Deguelin Inhibits the Migration and Invasion of U-2 OS Human Osteosarcoma Cells via the Inhibition of Matrix Metalloproteinase-2/-9 in Vitro


Abstract
:1. Introduction
2. Results and Discussion
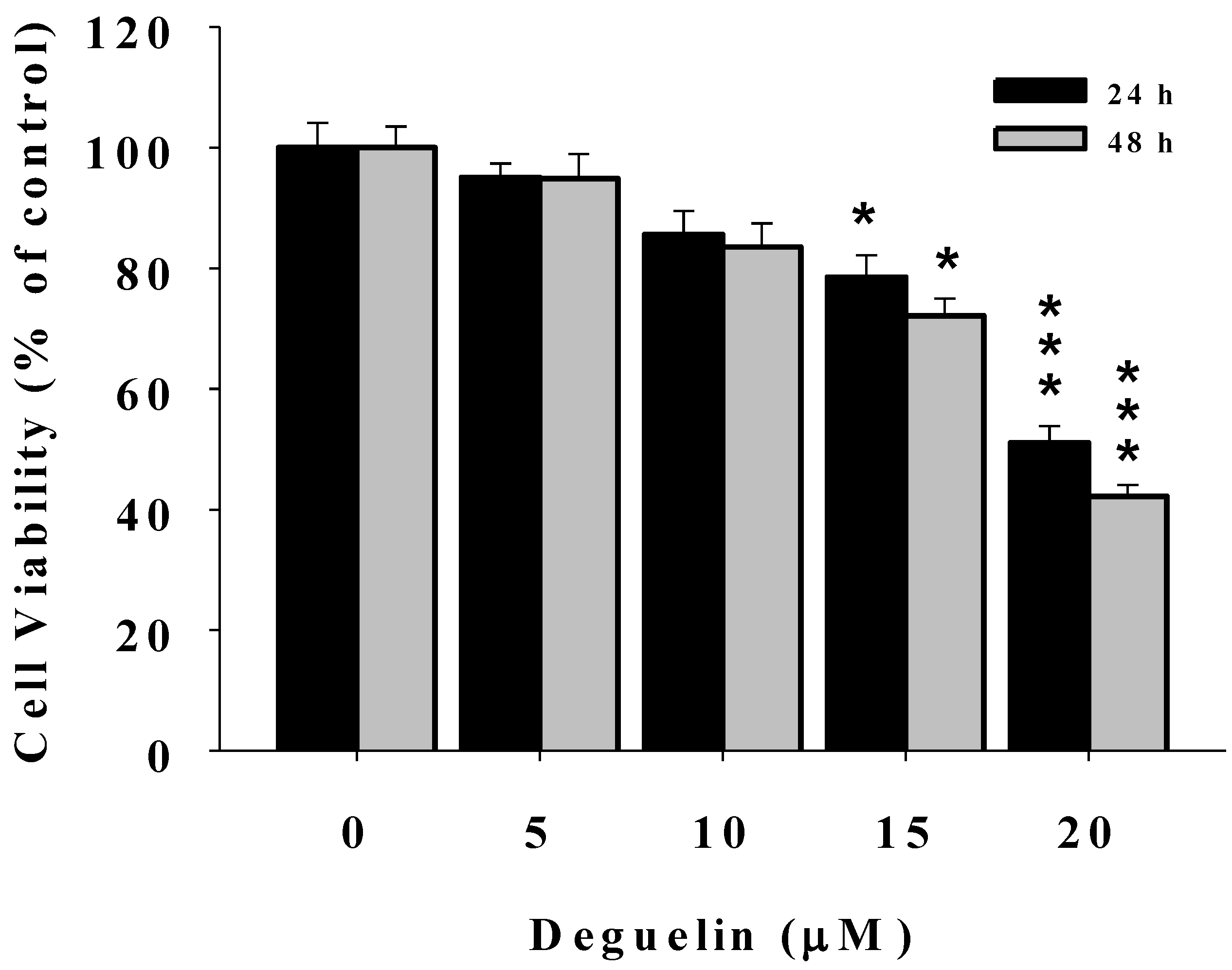
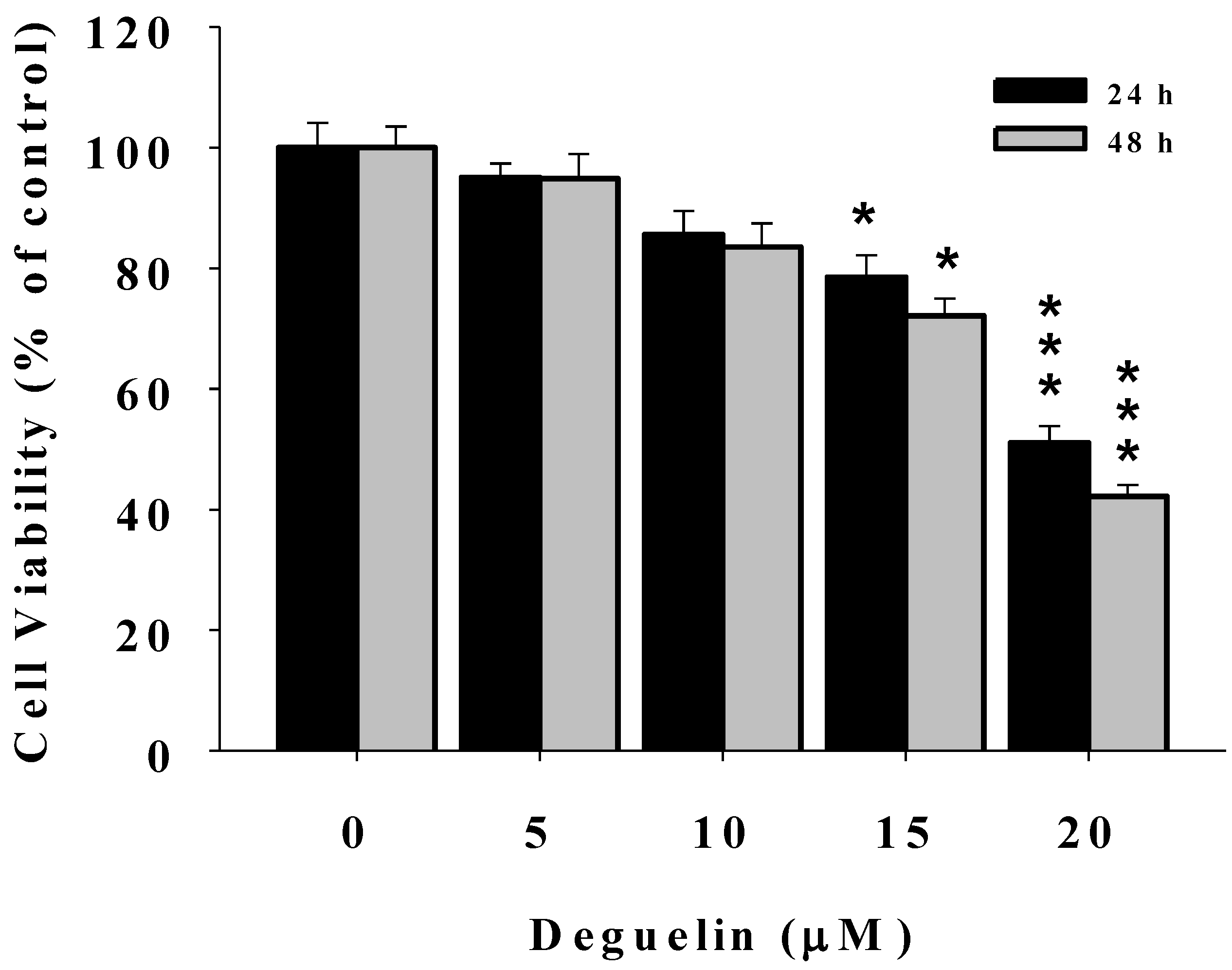
2.1. Deguelin Reduces the Percentage of Viable U2-OS Human Osteosarcoma Cells
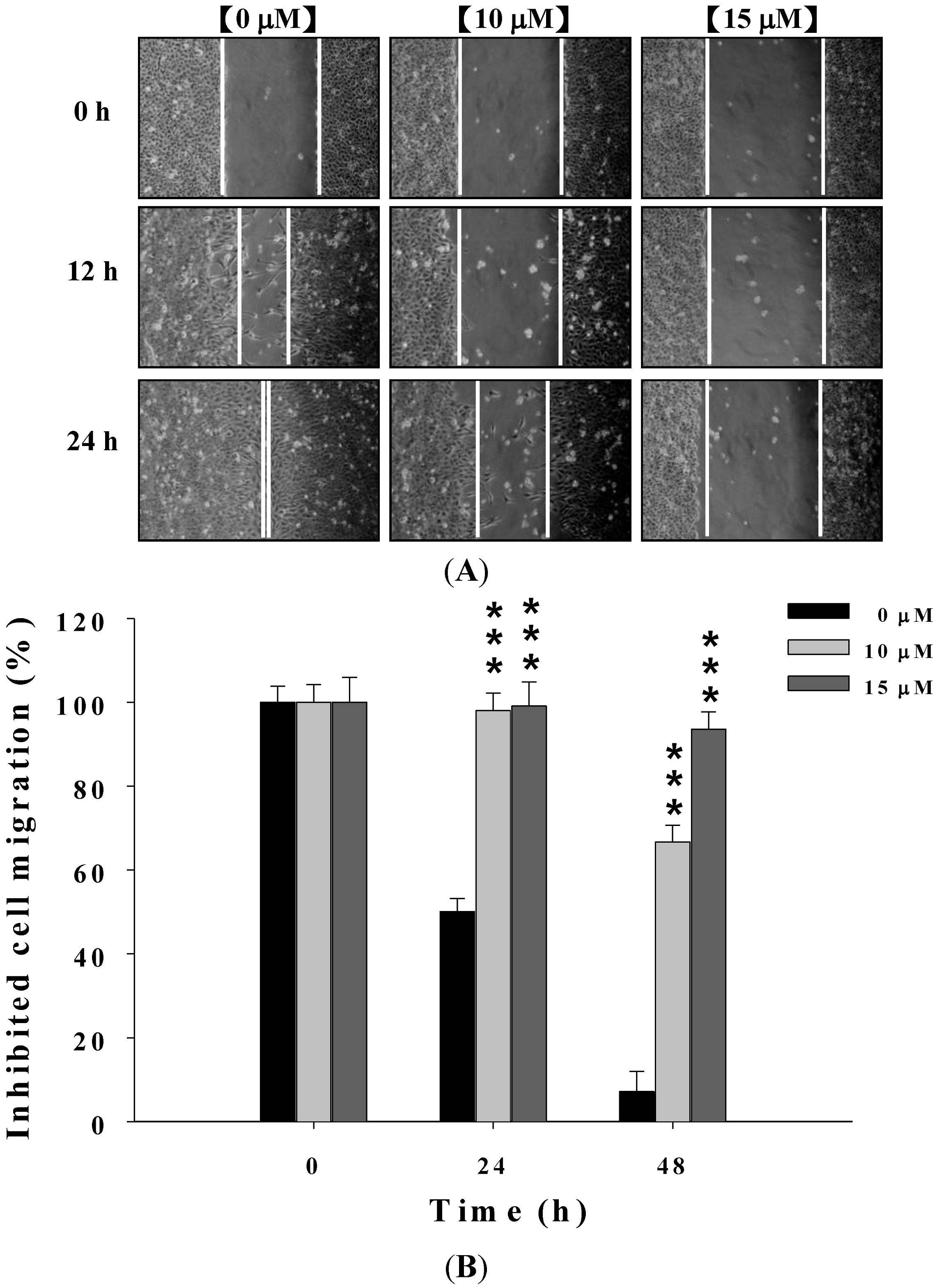
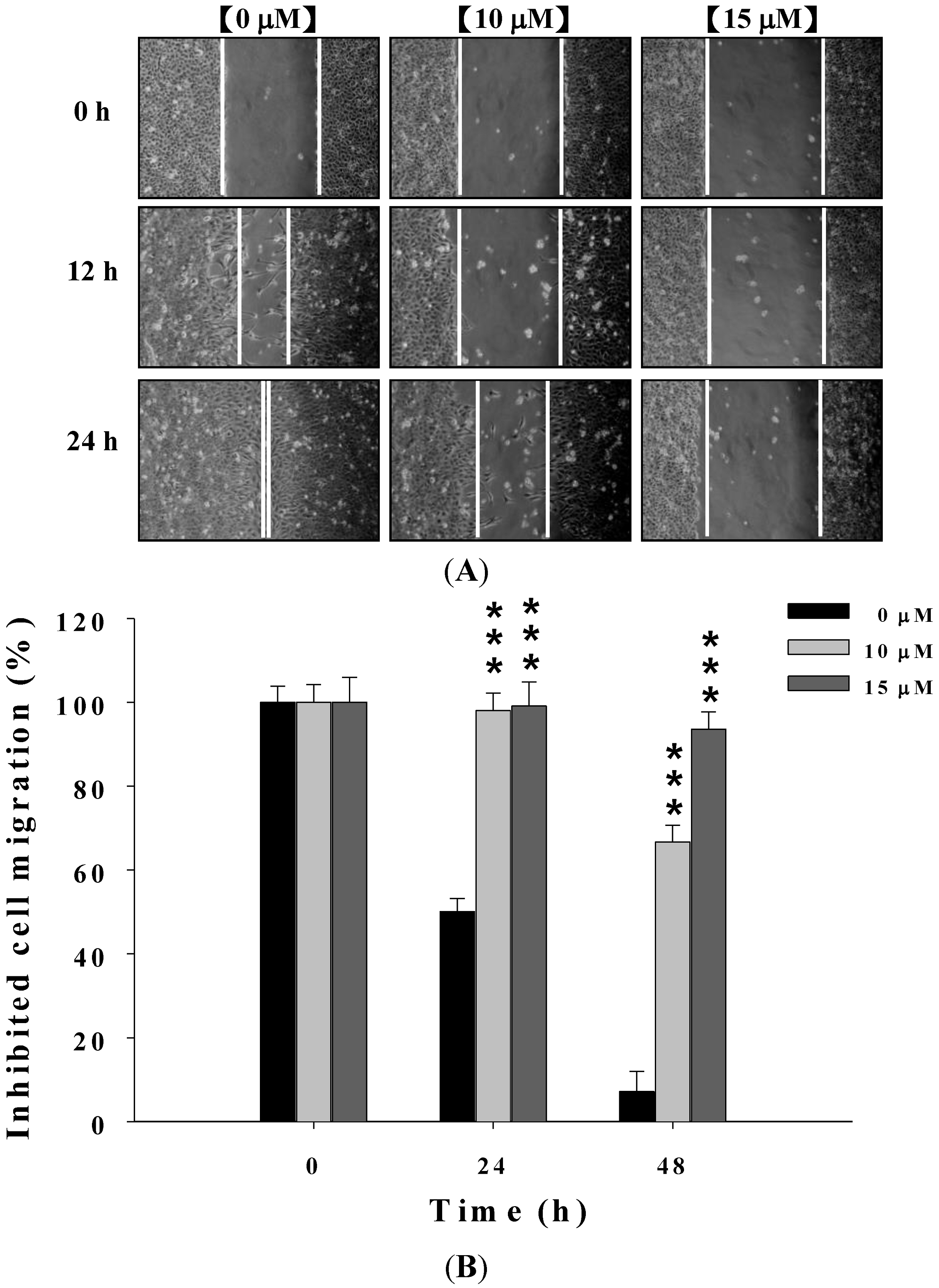
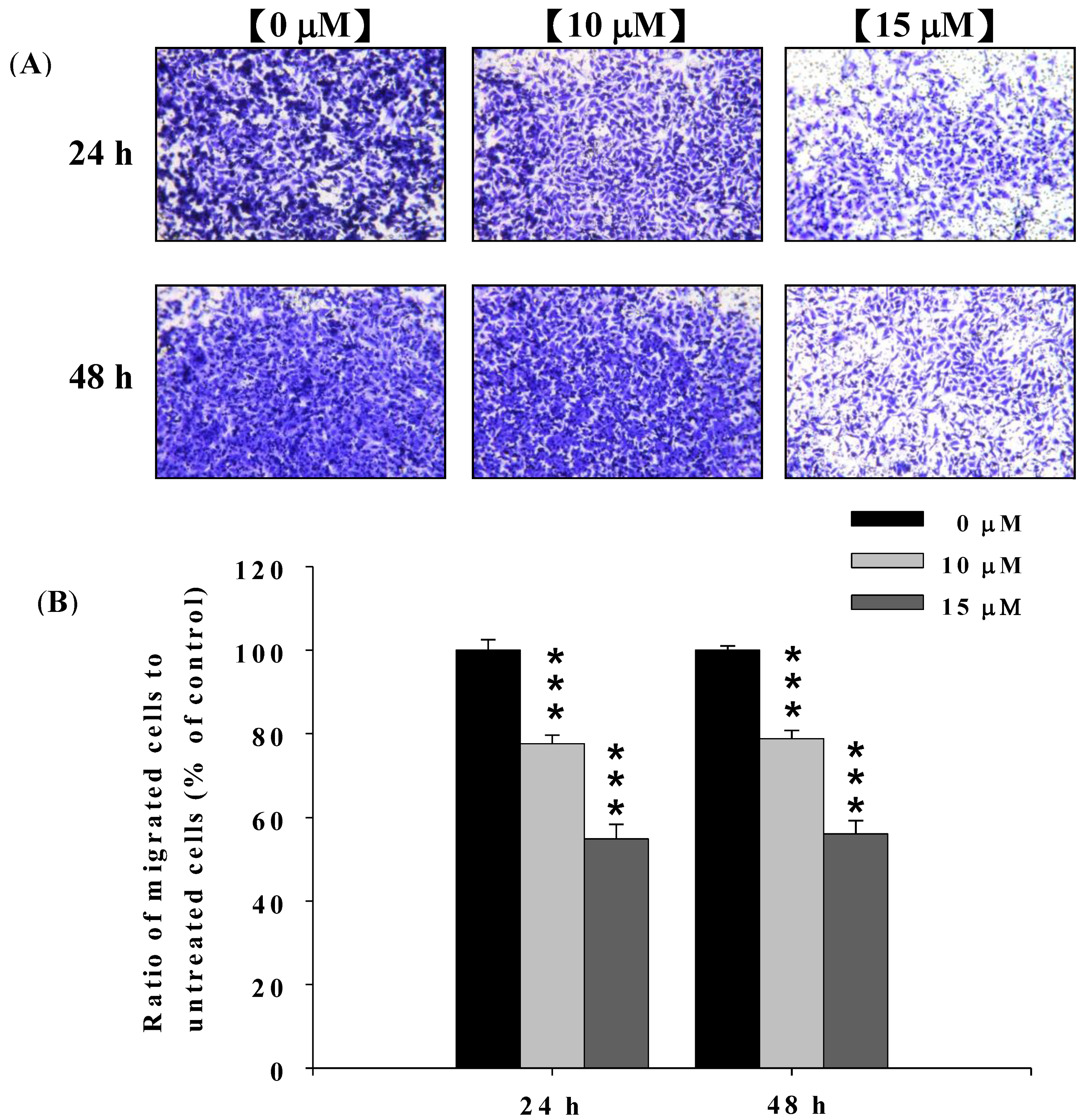
2.2. Deguelin Suppresses the Migration of U-2 OS Cells in Vitro
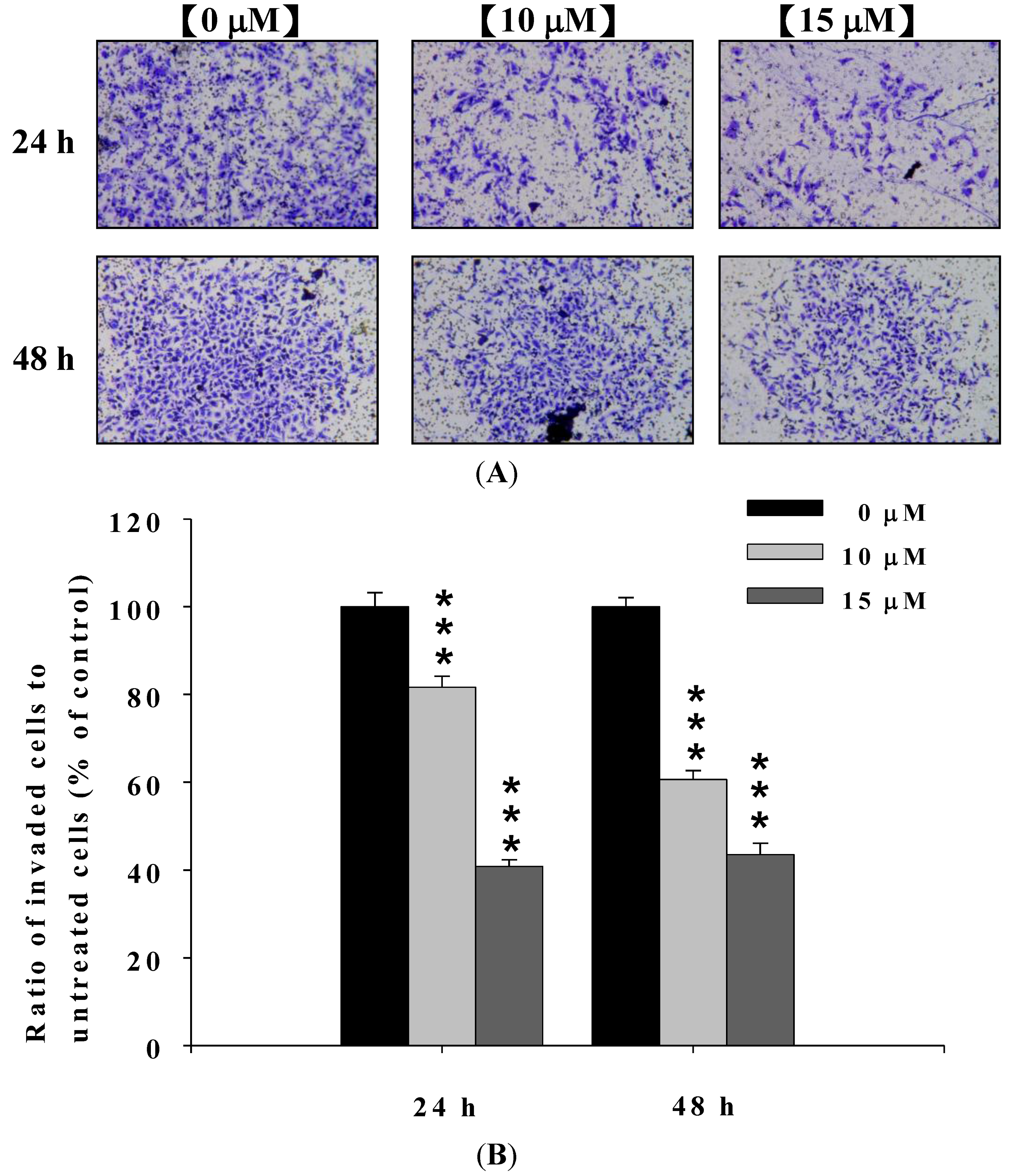
2.3. Deguelin Inhibited the Migration and Invasion of U2-OS Cells in Vitro
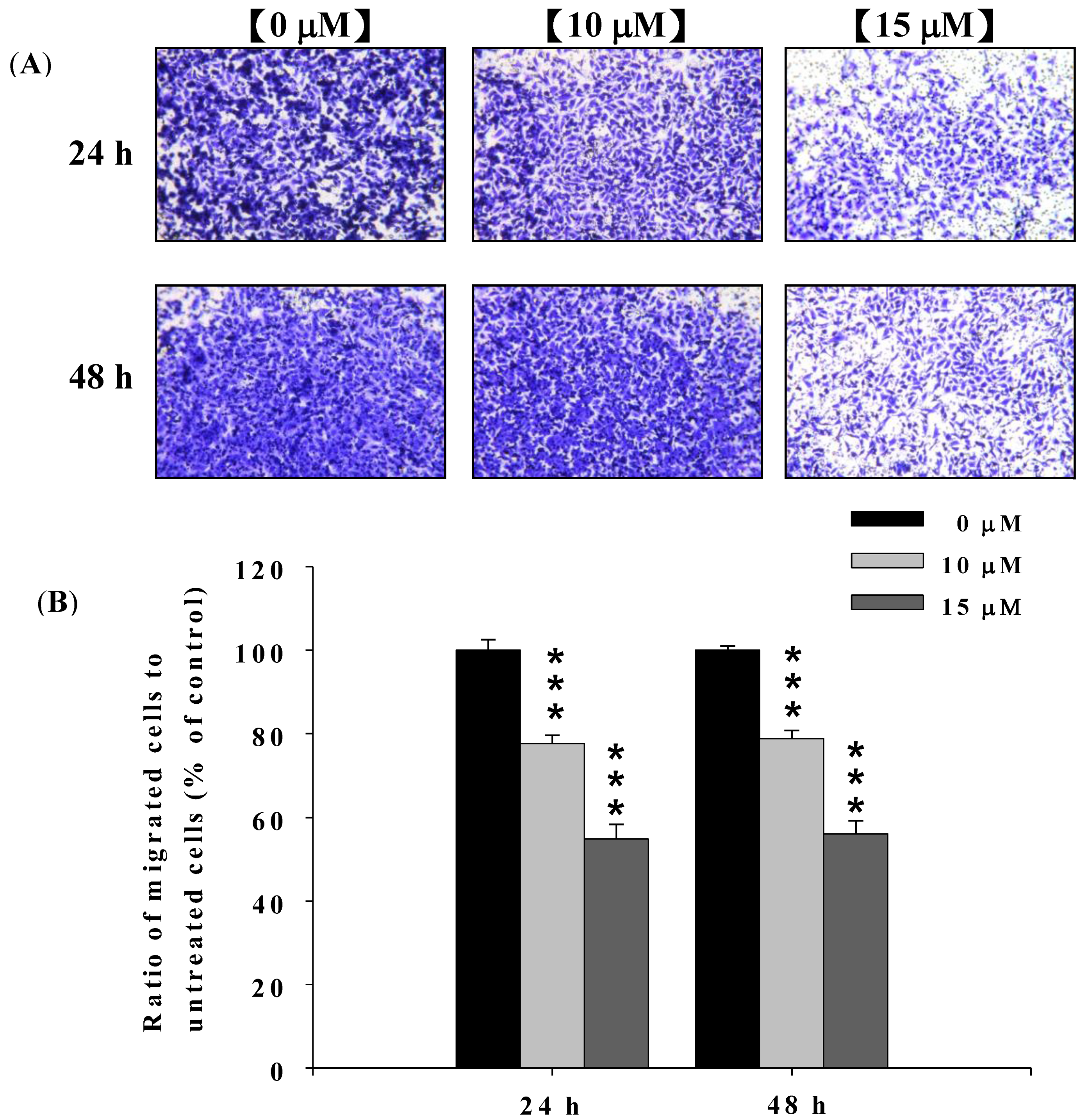
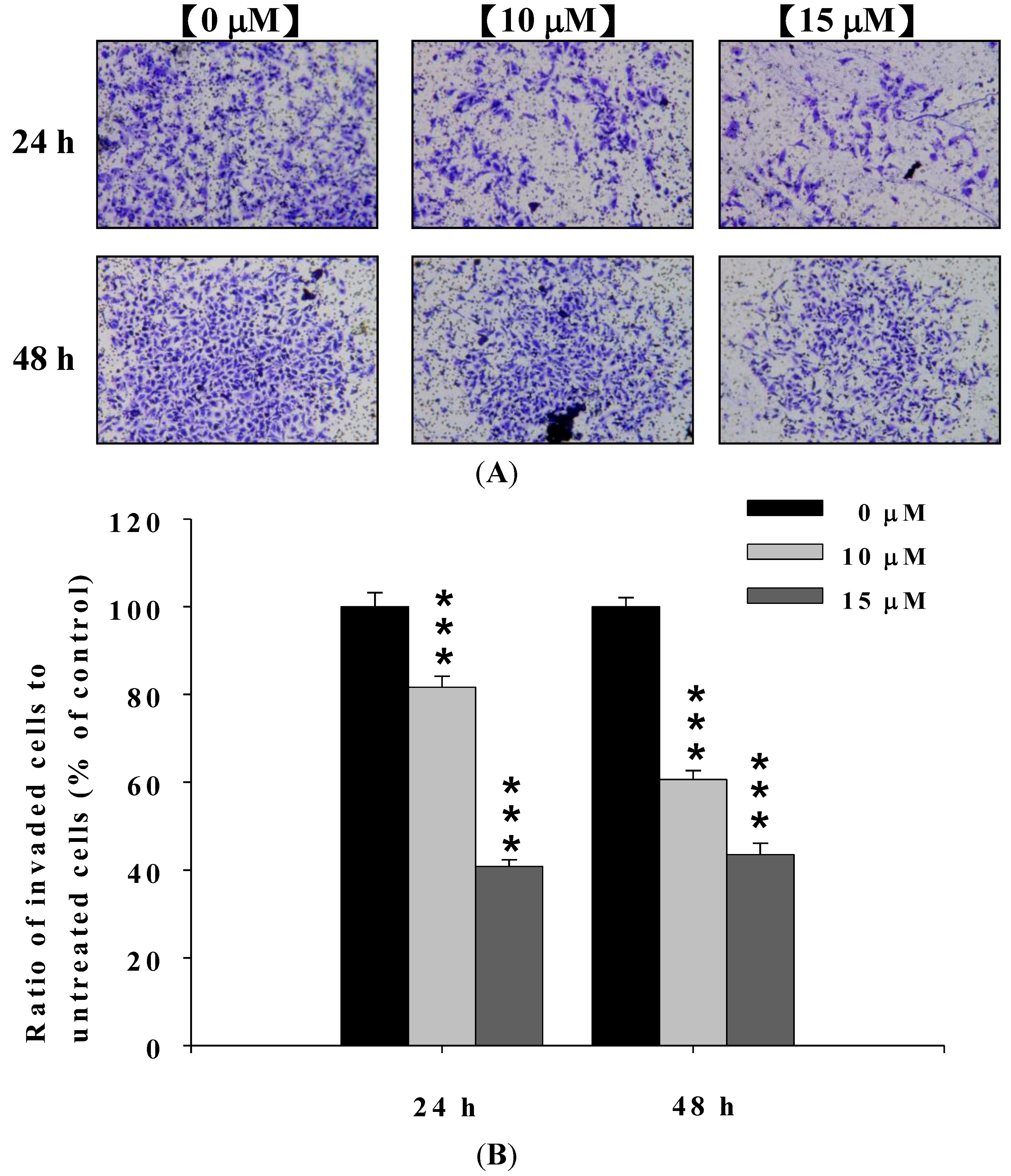
2.4. Deguelin Inhibited the Matrix Metalloproteinases-2 and -9 Activities in U-2 OS Cells

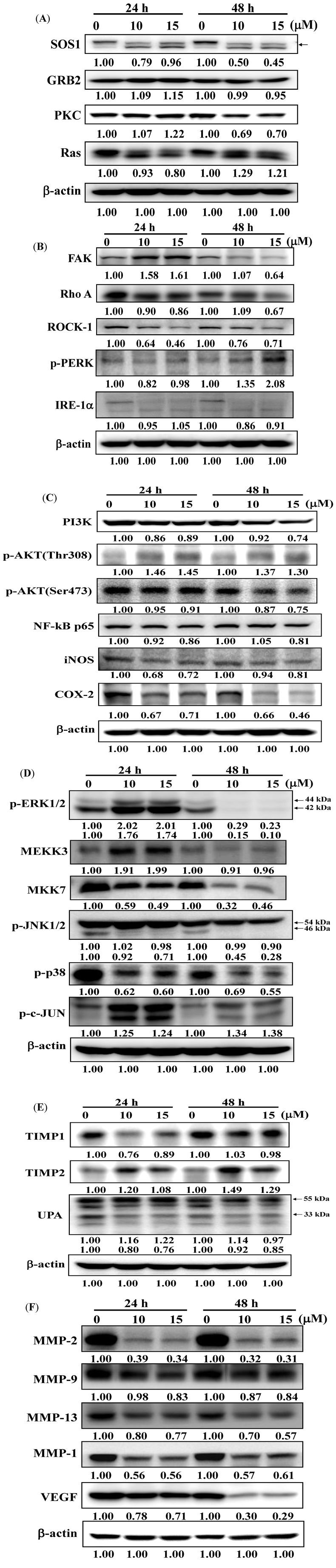
2.5. Deguelin Alters Levels of Proteins Associated with Migration and Invasion in U-2 OS Cells
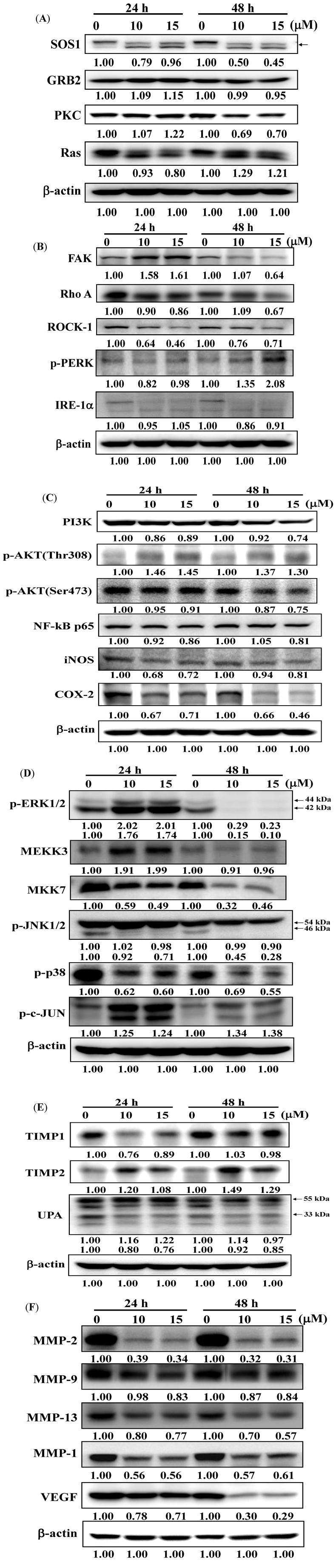
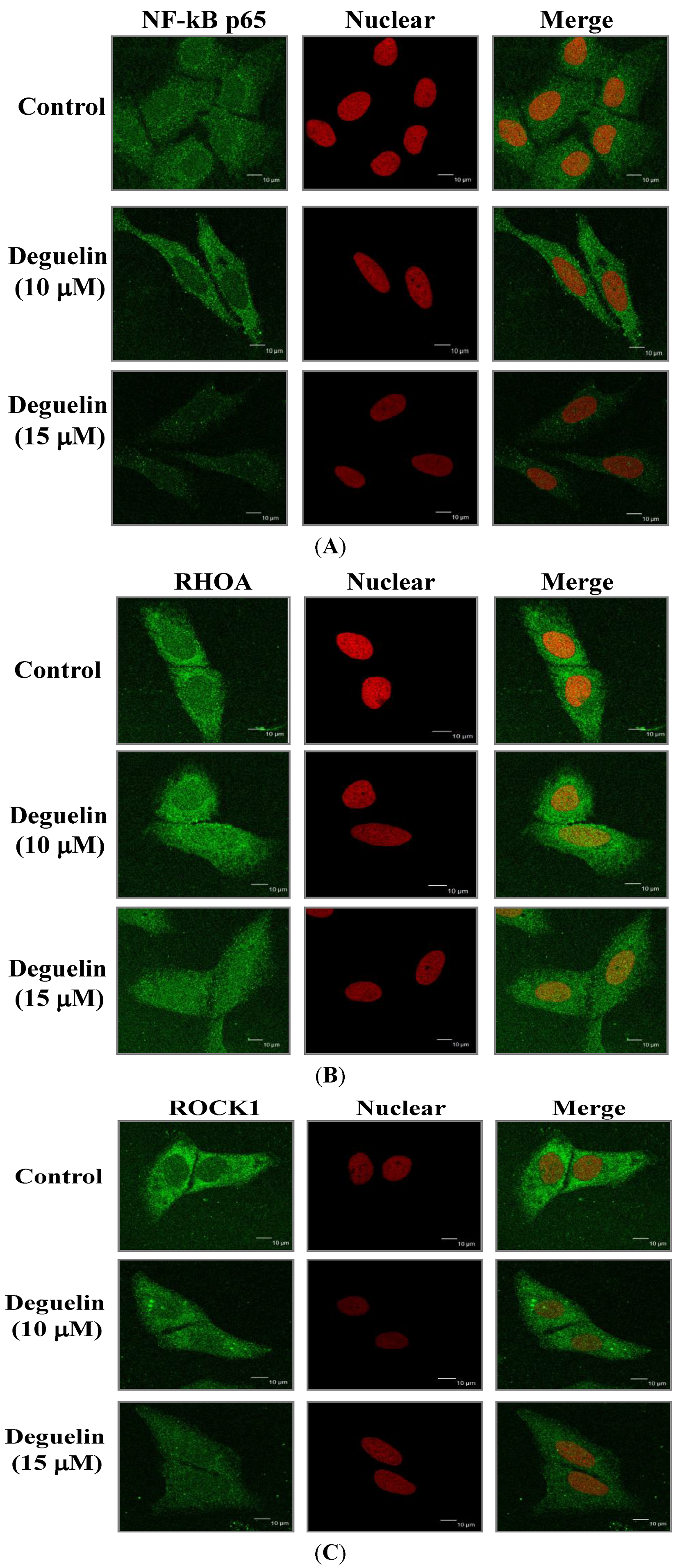
2.6. Effects of Deguelin on Protein Translocation

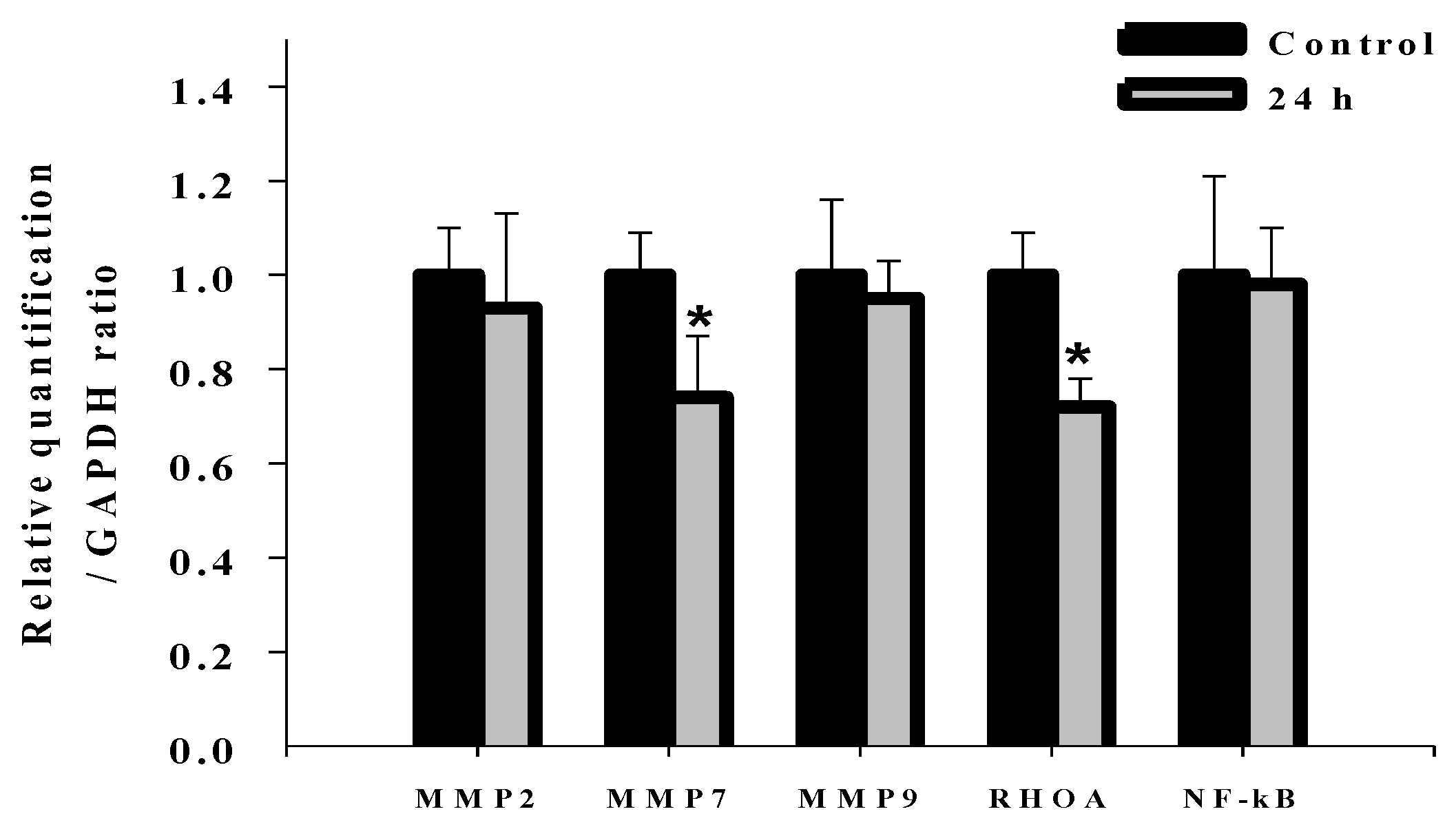
2.7. Effects of Deguelin on mRNA Expression of MMP-2, MMP-7, MMP-9, Rho A and NF-kB in U-2 OS Cells
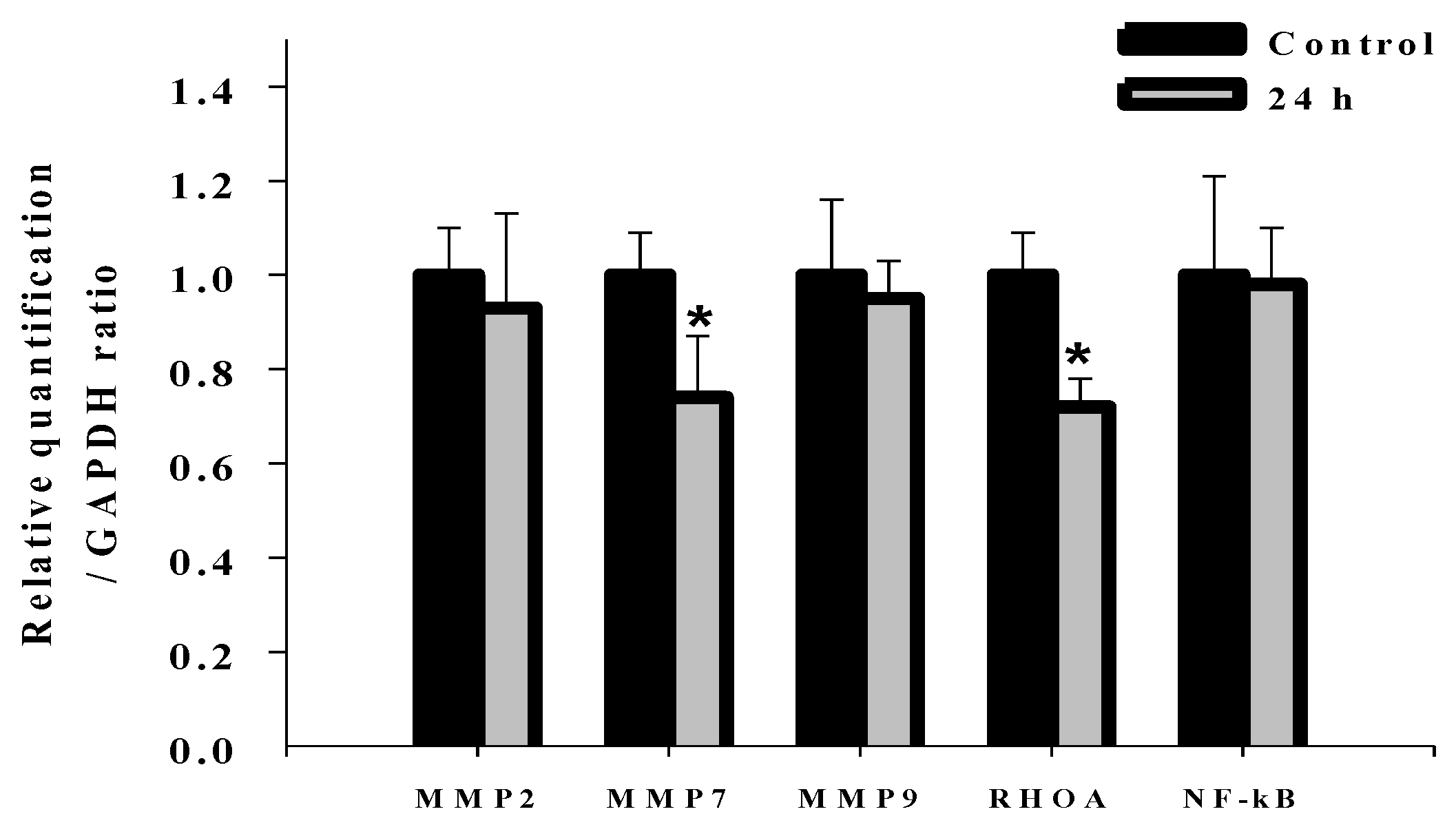
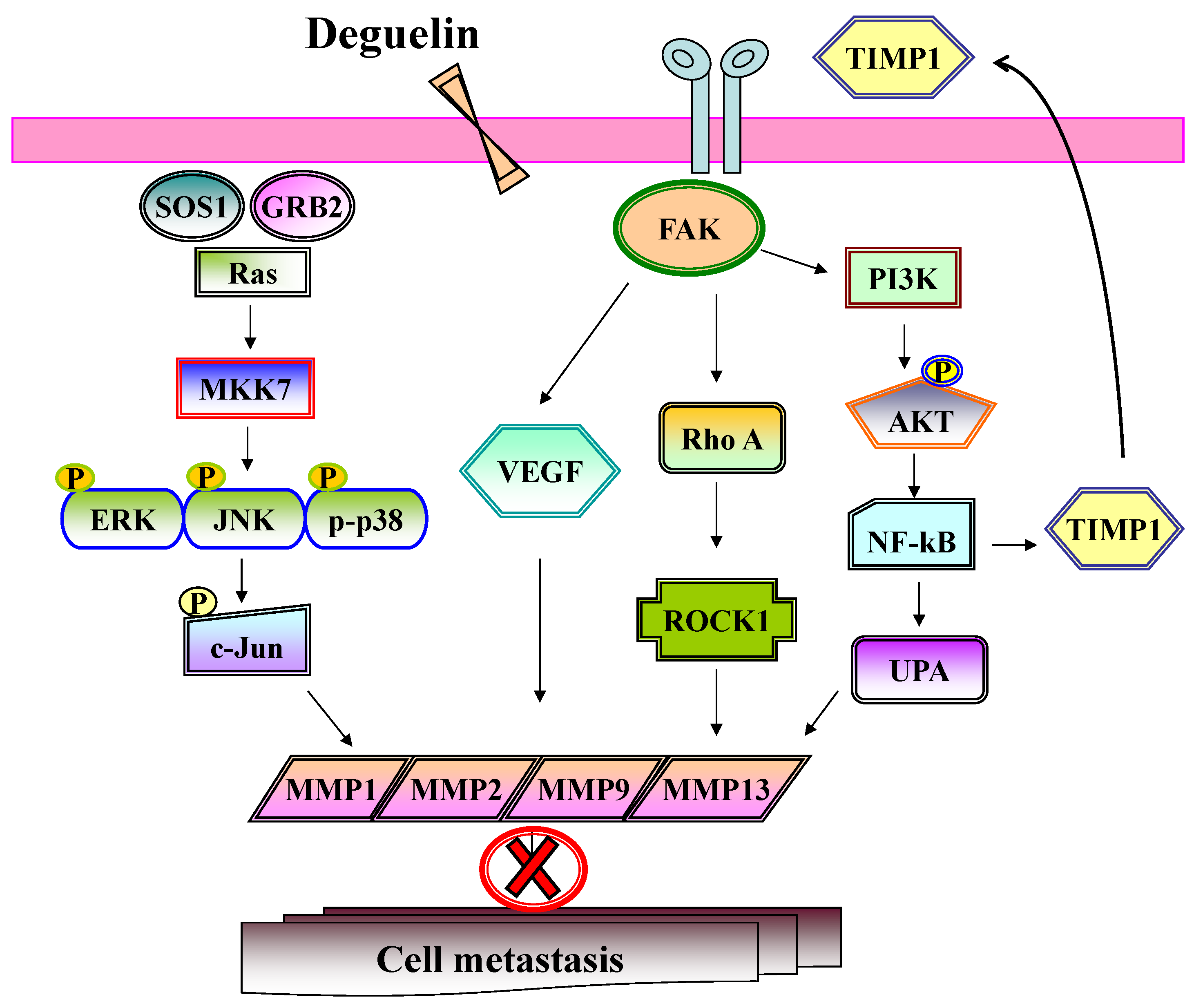
2.8. Discussion
3. Experimental Section
3.1. Chemicals and Reagents
3.2. U-2 OS Cell Culture
3.3. Cell Viability Assay
3.4. Scratch Wound Healing Assay
3.5. Cell Invasion and Migration Assays in Vitro
3.6. Gelatin Gel Zymographic Assay for MMP Activity
3.7. Immunofluorescence Staining and Confocal Laser Scanning Microscopy
3.8. Western Blotting Analysis
3.9. Real-time PCR of MMP-2, -7, and -9, Rho A and NF-kB

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Primer Name | Primer Sequence |
|---|---|
| homo MMP-2-F | CCCCAGACAGGTGATCTTGAC |
| homo MMP-2-R | GCTTGCGAGGGAAGAAGTTG |
| homo MMP-7-F | GGATGGTAGCAGTCTAGGGATTAACT |
