):5360-5378. https://doi.org/10.3390/molecules19045360
Chicago/Turabian StyleMelo-Silveira, Raniere Fagundes, Gabriel Pereira Fidelis, Rony Lucas Silva Viana, Vinícius Campelo Soeiro, Rodrigo Augusto da Silva, Daisy Machado, Leandro Silva Costa, Carmen Veríssima Ferreira, and Hugo Alexandre Oliveira Rocha. 2014. "Antioxidant and Antiproliferative Activities of Methanolic Extract from a Neglected Agricultural Product: Corn Cobs" Molecules 19, no. 4: 5360-5378. https://doi.org/10.3390/molecules19045360