Hepatoprotective Effects of Ixora parviflora Extract against Exhaustive Exercise-Induced Oxidative Stress in Mice

Abstract
:
1. Introduction
2. Results and Discussion
2.1. Effects of IPE Supplementation on Body Weight (BW), Exhaustive Swimming Time, and Fatigue-Related Biochemical Variables

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| C | Exhaustive exercise | ||||
|---|---|---|---|---|---|
| IPE0 | IPE10 | IPE50 | IPE100 | ||
| BW (g) | 19.5 ± 0.6 | 18.2 ± 0.6 | 19.3 ± 1.5 | 18.9 ± 2.0 | 19.0 ± 1.2 |
| Swimming time (s) | --- | 693 ± 182 | 749 ± 167 | 596 ± 185 | 771 ± 463 |
| Lactate (mmol/L) | 7.5 ± 3.6 | 9.5 ± 6.2 | 10.6 ± 2.8 | 9.2 ± 2.8 | 7.6 ± 2.5 |
| NH3 (mmol/L) | 352 ± 186 * | 732 ± 249 | 595 ± 336 | 373 ± 91 * | 251 ± 125 * |
| Glucose (mg/dL) | 159 ± 55 | 142 ± 43 | 178 ± 44 | 207 ± 35 * | 207 ± 55 * |
| Creatine kinase (U/L) | 3349 ± 1403 * | 10,940 ± 5464 | 8836 ± 3254 | 5878 ± 2294 * | 5267 ± 1785 * |
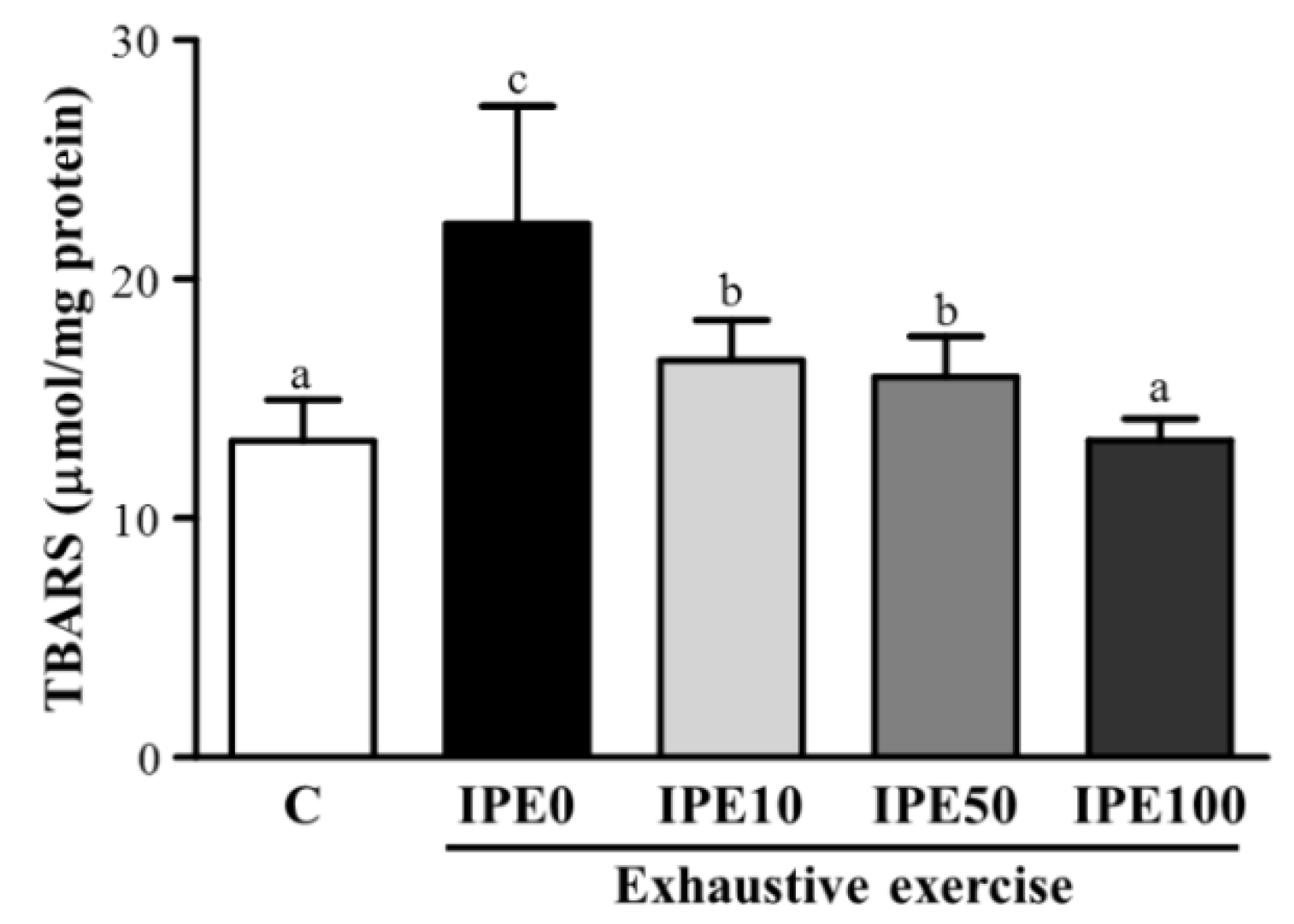
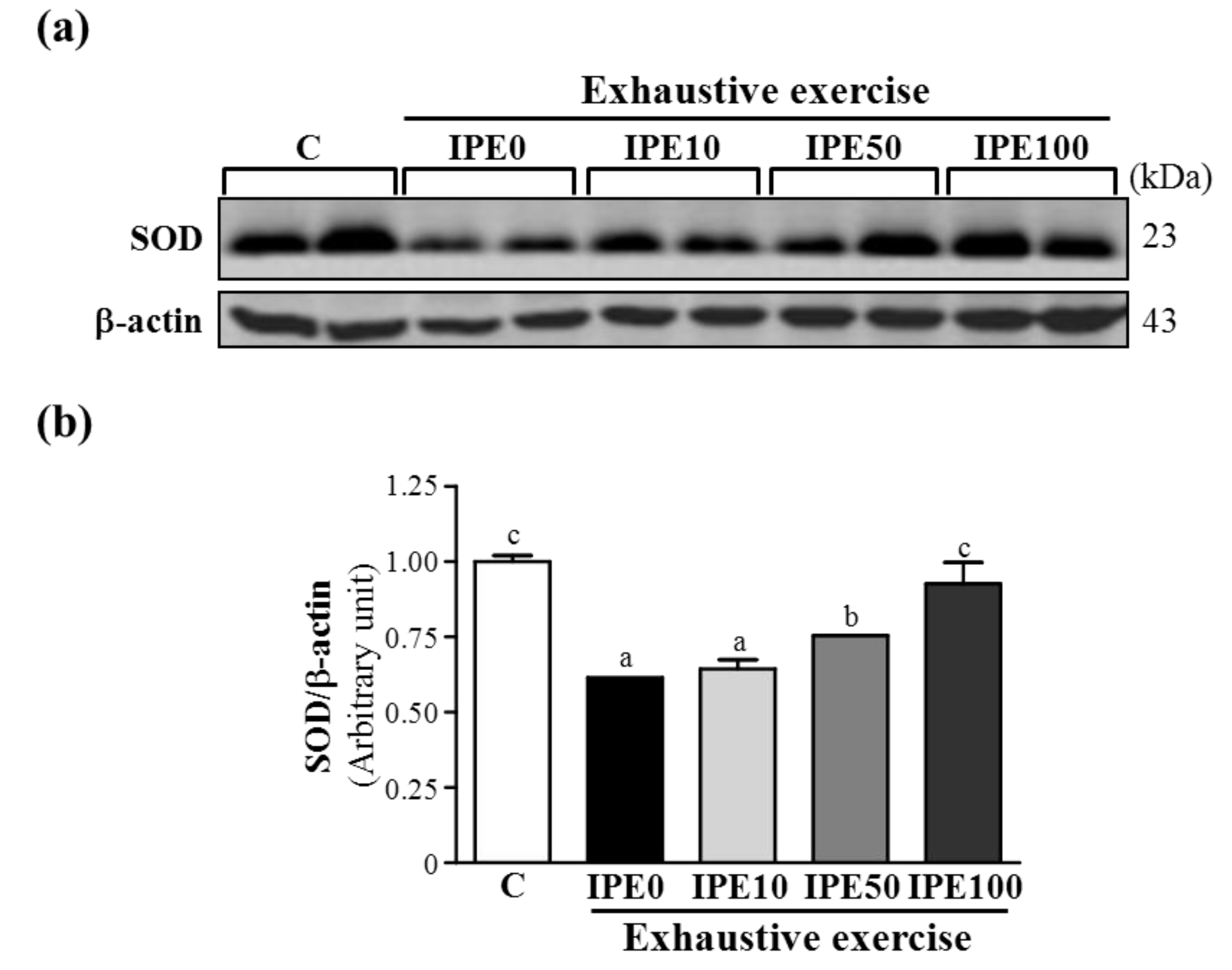
2.2. Effects of IPE Supplementation on Oxidative Stress after an Exhaustive Swimming Exercise Challenge