| homo MMP-7-R | AGGTTGGATACATCACTGCATTAGG |
| homo MMP-9-F | CGCTGGGCTTAGATCATTCC |
| homo MMP-9-R | AGGTTGGATACATCACTGCATTAGG |
| homo NF-κB-F | AGTTGAGGGGACTTTCCCAGGC |
| homo NF-κB-R | TCAACTCCCCTGAAAGGGTCCG |
| homo RhoA-F | TCAAGCCGGAGGTCAACAAC |
| homo RhoA-R | ACGAGCTGCCCATAGCAGAA |
| Homo GAPDH-F | ACACCCACTCCTCCACCTTT |
| Homo GAPDH-R | TAGCCAAATTCGTTGTCATACC |
3.10. Statistical Analysis
4. Conclusions

Acknowledgments
Author Contributions
Conflicts of Interest
References
- Sangle, N.A.; Layfield, L.J. Telangiectatic osteosarcoma. Arch. Pathol. Lab. Med. 2012, 136, 572–576. [Google Scholar]
- Huh, W.W.; Holsinger, F.C.; Levy, A.; Palla, F.S.; Anderson, P.M. Osteosarcoma of the jaw in children and young adults. Head Neck 2012, 34, 981–984. [Google Scholar]
- Kaste, S.C. Imaging pediatric bone sarcomas. Radiol. Clin. North Am. 2011, 49, 749–765. [Google Scholar]
- Thayanithy, V.; Park, C.; Sarver, A.L.; Kartha, R.V.; Korpela, D.M.; Graef, A.J.; Steer, C.J.; Modiano, J.F.; Subramanian, S. Combinatorial treatment of DNA and chromatin-modifying drugs cause cell death in human and canine osteosarcoma cell lines. PLoS One 2012, 7, e43720. [Google Scholar]
- Fuchs, B.; Pritchard, D.J. Etiology of osteosarcoma. Clin. Orthop. Relat. Res. 2002, 397, 40–52. [Google Scholar]
- Marina, N.; Gebhardt, M.; Teot, L.; Gorlick, R. Biology and therapeutic advances for pediatric osteosarcoma. Oncologist 2004, 9, 422–441. [Google Scholar]
- Geller, D.S.; Gorlick, R. Osteosarcoma: A review of diagnosis, management, and treatment strategie. Clin. Adv. Hematol. Oncol. 2010, 8, 705–718. [Google Scholar]
- Olson, M.F. Follow the leader: LIM kinases pave the way for collective tumor cell invasion. Cell Cycle 2010, 9, 4417–4418. [Google Scholar]
- Thompson, L.; Wang, S.; Tawfik, O.; Templeton, K.; Tancabelic, J.; Pinson, D.; Anderson, H.C.; Keighley, J.; Garimella, R. Effect of 25-hydroxyvitamin D3 and 1 alpha,25 dihydroxyvitamin D3 on differentiation and apoptosis of human osteosarcoma cell lines. J. Orthop. Res. 2012, 30, 831–844. [Google Scholar]
