Seasonal Variation of the Chemical Composition and Antimicrobial and Cytotoxic Activities of the Essential Oils from Inga laurina (Sw.) Willd.

Abstract
:1. Introduction
2. Results and Discussion
2.1. Yield and Chemical Composition of the Essential Oils

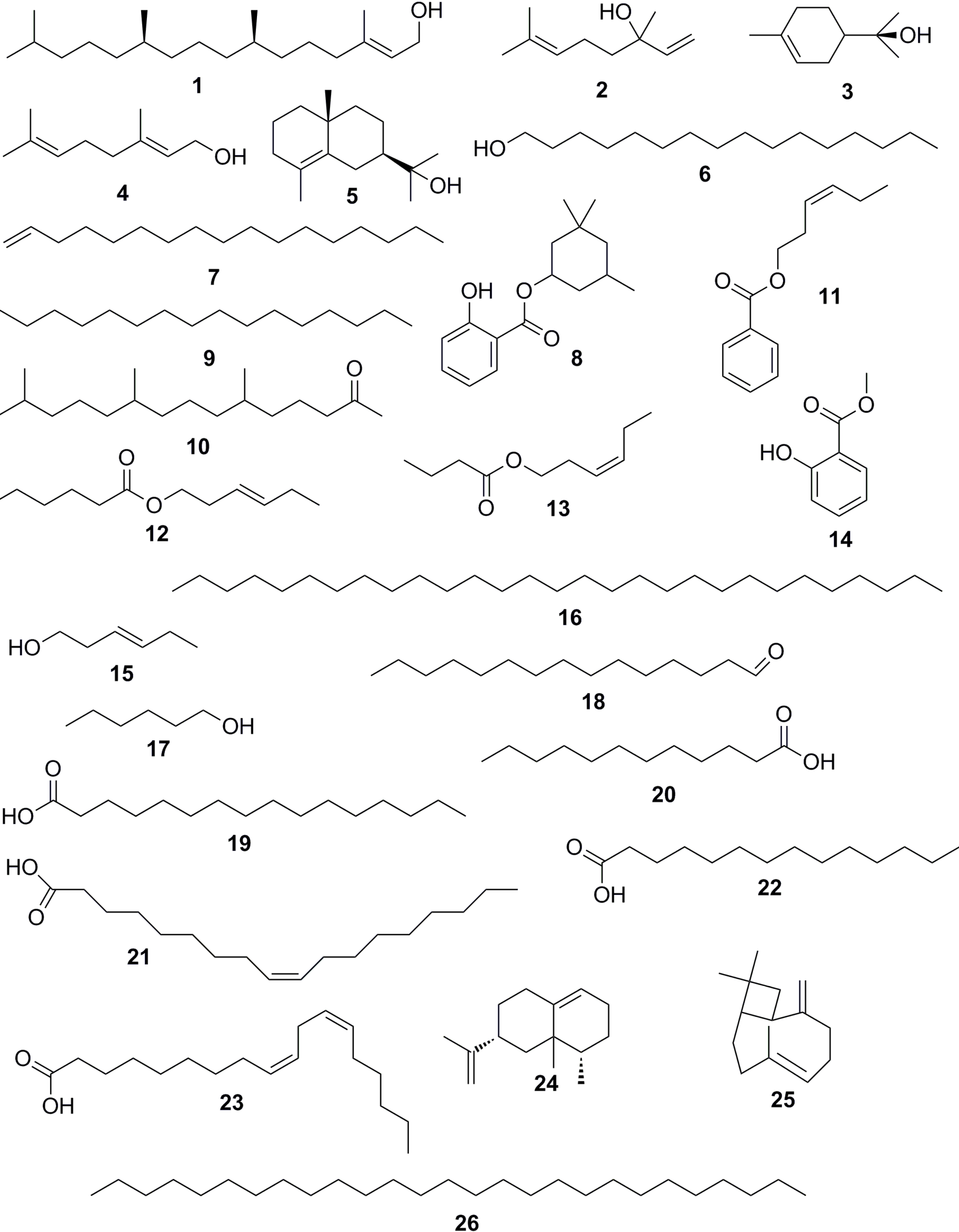{kind=link}
| Compound | AI Reference | AI Calculated | Composition % TIC | ||||||
|---|---|---|---|---|---|---|---|---|---|
| Stem bark | Leaves | ||||||||
| Ethyl butanoate | 802 b | 807 | 3.03 | - | |||||
| ( E)-Hex-3-en-1-ol | 852 a | 853 | - | 1.09 | |||||
| ( Z)-Hex-3-en-1-ol | 853 a | 856 | - | 14.23 | |||||
| Hexan-1-ol * | 863 b | 868 | - | 5.69 | |||||
| Benzyl alcohol * | 1034 a | 1041 | - | 1.46 | |||||
| Linalool * | 1100 a | 1105 | 5.17 | - | |||||
| Hotrienol | 1104 a | 1108 | - | 1.74 | |||||
| (3 E,6Z)-Nona-3,6-dien-1-ol | 1160 a | 1163 | - | 1.43 | |||||
| 3-Hexenyl butanoate | 1184 b | 1188 | - | 7.60 | |||||
| Hexyl butanoate | 1188 a | 1194 | - | 1.95 | |||||
| α-Terpineol * | 1195 a | 1199 | 4.18 | - | |||||
| Methyl salicylate * | 1196 a | 1200 | - | 4.16 | |||||
| ( Z)-Hex-3-enyl 2-methylbutanoate | 1229 b | 1235 | - | 2.06 | |||||
| Geraniol * | 1254 a | 1263 | 3.71 | - | |||||
| ( Z)-Hex-3-enyl hexanoate | 1378 b | 1383 | - | 8.29 | |||||
| Hexyl hexanoate | 1382 b | 1388 | - | 3.47 | |||||
| ( E)-Hex-2-enyl hexanoate | 1391 a | 1391 | - | 0.91 | |||||
| N.I. | - | 1436 | - | 2.19 | |||||
| N.I. | - | 1443 | 3.94 | - | |||||
| N.I. | - | 1568 | - | 1.70 | |||||
| ( Z)-Hex-3-enyl benzoate | 1572 a | 1576 | - | 10.15 | |||||
| Hexyl benzoate | 1579 b | 1582 | - | 2.03 | |||||
| ( E)-Hex-2-enyl benzoate | 1587 b | 1590 | - | 1.73 | |||||
| Hexadecane * | 1600 b | 1600 | 4.57 | - | |||||
| Oxygenated sesquiterpene | - | 1608 | 3.75 | - | |||||
| γ-Eudesmol | 1630 b | 1640 | 3.48 | - | |||||
| N.I. | - | 1670 | - | 1.32 | |||||
| N.I. | - | 1671 | 4.33 | - | |||||
| Heptadec-8-ene | 1677 a | 1678 | 7.66 | - | |||||
| Heptadecane * | 1700 b | 1700 | 2.79 | - | |||||
| N.I. | - | 1708 | 4.88 | - | |||||
| Pentadecanal | 1717 a | 1715 | - | 3.69 | |||||
| N.I. | - | 1773 | 1.85 | - | |||||
| 2-Ethylhexyl salicylate | 1807 b | 1810 | 3.25 | - | |||||
| Hexahydrofarnesyl acetone (phytone) | 1843 a | 1846 | 3.90 | 1.24 | |||||
| Hexadecanol | 1874 b | 1883 | 8.34 | - | |||||
| Heptadecadienal | - | 1889 | - | 1.78 | |||||
| 3,3,5-Trimethylcyclohexyl salicylate (Homosalate) | - | 1891 | 4.60 | - | |||||
| N.I. | - | 1895 | - | 4.29 | |||||
| Phytol * | 2114 a | 2116 | 9.76 | 2.58 | |||||
| N.I. | - | 2154 | 3.43 | - | |||||
| N.I. | - | 2203 | 3.01 | - | |||||
| N.I. | - | 2265 | 3.41 | - | |||||
| N.I. | - | 2308 | 3.22 | - | |||||
| Pentacosane * | 2500 b | 2500 | 3.16 | - | |||||
| Heptacosane * | 2700 b | 2700 | - | 2.66 | |||||