| Measurement | C | Exhaustive exercise | |||
|---|---|---|---|---|---|
| IPE0 | IPE10 | IPE50 | IPE100 | ||
| GPX (U/g protein) | 164 ± 27 | 148 ± 45 | 162 ± 29 | 179 ± 54 | 225 ± 84 * |
| GRD (U/g protein) | 3.3 ± 0.4 | 3.0 ± 0.4 | 3.5 ± 0.4 | 3.5 ± 0.7 | 3.9 ± 0.7 * |
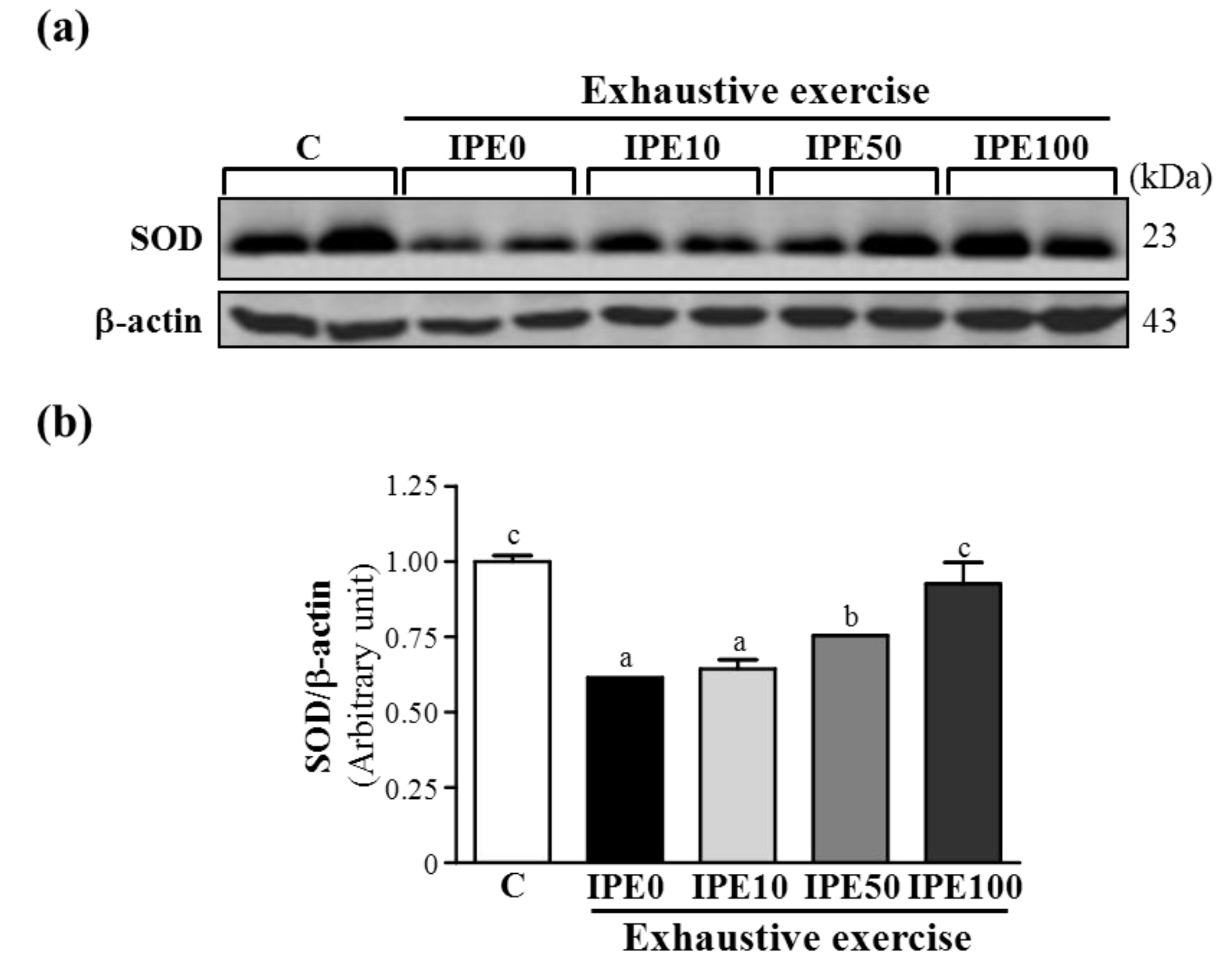
| SOD (U/g protein) | 13.7 ± 2.0 | 12.2 ± 2.3 | 14.2 ± 2.5 | 13.2 ± 2.4 | 15.8 ± 2.8 * |