- Benayahu, D.; Shur, I.; Marom, R.; Meller, I.; Issakov, J. Cellular and molecular properties associated with osteosarcoma cells. J. Cell. Biochem. 2001, 84, 108–114. [Google Scholar]
- Campbell, P.G.; Wines, K.; Yanosick, T.B.; Novak, J.F. Binding and activation of plasminogen on the surface of osteosarcoma cells. J. Cell. Physiol. 1994, 159, 1–10. [Google Scholar]
- Smeds, S.; Walz, T.; Blomquist, L.; Larsson, S.E. Cell proliferation rate and tumor volume in human osteosarcoma during exposure to methotrexate. A study on tissue transplants in nude mice. APMIS 1991, 99, 898–904. [Google Scholar]
- Tingting, R.; Wei, G.; Changliang, P.; Xinchang, L.; Yi, Y. Arsenic trioxide inhibits osteosarcoma cell invasiveness via MAPK signaling pathway. Cancer Biol. Ther. 2010, 10, 251–257. [Google Scholar]
- Sun, D.; Wang, X.; Zhang, H.; Deng, L.; Zhang, Y. MMP9 mediates MICA shedding in human osteosarcomas. Cell Biol. Int. 2011, 35, 569–574. [Google Scholar]
- Berge, G.; Pettersen, S.; Grotterod, I.; Bettum, I.J.; Boye, K.; Maelandsmo, G.M. Osteopontin—An important downstream effector of S100A4-mediated invasion and metastasis. Int. J. Cancer. 2011, 129, 780–790. [Google Scholar]
- Korpi, J.T.; Hagstrom, J.; Lehtonen, N.; Parkkinen, J.; Sorsa, T.; Salo, T.; Laitinen, M. Expression of matrix metalloproteinases-2, -8, -13, -26, and tissue inhibitors of metalloproteinase-1 in human osteosarcoma. Surg. Oncol. 2011, 20, e18–e22. [Google Scholar]
- Lee, H.Y.; Oh, S.H.; Woo, J.K.; Kim, W.Y.; van Pelt, C.S.; Price, R.E.; Cody, D.; Tran, H.; Pezzuto, J.M.; Moriarty, R.M.; et al. Chemopreventive effects of deguelin, a novel Akt inhibitor, on tobacco-induced lung tumorigenesis. J. Natl. Cancer Inst. 2005, 97, 1695–1699. [Google Scholar]