| Nonacosane * | 2900 b | 2900 | - | 9.77 | |||||
| Total (%): | 99.42 | 99.21 | |||||||
| Functional groups | Stem bark (%) | Leaves (%) |
|---|---|---|
| Alcohols | 8.34 (1) | 23.90 (5) |
| Esters | 10.88 (3) | 42.35 (10) |
| Aldehydes | - | 5.47 (2) |
| Ketones | 3.90 (1) | 1.24 (1) |
| Oxygenated monoterpenes | 13.06 (3) | 1.74 (1) |
| Oxygenated sesquiterpenes | 7.23 (2) | - |
| Oxygenated diterpenes | 9.76 (1) | 2.58 (1) |
| Long-chain alkanes | 10.52 (3) | 12.43 (2) |
| Long-chain alkenes | 7.66 (1) | - |
| N.I. | 28.07 (8) | 9.50 (4) |

| Compound | AI Reference | AI Calculated | Composition % TIC | ||||||
|---|---|---|---|---|---|---|---|---|---|
| Stem bark | Leaves | ||||||||
| 4-Methyhexan-3-one | - | 840 | 1.19 | - | |||||
| ( Z)-Hex-3-en-1-ol | 853 a | 856 | - | 9.59 | |||||
| Hexan-1-ol * | 863 b | 868 | - | 0.70 | |||||
| N.I. | - | 979 | 1.21 | - | |||||
| N.I. | - | 1001 | 1.18 | - | |||||
| Linalool * | 1100 a | 1105 | 2.69 | - | |||||
| Oxygenated monoterpenes | - | 1140 | 7.80 | - | |||||
| 3-Hexenyl butanoate | 1184 b | 1188 | - | 0.59 | |||||
| ( Z)-Hex-3-enyl hexanoate | 1378 b | 1383 | - | 0.37 | |||||
| Eremophylene | 1464 a | 1467 | 1.51 | - | |||||
| 9-epi-( E)-Caryophyllene | 1468 a | 1473 | 1.36 | - | |||||
| Sesquiterpene | - | 1491 | 1.40 | - | |||||
| ( E)-Nerolidol * | 1564 a | 1568 | - | 0.28 | |||||
| Dodecanoic acid (lauric acid) * | 1568 b | 1575 | 5.80 | 0.43 | |||||
| Tetradecanal | 1611 b | 1611 | 0.94 | - | |||||
| Oxygenated sesquiterpene | - | 1637 | 1.27 | - | |||||
| N.I. | - | 1644 | 1.29 | - | |||||
| N.I. | - | 1668 | 1.83 | - | |||||
| Heptadec-8-ene | 1677 a | 1678 | 3.48 | - | |||||
| Heptadecane * | 1700 b | 1700 | 1.68 | - | |||||
| Oxygenated sesquiterpene | - | 1706 | 1.59 | - | |||||
| Oxygenated sesquiterpene | - | 1713 | 2.01 | - | |||||
| N.I. | - | 1714 | - | 0.43 | |||||
| Oxygenated sesquiterpene | - | 1746 | 5.61 | - | |||||
| Tetradecanoic acid (myristic acid) * | 1775 a | 1771 | 4.06 | 0.32 | |||||
| Hexahydrofarnesyl acetone (phytone) | 1843 a | 1846 | 1.99 | 0.61 | |||||
| Hexadecanol | 1874 b | 1883 | 2.15 | 0.37 | |||||
| 3,3,5-Trimethylcyclohexyl salicylate (homosalate) | - | 1891 | 3.87 | - | |||||
| N.I. | - | 1919 | - | 0.99 | |||||
| N.I. | - | 1941 | 1.16 | - | |||||
| Isophytol | 1946 b | 1947 | - | 0.35 | |||||
| Hexadec-9-enoic acid (palmitoleic acid) | 1957 c | 1948 | 1.58 | - | |||||
| N.I. | - | 1962 | 1.04 | - | |||||
| Hexadecanoic acid (palmitic acid) * | 1970 a | 1972 | 25.40 | 15.20 | |||||
| N.I. | - | 2082 | - | 0.87 | |||||
| N.I. | - | 2099 | - | 0.80 | |||||
| Phytol * | 2114 a | 2116 | 1.39 | 33.21 | |||||
| (9 Z,12Z)-Octadeca-9,12-dienoic acid (linoleic acid) * | 2132 b | 2140 | 3.58 | 1.19 | |||||
| ( Z)-Octadec-9-enoic acid (oleic acid) * | 2141 b | 2144 | 5.52 | 2.33 | |||||