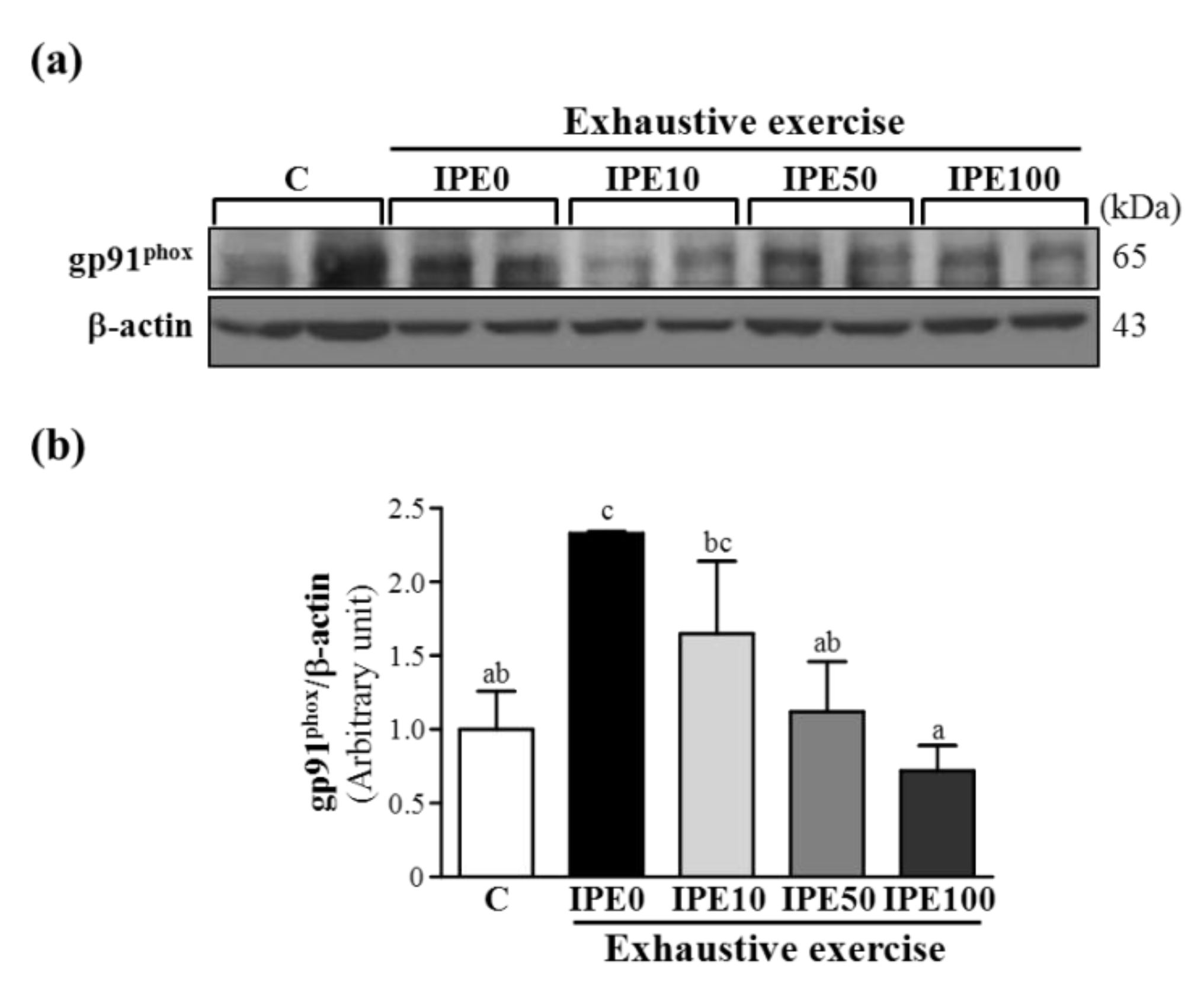
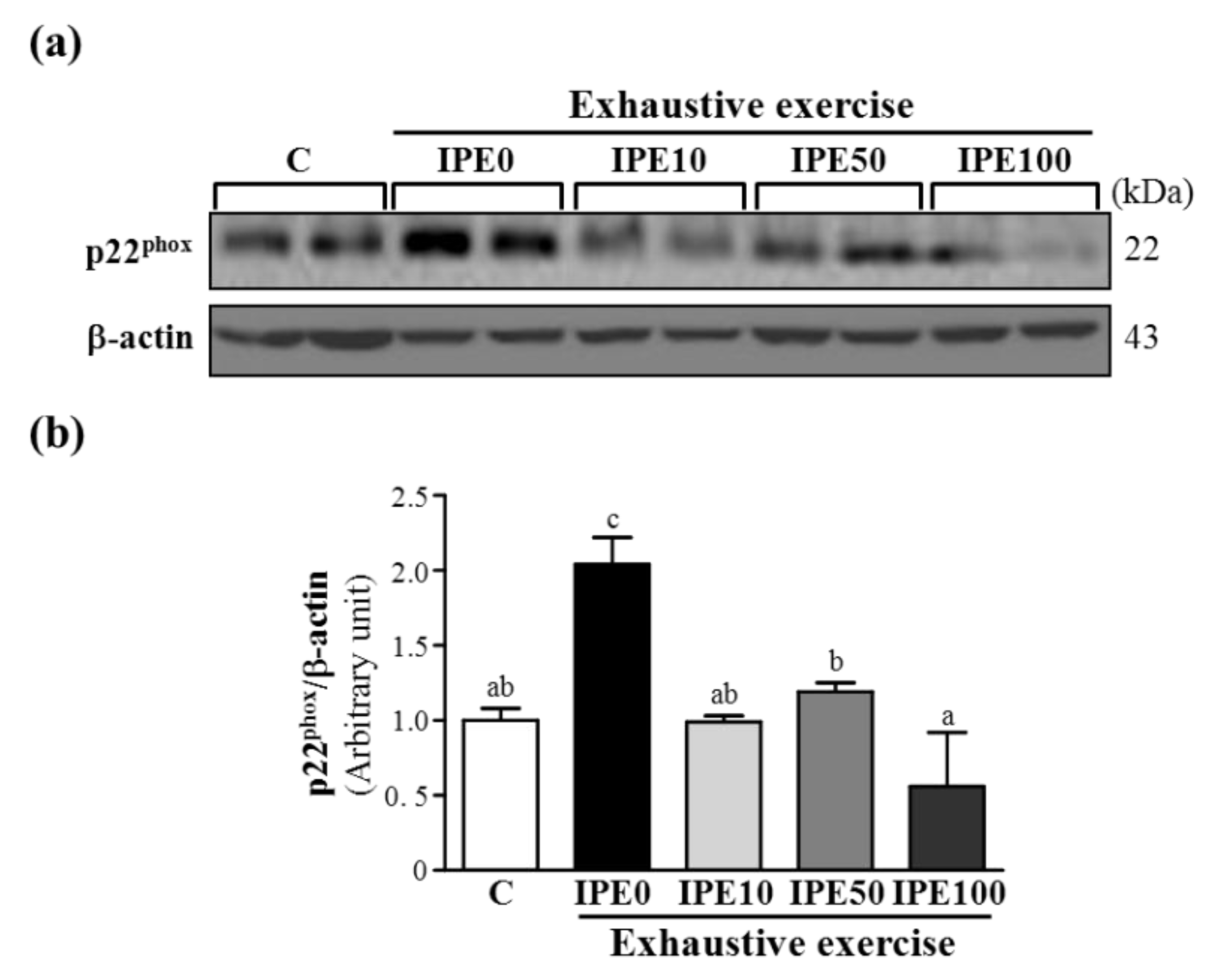
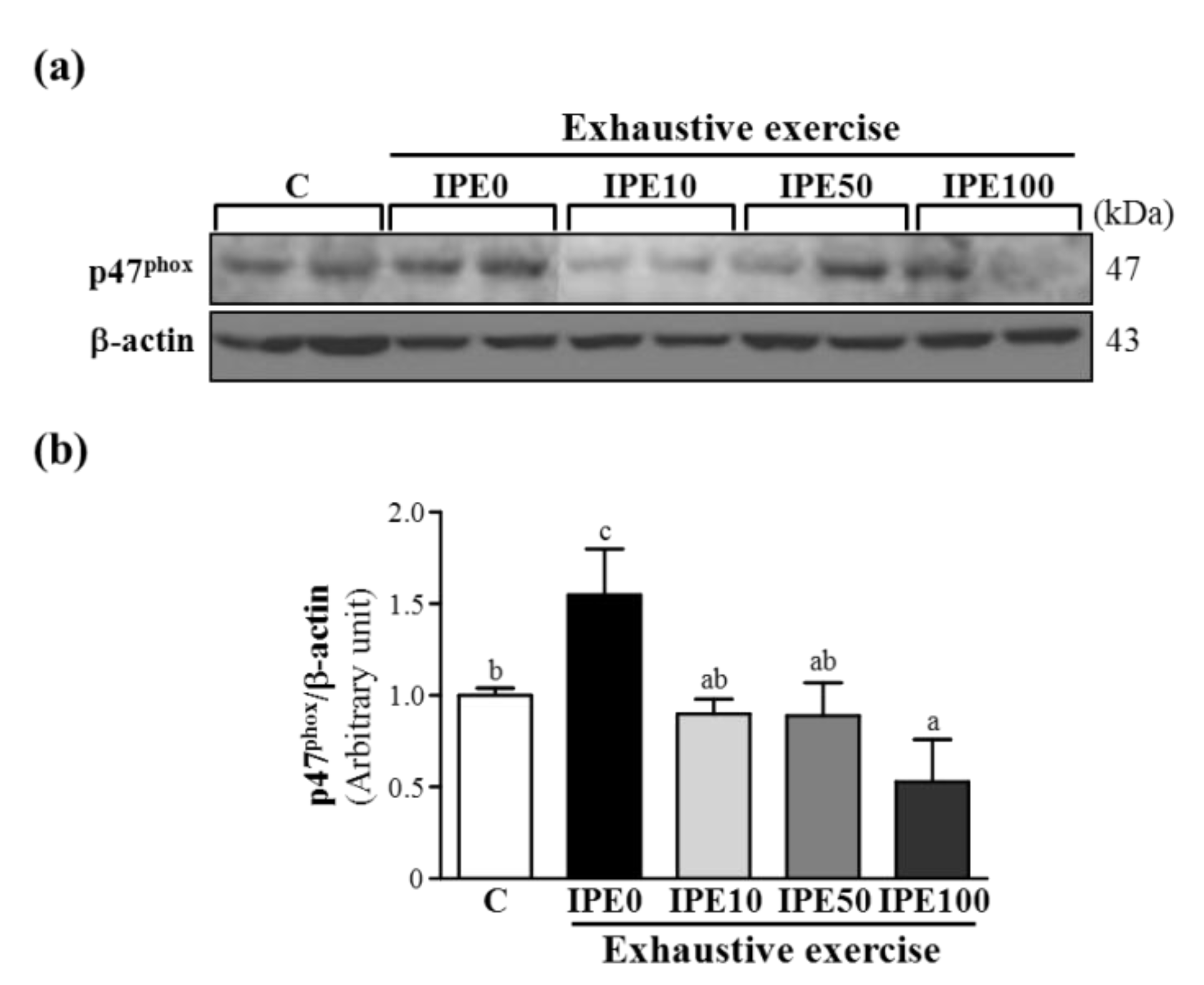
2.3. Effects of IPE Supplementation on the gp91phox, p22phox, and p47phox subunits of Nicotinamide Adenine Dinucleotide Phosphate (NADPH) Oxidase Complex Expression in Liver Tissue
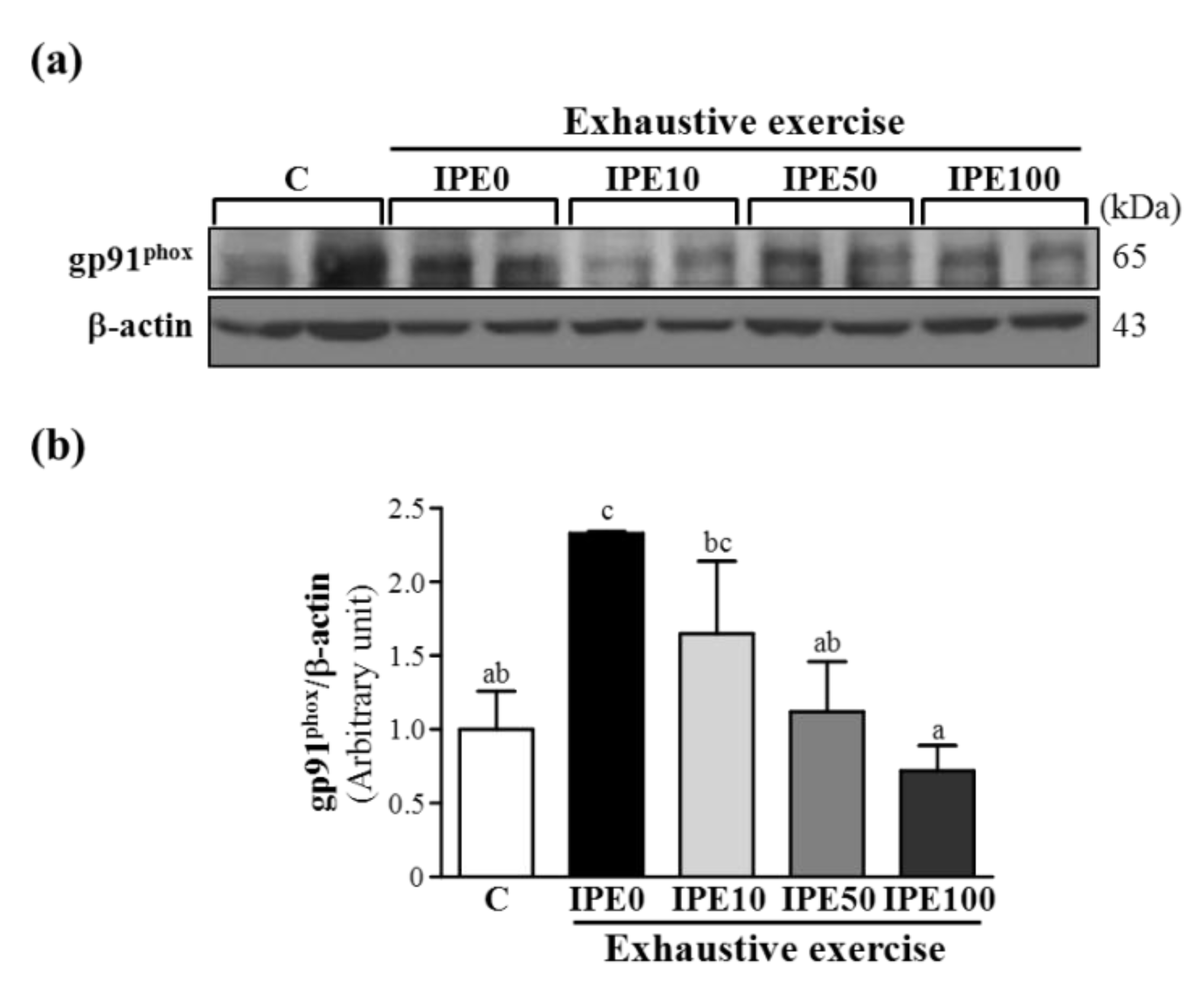
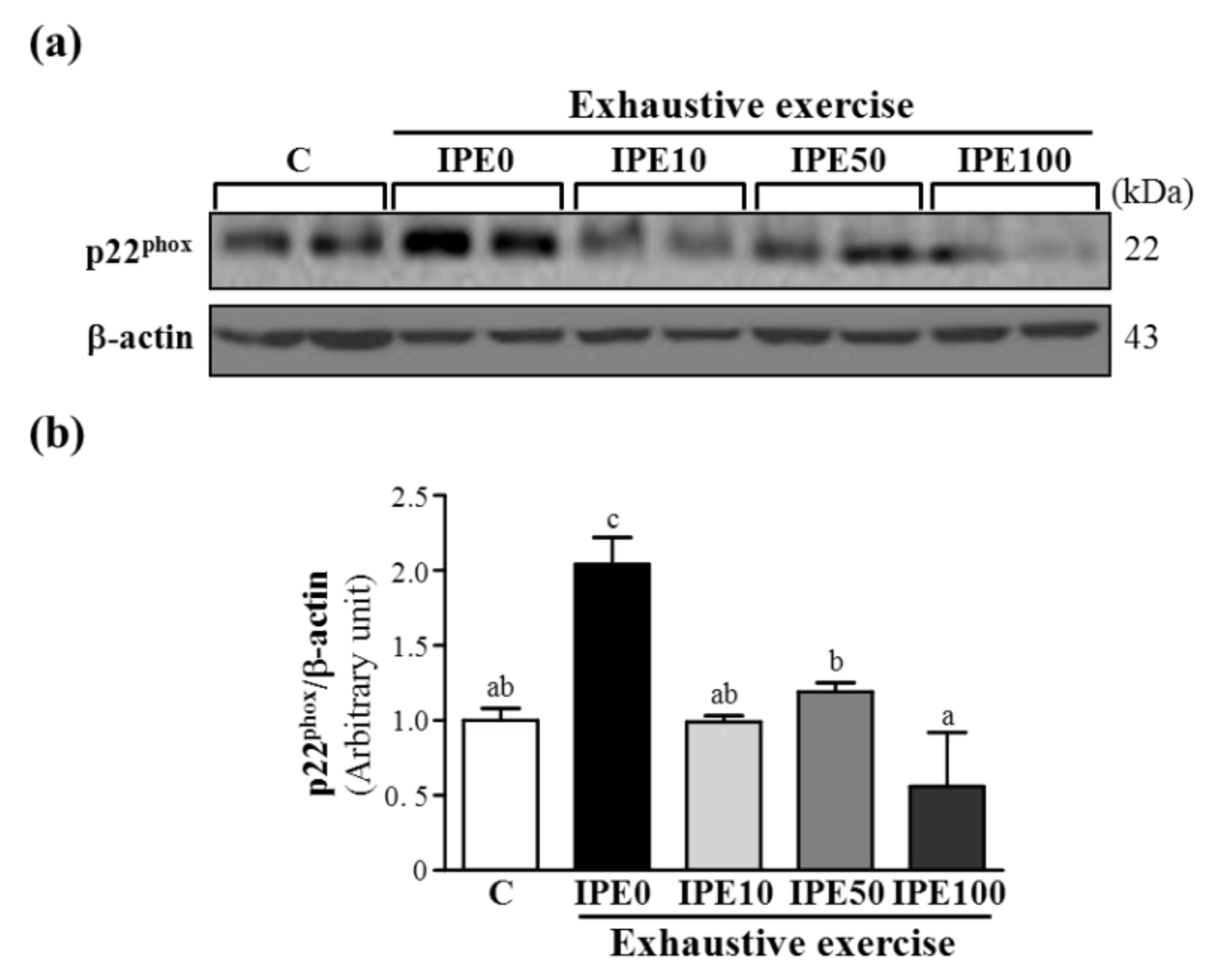