- Udeani, G.O.; Gerhauser, C.; Thomas, C.F.; Moon, R.C.; Kosmeder, J.W.; Kinghorn, A.D.; Moriarty, R.M.; Pezzuto, J.M. Cancer chemopreventive activity mediated by deguelin, a naturally occurring rotenoid. Cancer Res. 1997, 57, 3424–3428. [Google Scholar]
- Gerhauser, C.; Lee, S.K.; Kosmeder, J.W.; Moriarty, R.M.; Hamel, E.; Mehta, R.G.; Moon, R.C.; Pezzuto, J.M. Regulation of ornithine decarboxylase induction by deguelin, a natural product cancer chemopreventive agent. Cancer Res. 1997, 57, 3429–3435. [Google Scholar]
- Murillo, G.; Kosmeder, J.W.; Pezzuto, J.M.; Mehta, R.G. Deguelin suppresses the formation of carcinogen-induced aberrant crypt foci in the colon of CF-1 mice. Int. J. Cancer 2003, 104, 7–11. [Google Scholar]
- Gerhauser, C.; Mar, W.; Lee, S.K.; Suh, N.; Luo, Y.; Kosmeder, J.; Luyengi, L.; Fong, H.H.; Kinghorn, A.D.; Moriarty, R.M.; et al. Rotenoids mediate potent cancer chemopreventive activity through transcriptional regulation of ornithine decarboxylase. Nat. Med. 1995, 1, 260–266. [Google Scholar]
- Yan, Y.; Wang, Y.; Tan, Q.; Lubet, R.A.; You, M. Efficacy of deguelin and silibinin on benzo(a)pyrene-induced lung tumorigenesis in A/J mice. Neoplasia 2005, 7, 1053–1057. [Google Scholar]
- Lee, H.Y.; Suh, Y.A.; Kosmeder, J.W.; Pezzuto, J.M.; Hong, W.K.; Kurie, J.M. Deguelin-induced inhibition of cyclooxygenase-2 expression in human bronchial epithelial cells. Clin. Cancer Res. 2004, 10, 1074–1079. [Google Scholar]
- Nair, A.S.; Shishodia, S.; Ahn, K.S.; Kunnumakkara, A.B.; Sethi, G.; Aggarwal, B.B. Deguelin, an Akt inhibitor, suppresses IkappaBalpha kinase activation leading to suppression of NF-kappaB-regulated gene expression, potentiation of apoptosis, and inhibition of cellular invasion. J. Immunol. 2006, 177, 5612–5622. [Google Scholar]
- Murillo, G.; Salti, G.I.; Kosmeder, J.W.; Pezzuto, J.M.; Mehta, R.G. Deguelin inhibits the growth of colon cancer cells through the induction of apoptosis and cell cycle arrest. Eur. J. Cancer 2002, 38, 2446–2454. [Google Scholar]