| N.I. | - | 2149 | - | 1.28 | |||||
| Octadecanoic acid (stearic acid) * | 2170 a | 2169 | 0.90 | 2.25 | |||||
| Tricosane * | 2300 b | 2300 | - | 0.43 | |||||
| Pentacosane * | 2500 b | 2500 | 0.89 | - | |||||
| Heptacosane * | 2700 b | 2700 | 0.69 | 3.33 | |||||
| Octacosane * | 2800 b | 2800 | - | 1.33 | |||||
| Nonacosane * | 2900 b | 2900 | - | 21.95 | |||||
| Triacontane * | 3000 b | 3000 | 0.95 | - | |||||
| Total (%) | 99.01 | 99.20 | |||||||
| Functional groups | Stem bark (%) | Leaves (%) |
|---|---|---|
| Alcohols | 2.15 (1) | 10.66 (3) |
| Esters | 3.87 (1) | 0.96 (2) |
| Aldehydes | 0.94 (1) | - |
| Ketones | 3.18 (2) | 0.61 (1) |
| Oxygenated monoterpenes | 10.49 (2) | - |
| Sesquiterpenes | 4.27 (3) | - |
| Oxygenated sesquiterpenes | 10.48 (4) | 0.28 (1) |
| Oxygenated diterpenes | 1.39 (1) | 33.56 (2) |
| Long chain alkanes | 4.21 (4) | 27.04 (4) |
| Alkenes | 3.48 (1) | - |
| Fatty acids | 46.84 (7) | 21.72 (6) |
| N.I. | 7.71 (6) | 4.37 (5) |
2.2. Antimicrobial and Cytotoxic Activities of the Essential Oils
| Sample/Season | ||||||
|---|---|---|---|---|---|---|
| Dry | Rainy | |||||
| Microorganisms | Stem bark | Leaves | Stem bark | Leaves | CHD * | |
| Anaerobic | Porphyromonas gingivalis a ATCC 33277 | 100 | 100 | 100 | 50 | 3688 |
| Prevotella nigrescens a ATCC 33563 | 200 | 100 | 400 | 100 | 1844 | |
| Fusobacterium nucleatum a ATCC 25586 | >400 | >400 | 400 | 200 | 3688 | |
| Actinomyces naeslundii b ATCC 19039 | >400 | >400 | >400 | 400 | 1844 | |
| Bacteroides fragilis a ATCC 25285 | >400 | >400 | >400 | >400 | 1475 | |
| Aerobic | Streptococcus mutans b ATCC 25175 | 200 | 200 | 25 | 50 | 0922 |
| Streptococcus sanguinis b ATCC 10556 | 200 | 100 | 50 | 50 | 3688 | |
| Streptococcus salivarius b ATCC 25975 | 200 | 100 | 25 | 25 | 0922 | |
| Streptococcus sobrinus b ATCC 33478 | 200 | 200 | 25 | 25 | 1844 | |
| Streptococcus mitis b ATCC 49456 | 100 | 100 | 50 | 50 | 3688 | |
| Cytotoxic activity | Vero cells ATCC CCL 81 | 456 ± 9 | 227 ± 16 | >512 | >512 | ----- |
3. Experimental
3.1. Plant Material and Essential Oil Extraction
3.2. Analysis and Identification of the Constituents
3.3. Microbial Strains
3.4. Antimicrobial Activity
3.5. Cytotoxic Activity
3.6. Statistical Analysis
4. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Vivot, E.; Muñoz, J.D.; Cruañes, M.C.; Cruañes, M.J.; Tapia, A.; Hirschmann, G.S.; Martínez, E.; di Sapio, O.; Gattuso, M.; Zacchino, S. Inhibitory activity of xanthine-oxidase and superoxide scavenger properties of Inga verna subsp. affinis. Its morphological and micrographic characteristics. J. Ethnopharmacol. 2001, 76, 65–71. [Google Scholar] [CrossRef]