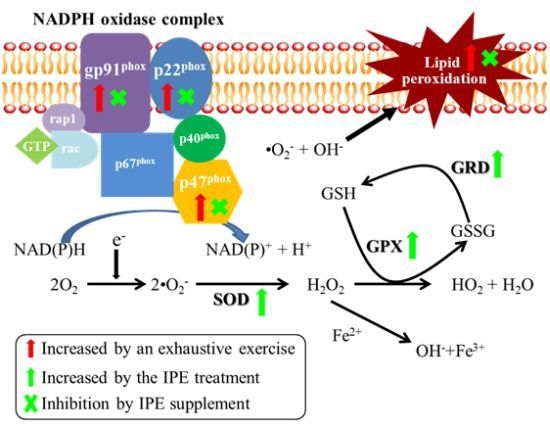
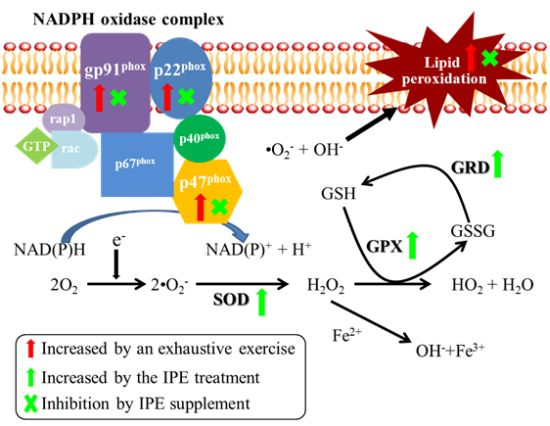
2.4. Proposed Mechanisms by Which IPE Acts against Oxidative Stress Due to Exhaustive Exercise
 ) and green arrows (
) and green arrows (  ) represent upregulation by exhaustive exercise challenge and IPE treatment, respectively. The (×) symbol represents inhibition by IPE treatment.