- Ji, B.C.; Yu, C.C.; Yang, S.T.; Hsia, T.C.; Yang, J.S.; Lai, K.C.; Ko, Y.C.; Lin, J.J.; Lai, T.Y.; Chung, J.G. Induction of DNA damage by deguelin is mediated through reducing DNA repair genes in human non-small cell lung cancer NCI-H460 cells. Oncol. Rep. 2012, 27, 959–964. [Google Scholar]
- Toth, M.; Sohail, A.; Fridman, R. Assessment of gelatinases (MMP-2 and MMP-9) by gelatin zymography. Methods Mol. Biol. 2012, 878, 121–135. [Google Scholar]
- Bianchi-Smiraglia, A.; Paesante, S.; Bakin, A.V. Integrin beta5 contributes to the tumorigenic potential of breast cancer cells through the Src-FAK and MEK-ERK signaling pathways. Oncogene 2012, 32, 3049–3058. [Google Scholar]
- Shen, J.; Xu, L.; Owonikoko, T.K.; Sun, S.Y.; Khuri, F.R.; Curran, W.J.; Deng, X. NNK promotes migration and invasion of lung cancer cells through activation of c-Src/PKCiota/FAK loop. Cancer Lett. 2012, 318, 106–113. [Google Scholar]
- Oktay, M.; Wary, K.K.; Dans, M.; Birge, R.B.; Giancotti, F.G. Integrin-mediated activation of focal adhesion kinase is required for signaling to Jun NH2-terminal kinase and progression through the G1 phase of the cell cycle. J. Cell Biol. 1999, 145, 1461–1469. [Google Scholar]
- Kang, H.W.; Kim, J.M.; Cha, M.Y.; Jung, H.C.; Song, I.S.; Kim, J.S. Deguelin, an Akt inhibitor, down-regulates NF-kappaB signaling and induces apoptosis in colon cancer cells and inhibits tumor growth in mice. Dig. Dis. Sci. 2012, 57, 2873–2882. [Google Scholar]
- Li, Q.; Verma, I.M. NF-kappaB regulation in the immune system. Nat. Rev. Immunol. 2002, 2, 725–734. [Google Scholar]
- Chang, Y.W.; Marlin, J.W.; Chance, T.W.; Jakobi, R. RhoA mediates cyclooxygenase-2 signaling to disrupt the formation of adherens junctions and increase cell motility. Cancer Res. 2006, 66, 11700–11708. [Google Scholar]