- Silva, E.M.; Rogez, H.; Larondelle, Y. Optimization of extraction of phenolics from Inga edulis leaves using response surface methodology. Sep. Purif. Technol. 2007, 55, 381–387. [Google Scholar] [CrossRef]
- Lokvam, J.; Clausen, T.P.; Grapov, D.; Coley, P.D.; Kursar, T.A. Galloyl depsides of tyrosine from young leaves of Inga laurina. J. Nat. Prod. 2007, 70, 134–136. [Google Scholar] [CrossRef]
- Coley, P.D.; Lokvam, J.; Rudolph, K.; Bromberg, K.; Sackett, T.E.; Wright, L.; Brenes-Arguedas, T.; Dvorett, D.; Ring, S.; Clark, A.; et al. Divergent defensive strategies of young leaves in two species of Inga. Ecology 2005, 86, 2633–2643. [Google Scholar] [CrossRef]
- Morton, T.C.; Zektzer, A.S.; Rife, J.P.; Romeo, J.T. Trans-4-methoxypipecolic acid, an amino acid from Inga paterno. Phytochemistry 1991, 30, 2397–2399. [Google Scholar] [CrossRef]
- Morton, T.C. Chemotaxonomic significance of hydroxylated pipecolic acids in Central American Inga (Fabaceae: Mimosoideae: Ingeae). Biochem. Sys. Ecol. 1998, 26, 379–401. [Google Scholar] [CrossRef]
- Correa, S.M.V.; Conserva, L.M.; Maia, J.G.S. Constituents of roots of Inga edulis var. parviflora. Fitoterapia 1995, 66, 379–380. [Google Scholar]
- Dias, A.L.S.; Souza, J.N.S.; Rogez, H. Enriquecimento de compostos fenólicos de folhas de Inga edulis por extração em fase sólida: Quantificação de seus compostos majoritários e avaliação da capacidade antioxidante. Quím. Nova 2010, 33, 38–42. [Google Scholar] [CrossRef]
- Souza, J.N.S.; Silva, E.M.; Silva, M.N.; Arruda, M.S.R.; Larondelle, Y.; Rogez, H. Identification and antioxidant activity of several flavonoids of Inga edulis leaves. J. Braz. Chem. Soc. 2007, 18, 1276–1280. [Google Scholar] [CrossRef]
- Lokvam, J.; Coley, P.D.; Kursar, T.A. Cinnamoyl glucosides of catechin and dimeric procyanidins from young leaves of Inga umbellifera (Fabaceae). Phytochemistry 2004, 65, 351–358. [Google Scholar] [CrossRef]
- Kursar, T.A.; Dexter, K.G.; Lokvam, J.; Pennington, R.T.; Richardson, J.E.; Weber, M.G.; Murakami, E.T.; Drake, C.; McGregor, R.; Coley, P.D. The evolution of antiherbivore defenses and their contribution to species coexistence in the tropical tree genus Inga. Proc. Natl. Acad. Sci. USA 2009, 106, 18073–18078. [Google Scholar] [CrossRef]
- Macedo, M.L.R.; Garcia, V.A.; Freire, M.D.G.M.; Richardson, M. Characterization of a Kunitz trypsin inhibitor with a single disulfide bridge from seeds of Inga laurina (SW.) Willd. Phytochemistry 2007, 68, 1104–1111. [Google Scholar] [CrossRef]
- Rao, B.V.; Ramanjaneyulu, K.; Rambabu, T.; Devi, C.H.B.T.S. Synthesis and antioxidant activity of galloyltyrosine, derivatives from young leaves of Inga laurina. Int. J. Pharm. Biol. Sci. 2011, 2, 39–44. [Google Scholar]
- Antoun, M.D.; Ramos, Z.; Vasquez, J.; Oquendo, I.; Proctor, G.R.; Gerena, L.; Franzblau, S.G. Evaluation of the flora of Puerto Rico for in vitro antiplasmodial and antimycobacterial activities. Phytother. Res. 2001, 15, 638–642. [Google Scholar]