) and green arrows ( ) represent upregulation by exhaustive exercise challenge and IPE treatment, respectively. The (×) symbol represents inhibition by IPE treatment.
) represent upregulation by exhaustive exercise challenge and IPE treatment, respectively. The (×) symbol represents inhibition by IPE treatment.
) and green arrows ( ) represent upregulation by exhaustive exercise challenge and IPE treatment, respectively. The (×) symbol represents inhibition by IPE treatment.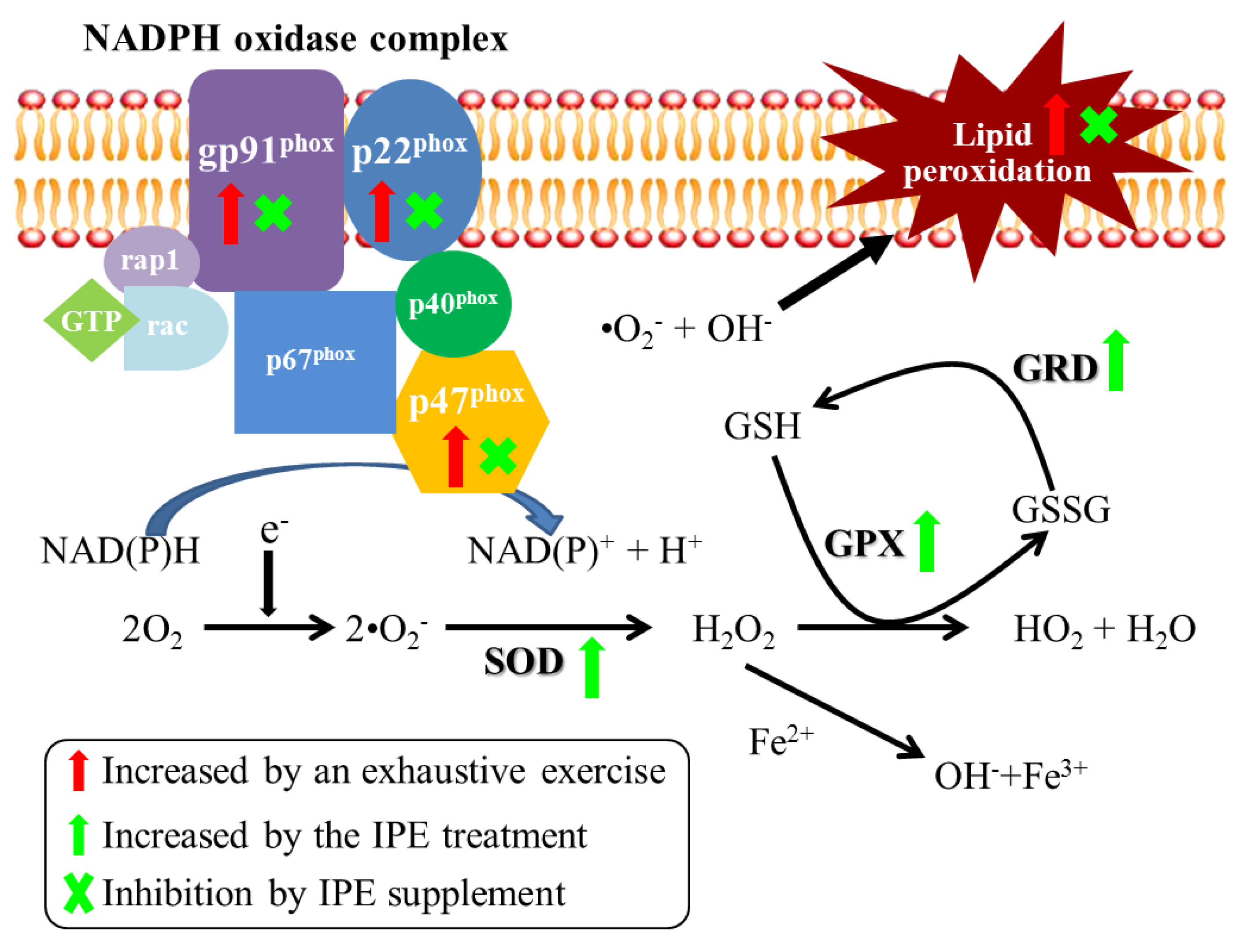
3. Experimental
3.1. Plant Material and Extraction
3.2. Animals, Treatment, and Exhaustive Swimming Exercise Challenge
3.3. Determination of Blood Biochemical Variables
3.4. Analysis of Oxidative Stress-Associated Parameters in Liver Tissues
3.5.Immunoblotting
3.6. Statistical Analysis
4. Conclusions
Acknowledgments
Conflicts of Interest
References
- Khan, K.M.; Thompson, A.M.; Blair, S.N.; Sallis, J.F.; Powell, K.E.; Bull, F.C.; Bauman, A.E. Sport and exercise as contributors to the health of nations. Lancet 2012, 380, 59–64. [Google Scholar] [CrossRef]
- Imayama, I.; Alfano, C.; Mason, C.E.; Wang, C.; Xiao, L.; Duggan, C.; Campbell, K.; Foster-Schubert, K.; Wang, C.Y.; McTiernan, A. Exercise adherence, cardiopulmonary fitness and anthropometric changes improve exercise self-efficacy and health-related quality of life. J. Phys. Act. Health. 2013, 10, 676–689. [Google Scholar]
- Davies, K.J.; Quintanilha, A.T.; Brooks, G.A.; Packer, L. Free radicals and tissue damage produced by exercise. Biochem. Biophys. Res. Commun. 1982, 107, 1198–1205. [Google Scholar] [CrossRef]
- Viña, J.; Gomez-Cabrera, M.C.; Lloret, A.; Marquez, R.; Miñana, J.B.; Pallardó, F.V.; Sastre, J. Free radicalsinexhaustivephysical exercise:Mechanismofproduction, andprotectionby antioxidants. IUBMB Life 2000, 50, 271–277. [Google Scholar] [CrossRef]
- Huang, C.C.; Lin, T.J.; Lu, Y.F.; Chen, C.C.; Huang, C.Y.; Lin, W.T. Protective effects of L-arginine supplementation against exhaustive exercise-induced oxidative stress in young rat tissues. Chin. J. Physiol. 2009, 52, 306–315. [Google Scholar] [CrossRef]
- Huang, C.C.; Lin, T.J.; Chen, C.C.; Lin, W.T. Endurance training accelerates exhaustive exercise-induced mitochondrial DNA deletion and apoptosis of left ventriclemyocardium in rats. Eur. J. Appl. Physiol. 2009, 107, 697–706. [Google Scholar] [CrossRef]