- Sanders, M.A.; Basson, M.D. Collagen IV regulates Caco-2 migration and ERK activation via alpha1beta1- and alpha2beta1-integrin-dependent Src kinase activation. Am. J. Physiol. Gastrointest. Liver Physiol. 2004, 286, G547–557. [Google Scholar]
- Wiedmann, M.W.; Mossner, J. Molecular targeted therapy of hepatocellular carcinoma—Results of the first clinical studies. Curr. Cancer Drug Targets 2011, 11, 714–733. [Google Scholar]
- Mayoral, R.; Fernandez-Martinez, A.; Bosca, L.; Martin-Sanz, P. Prostaglandin E2 promotes migration and adhesion in hepatocellular carcinoma cells. Carcinogenesis 2005, 26, 753–761. [Google Scholar]
- Boreddy, S.R.; Srivastava, S.K. Deguelin suppresses pancreatic tumor growth and metastasis by inhibiting epithelial-to-mesenchymal transition in an orthotopic model. Oncogene 2012, 32, 3980–3991. [Google Scholar]
- Chiu, Y.J.; Hour, M.J.; Lu, C.C.; Chung, J.G.; Kuo, S.C.; Huang, W.W.; Chen, H.J.; Jin, Y.A.; Yang, J.S. Novel quinazoline HMJ-30 induces U-2 OS human osteogenic sarcoma cell apoptosis through induction of oxidative stress and up-regulation of ATM/p53 signaling pathway. J. Orthop. Res. 2011, 29, 1448–1456. [Google Scholar]
- Huang, W.W.; Chiu, Y.J.; Fan, M.J.; Lu, H.F.; Yeh, H.F.; Li, K.H.; Chen, P.Y.; Chung, J.G.; Yang, J.S. Kaempferol induced apoptosis via endoplasmic reticulum stress and mitochondria-dependent pathway in human osteosarcoma U-2 OS cells. Mol. Nutr. Food Res. 2010, 54, 1585–1595. [Google Scholar]
- Chueh, F.S.; Chen, Y.Y.; Huang, A.C.; Ho, H.C.; Liao, C.L.; Yang, J.S.; Kuo, C.L.; Chung, J.G. Bufalin-inhibited migration and invasion in human osteosarcoma U-2 OS cells is carried out by suppression of the matrix metalloproteinase-2, ERK, and JNK signaling pathways. Environ. Toxicol. 2011, 29, 21–29. [Google Scholar]
- Chen, K.T.; Hour, M.J.; Tsai, S.C.; Chung, J.G.; Kuo, S.C.; Lu, C.C.; Chiu, Y.J.; Chuang, Y.H.; Yang, J.S. The novel synthesized 6-fluoro-(3-fluorophenyl)-4-(3-methoxyanilino)quinazoline (LJJ-10) compound exhibits anti-metastatic effects in human osteosarcoma U-2 OS cells through targeting insulin-like growth factor-I receptor. Int. J. Oncol. 2011, 39, 611–619. [Google Scholar]