- Oliva, M.L.V. A novel subclassification for Kunitz protease inhibitors from leguminous seeds. Biochimie 2010, 92, 1667–1673. [Google Scholar] [CrossRef]
- Ramos, V.S.; Cabrera, O.G.; Camargo, E.L.; Ambrósio, A.B.; Vidal, R.O.; Silva, D.S.; Guimarães, L.C.; Marangoni, S.; Parra, J.R.; Pereira, G.A.; et al. Molecular cloning and insecticidal effect of Inga laurina trypsin inhibitor on Diatraea saccharalis and Heliothis virescens. Comp. Biochem. Physiol. C Toxicol. Pharmacol. 2012, 156, 148–158. [Google Scholar] [CrossRef]
- Macedo, M.L.R.; Freire, M.G.M.; Franco, O.L.; Migliolo, L.; Oliveira, C.F.R. Practical and theoretical characterization of Inga laurina Kunitz inhibitor on the control of Homalinotus coriaceus. Comp. Biochem. Physiol. B Biochem. Mol. Biol. 2011, 158, 164–172. [Google Scholar] [CrossRef]
- Zoghbi, M.G.B.; Andrade, E.H.A.; Maia, J.G.S. Aroma de flores na Amazônia, Museu Paraense Emílio Goeldi. Série Adolpho Ducke: Belém, Brasil, 2001; p. 240. [Google Scholar]
- Pisseri, F.; Bertoli, A.; Pistelli, L. Essential oils in medicine: Principles of therapy. Parassitologia 2008, 50, 89–91. [Google Scholar]
- Edris, A.E. Pharmaceutical and therapeutic potentials of essential oils and their individual volatile constituents: A review. Phytother. Res. 2007, 21, 308–323. [Google Scholar] [CrossRef]
- Chang, R.; Morais, S.A.L.; Nascimento, E.A.; Cunha, L.C.S.; Rocha, E.O.; Aquino, F.J.T.; Souza, M.G.M.; Cunha, W.R.; Martins, C.H.G. Essential oil composition and antioxidant and antimicrobial properties of Campomanesia pubescens O. Berg, Native of Brazilian Cerrado. Lat. Am. J. Pharm. 2011, 30, 1843–1848. [Google Scholar]
- Cunha, L.C.S.; Morais, S.A.L.; Martins, C.H.G.; Martins, M.M.; Chang, R.; Aquino, F.J.T.; Oliveira, A.; Moraes, T.S.; Machado, F.C.; Silva, C.V.; et al. Chemical composition, cytotoxic and antimicrobial activity of essential oils from Cassia bakeriana Craib. against aerobic and anaerobic oral pathogens. Molecules 2013, 18, 4588–4598. [Google Scholar] [CrossRef]
- Rezende, C.M.; Corrêa, V.F.S.; Costa, A.V.M.; Castro, B.C.S. Constituintes químicos voláteis das flores e folhas do pau-Brasil (Caesalpinia echinata, Lam.). Quím. Nova 2004, 27, 414–416. [Google Scholar]
- NIST Standard Reference Data. Available online: http://webbook.nist.gov/chemistry/name-ser.html (accessed on 15 January 2013).
- Adams, R.P. Identification of Essential Oil Components by Gas Chromatography/Quadrupole Mass Spectroscopy, 4th ed.Allured Publishing Corporation: Carol Stream, IL, USA, 2007; p. 803. [Google Scholar]