- Kwon, S.H.; Park, H.Y.; Kim, J.Y.; Jeong, I.Y.; Lee, M.K.; Seo, K.I. Apoptotic action of ursolic acid isolated from Corni fructus in RC-58T/h/SA#4 primary human prostate cancer cells. Bioorg. Med. Chem. Lett. 2010, 20, 6435–6438. [Google Scholar]
- Wang, M.Y.; Zhao, F.M.; Peng, H.Y.; Lou, C.H.; Li, Y.; Ding, X.; Yu, X.Y.; Yang, G.M.; Xu, D.Q.; Jiang, L.H.; et al. Investigation on the morphological protective effect of 5-hydroxymethylfurfural extracted from wine-processed Fructus corni on human L02 hepatocytes. J. Ethnopharmacol. 2010, 130, 424–428. [Google Scholar]
- Liao, C.L.; Lai, K.C.; Huang, A.C.; Yang, J.S.; Lin, J.J.; Wu, S.H.; Gibson Wood, W.; Lin, J.G.; Chung, J.G. Gallic acid inhibits migration and invasion in human osteosarcoma U-2 OS cells through suppressing the matrix metalloproteinase-2/-9, protein kinase B (PKB) and PKC signaling pathways. Food Chem. Toxicol. 2012, 50, 1734–1740. [Google Scholar]
- Pilcher, B.K.; Dumin, J.A.; Sudbeck, B.D.; Krane, S.M.; Welgus, H.G.; Parks, W.C. The activity of collagenase-1 is required for keratinocyte migration on a type I collagen matrix. J. Cell Biol. 1997, 137, 1445–1457. [Google Scholar]
- Fishman, D.A.; Liu, Y.; Ellerbroek, S.M.; Stack, M.S. Lysophosphatidic acid promotes matrix metalloproteinase (MMP) activation and MMP-dependent invasion in ovarian cancer cells. Cancer Res. 2001, 61, 3194–3199. [Google Scholar]
- Huang, Y.T.; Hwang, J.J.; Lee, L.T.; Liebow, C.; Lee, P.P.; Ke, F.C.; Lo, T.B.; Schally, A.V.; Lee, M.T. Inhibitory effects of a luteinizing hormone-releasing hormone agonist on basal and epidermal growth factor-induced cell proliferation and metastasis-associated properties in human epidermoid carcinoma A431 cells. Int. J. Cancer 2002, 99, 505–513. [Google Scholar]