- Van Poecke, R.M.P.; Posthumus, M.A.; Dicke, M. Herbivore-induced volatile production by Arabidopsis thaliana leads to attraction of the parasitoid Cotesia rubecula: Chemical, behavioral, and gene-expression analysis. J. Chem. Ecol. 2001, 27, 1911–1928. [Google Scholar] [CrossRef]
- THE PHEROBASE. Database of Insect Pheromones and Semiochemicals. Available online: http://www.pherobase.com (accessed on 8 January 2013).
- McCaskill, D.; Croteau, R. Strategies for bioengineering the development and metabolism of glandular tissues in plants. Nat. Biotechnol. 1998, 17, 31–37. [Google Scholar]
- Yamaura, T.; Tanaka, S.; Tabata, M. Light-dependent formation of glandular trichomes and monoterpenes in thyme seedlings. Phytochemistry 1989, 28, 741–744. [Google Scholar] [CrossRef]
- Ioannidis, D.; Bonner, L.; Johnson, C.B. UV-B is required for normal development of oil glands in Ocimum basilicum L. (Sweet Basil). Ann. Bot. 2002, 90, 453–460. [Google Scholar] [CrossRef]
- Heredia, A.; Domínguez, E.; Serrano, J.M.; Laguna, L.; Reina, J.J.; Casado, C.G. La cutícula vegetal: Estructura y funciones. Ecología 1998, 12, 293–305. [Google Scholar]
- Gazim, Z.C.; Amorim, A.C.L.; Hovell, A.M.C.; Rezende, C.M.; Nascimento, I.A.; Ferreira, G.A.; Cortez, D.A.G. Seasonal variation, chemical composition, and analgesic and antimicrobial activities of the essential oils from leaves of Tetradenia riparia (Hochst.) Codd. in Southern Brazil. Molecules 2010, 15, 5509–5524. [Google Scholar] [CrossRef]
- Cardoso, M.G.; Shan, A.Y.K.V.; Pinto, J.E.B.P.; Delu Filho, N.; Bertolucci, S.K.V. Metabólitos Secundários Vegetais: Visão Geral, Química e Medicinal. Editora UFLA-Lavras: Lavras-MG, Brasil, 2001; p. 81. [Google Scholar]
- Rios, J.L.; Recio, M.C. Medicinal plants and antimicrobial activity. J. Ethnopharmacol. 2005, 100, 80–84. [Google Scholar] [CrossRef]
- Ouattara, B.; Simard, R.E.; Holley, R.A.; Piette, G.J.P.; Bégin, A. Antibacterial activity of selected fatty acids and essential oils against six meat spoilage organisms. Int. J. Food Microbiol. 1997, 37, 155–162. [Google Scholar] [CrossRef]
- Desbois, A.P.; Smith, V.J. Antibacterial free fatty acids: Activities, mechanisms of action and biotechnological potential. Appl. Microbiol. Biotechnol. 2010, 85, 1629–1642. [Google Scholar] [CrossRef] [Green Version]
- Branen, A.L.; Davidson, P.M.; Katz, B. Antibacterial properties of phenolic antioxidants and lipids. Food Technol. 1980, 34, 51–53. [Google Scholar]
- Wang, L.; Johnson, E.A. Inhibition of Listeria monocytogenes by fatty acids and monoglycerides. Appl. Environ Microbiol. 1992, 58, 624–629. [Google Scholar]
- Carvalho, T.C.; Simão, M.R.; Ambrósio, S.R.; Furtado, N.A.; Veneziani, R.C.; Heleno, V.C.; da Costa, F.B.; Gomes, B.P.; Souza, M.G.; Reis, R.B.; et al. Antimicrobial activity of diterpenes from Viguiera arenaria against endodontic bacteria. Molecules 2011, 16, 543–551. [Google Scholar] [CrossRef]
- Botelho, M.A.; Nogueira, N.A.; Bastos, G.M.; Fonseca, S.G.; Lemos, T.L.; Matos, F.J.; Montenegro, D.; Heukelbach, J.; Rao, V.S.; Brito, G.A. Antimicrobial activity of the essential oil from Lippia sidoides, carvacrol and thymol against oral pathogens. Braz. J. Med. Biol. Res. 2007, 40, 349–356. [Google Scholar] [CrossRef]
- Xiong, L.; Peng, C.; Zhou, Q.M.; Wan, F.; Xie, X.F.; Guo, L.; Li, X.H.; He, C.J.; Dai, O. Chemical composition and antibacterial activity of essential oils from different parts of Leonurus japonicus Houtt. Molecules 2013, 18, 963–973. [Google Scholar] [CrossRef]
- Dorman, H.J.D.; Deans, S.G. Antimicrobial agents from plants: Antibacterial activity of plant volatile oils. J. Appl. Microbiol. 2000, 88, 308–316. [Google Scholar] [CrossRef]
- Skaltsa, H.D.; Lazzari, D.M.; Mavromati, A.S.; Tiligada, E.A.; Constantinidis, T.A. Composition and antimicrobial activity of the essential oil of Scutellaria albida ssp. albida from Greece. Planta Med. 2000, 66, 672–674. [Google Scholar] [CrossRef]
- Pavithra, P.S.; Sreevidya, N.; Verma, R.S. Antibacterial activity and chemical composition of essential oil of Pamburus missionis. J. Ethnopharmacol. 2009, 124, 151–153. [Google Scholar] [CrossRef]
- Inoue, Y.; Hada, T.; Shiraishi, A.; Hirose, K.; Hamashima, H.; Kobayashi, S. Biphasic Effects of Geranylgeraniol, Teprenone, and Phytol on the Growth of Staphylococcus aureus. Antimicrob. Agents Chemother. 2005, 49, 1770–1774. [Google Scholar] [CrossRef]
- Yoon, W.J.; Kim, S.S.; Oh, T.H.; Lee, N.H.; Hyun, C.G. Cryptomeria japonica essential oil inhibits the growth of drug-resistant skin pathogens and LPS-induced nitric oxide and pro-inflammatory cytokine production. Pol. J. Microbiol. 2009, 58, 61–68. [Google Scholar]
- Rath, C.C.; Devi, S.; Dash, S.K.; Mishra, R. Antibacterial potential assessment of Jasmine essential oil against E. coli. Indian J. Pharm. Sci. 2008, 70, 238–241. [Google Scholar] [CrossRef]
- Ulusoy, S.; Bosgelmez-Tinaz, G.; Seçilmis-Canbay, H. Tocopherol, carotene, phenolic contents and antibacterial properties of rose essential oil, hydrosol and absolute. Curr. Microbiol. 2009, 59, 554–558. [Google Scholar] [CrossRef]
- Takarada, K.; Kimizuka, R.; Takarashi, R.; Honma, K.; Okuda, K.; Kato, T. A comparison of the antibacterial efficacies of essential oils against oral pathogens. Oral Microbiol. Immunol. 2004, 19, 61–64. [Google Scholar] [CrossRef]
- Cha, J.D. Chemical composition and antibacterial activity against oral bacteria by the essential oil of Artemisia iwayomogi. J. Bacteriol. Virol. 2007, 37, 129–136. [Google Scholar] [CrossRef]
- Galvão, L.C.C.; Furletti, V.F.; Bersan, S.M.F.; Cunha, M.G.; Ruiz, A.L.T.G.; Carvalho, J.E.; Sartoratto, A.; Rehder, V.L.G.; Figueira, G.M.; Duarte, M.C.T.; et al. Antimicrobial activity of essential oils against Streptococcus mutans and their antiproliferative effects. Evid. Based Complement. Alternat. Med. 2012, 2012, 1–12. [Google Scholar]