- Hsu, S.C.; Kuo, C.L.; Lin, J.P.; Lee, J.H.; Lin, C.C.; Su, C.C.; Yang, M.D.; Chung, J.G. Crude extracts of Euchresta formosana radix inhibit invasion and migration of human hepatocellular carcinoma cells. Anticancer Res. 2007, 27, 2377–2384. [Google Scholar]
- Yang, Y.L.; Ji, C.; Bi, Z.G.; Lu, C.C.; Wang, R.; Gu, B.; Cheng, L. Deguelin induces both apoptosis and autophagy in cultured head and neck squamous cell carcinoma cells. PLoS One 2013, 8, e54736. [Google Scholar]
- Mehta, R.; Katta, H.; Alimirah, F.; Patel, R.; Murillo, G.; Peng, X.; Muzzio, M.; Mehta, R.G. Deguelin action involves c-Met and EGFR signaling pathways in triple negative breast cancer cells. PLoS One 2013, 8, e65113. [Google Scholar]
- Sample Availability: Samples of the deguelin are not available.
© 2014 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license ( http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Shang, H.-S.; Chang, J.-B.; Lin, J.-H.; Lin, J.-P.; Hsu, S.-C.; Liu, C.-M.; Liu, J.-Y.; Wu, P.-P.; Lu, H.-F.; Au, M.-K.; et al. Deguelin Inhibits the Migration and Invasion of U-2 OS Human Osteosarcoma Cells via the Inhibition of Matrix Metalloproteinase-2/-9 in Vitro. Molecules 2014, 19, 16588-16608. https://doi.org/10.3390/molecules191016588
Shang H-S, Chang J-B, Lin J-H, Lin J-P, Hsu S-C, Liu C-M, Liu J-Y, Wu P-P, Lu H-F, Au M-K, et al. Deguelin Inhibits the Migration and Invasion of U-2 OS Human Osteosarcoma Cells via the Inhibition of Matrix Metalloproteinase-2/-9 in Vitro. Molecules. 2014; 19(10):16588-16608. https://doi.org/10.3390/molecules191016588
Chicago/Turabian StyleShang, Hung-Sheng, Jin-Biou Chang, Ju-Hwa Lin, Jing-Pin Lin, Shu-Chun Hsu, Chi-Ming Liu, Jia-You Liu, Ping-Ping Wu, Hsu-Feng Lu, Man-Kuan Au, and et al. 2014. "Deguelin Inhibits the Migration and Invasion of U-2 OS Human Osteosarcoma Cells via the Inhibition of Matrix Metalloproteinase-2/-9 in Vitro" Molecules 19, no. 10: 16588-16608. https://doi.org/10.3390/molecules191016588