- Case, R.J.; Franzblau, S.G.; Wang, Y.; Cho, S.H.; Soejarto, D.D.; Pauli, G.F. Ethnopharmacological evaluation of the informant consensus model on anti-tuberculosis claims among the Manus. J. Ethnopharmacol. 2006, 106, 82–89. [Google Scholar] [CrossRef]
- Van den Dool, H.; Kratz, P.D. A generalization of the retention index system including linear temperature programmed gas–liquid partition chromatography. J. Chromatogr. 1963, 11, 463–471. [Google Scholar] [CrossRef]
- National Committee for Clinical Laboratory Standards (NCCLS). NCCLS Document M7-A6—Methods for Dilution Antimicrobial Susceptibility Tests for Bacteria that Grow Aerobically. NCCLS: Wayne, PA, USA, 2003. [Google Scholar]
- Gómez-Barrio, A.; Veja, C.; Escario, J.A.; Rolón, M. Development of resazurin microtiter assay for drug sensibility testing of Trypanosoma cruzi epimastigotes. Parasitol. Res. 2006, 99, 103–107. [Google Scholar] [CrossRef]
- Chibale, K.; Chouteau, F.; Lategan, C.A.; Maharaj, V.J.; Pillay, P.; Smith, P.J.; Vleggaar, R. Antiplasmodial hirsutinolides from Vernonia staehelinoides and their utilization towards a simplified pharmacophore. Phytochemistry 2007, 68, 1200–1205. [Google Scholar] [CrossRef]
- Sample Availability: Samples of the essential oils of Inga laurina are available from the authors.
© 2014 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license ( http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Furtado, F.B.; De Aquino, F.J.T.; Nascimento, E.A.; Martins, C.D.M.; De Morais, S.A.L.; Chang, R.; Cunha, L.C.S.; Leandro, L.F.; Martins, C.H.G.; Martins, M.M.; et al. Seasonal Variation of the Chemical Composition and Antimicrobial and Cytotoxic Activities of the Essential Oils from Inga laurina (Sw.) Willd. Molecules 2014, 19, 4560-4577. https://doi.org/10.3390/molecules19044560
Furtado FB, De Aquino FJT, Nascimento EA, Martins CDM, De Morais SAL, Chang R, Cunha LCS, Leandro LF, Martins CHG, Martins MM, et al. Seasonal Variation of the Chemical Composition and Antimicrobial and Cytotoxic Activities of the Essential Oils from Inga laurina (Sw.) Willd. Molecules. 2014; 19(4):4560-4577. https://doi.org/10.3390/molecules19044560
Chicago/Turabian StyleFurtado, Fabiana B., Francisco J. T. De Aquino, Evandro A. Nascimento, Carla De M. Martins, Sérgio A. L. De Morais, Roberto Chang, Luís C. S. Cunha, Luís F. Leandro, Carlos H. G. Martins, Mário M. Martins, and et al. 2014. "Seasonal Variation of the Chemical Composition and Antimicrobial and Cytotoxic Activities of the Essential Oils from Inga laurina (Sw.) Willd." Molecules 19, no. 4: 4560-4577. https://doi.org/10.3390/molecules19044560