- Huang, C.C.; Huang, W.C.; Yang, S.C.; Chan, C.C.; Lin, W.T. Ganoderma tsugae Hepatoprotection against exhaustive exercise-induced liver injury in rats. Molecules 2013, 18, 1741–1754. [Google Scholar] [CrossRef]
- Baliga, M.S.; Kurian, P.J. Ixora coccinea Linn.: Traditional uses, phytochemistry and pharmacology. Chin. J. Integr. Med. 2012, 18, 72–79. [Google Scholar] [CrossRef]
- Wen, K.C.; Chiu, H.H.; Fan, P.C.; Chen, C.W.; Wu, S.M.; Chang, J.H.; Chiang, H.M. Antioxidant activity of Ixora parviflora in a cell/cell-free system and in UV-exposed human fibroblasts. Molecules 2011, 16, 5735–5752. [Google Scholar] [CrossRef]
- Wen, K.C.; Fan, P.C.; Tsai, S.Y.; Shih, I.C.; Chiang, H.M. Ixora parviflora protects against UVB-induced photoaging by inhibiting the expression of MMPs, MAP kinases, and COX-2 and by promoting type I procollagen synthesis. Evid. Based Complement. Altern. Med. 2012, 2012, 417346. [Google Scholar]
- Huang, C.C.; Hsu, M.C.; Huang, W.C.; Yang, H.R.; Hou, C.C. Triterpenoid-rich extract from Antrodia camphorata improves physical fatigue and exercise performance in mice. Evid. Based Complement Altern. Med. 2012, 2012, 364741. [Google Scholar]
- Wang, S.Y.; Huang, W.C.; Liu, C.C.; Wang, M.F.; Ho, C.S.; Huang, W.P.; Hou, C.C.; Chuang, H.L.; Huang, C.C. Pumpkin (Cucurbita moschata) fruit extract improves physical fatigue and exercise performance in mice. Molecules 2012, 17, 11864–11876. [Google Scholar] [CrossRef]
- Wu, R.E.; Huang, W.C.; Liao, C.C.; Chang, Y.K.; Kan, N.W.; Huang, C.C. Resveratrol protects against physical fatigue and improves exercise performance in mice. Molecules 2013, 18, 4689–4702. [Google Scholar] [CrossRef]
- Tashiro, S. Studies on alkaligenesis in tissues: I. Ammonia production in the nerve fiber during excitation. Am. J. Physiol. 1922, 60, 519–543. [Google Scholar]
- Carvalho-Peixoto, J.; Alves, R.C.; Cameron, L.C. Glutamine and carbohydrate supplements reduce ammonemia increase during endurance field exercise. Appl. Physiol. Nutr. Metab. 2007, 32, 1186–1190. [Google Scholar] [CrossRef]
- Suh, S.H.; Paik, I.Y.; Jacobs, K. Regulation of blood glucose homeostasis during prolonged exercise. Mol. Cells 2007, 23, 272–279. [Google Scholar]
- Warren, G.L.; Ingalls, C.P.; Lowe, D.A.; Armstrong, R.B. Excitation-contraction uncoupling: Major role in contraction-induced muscle injury. Exerc. Sport Sci. Rev. 2001, 29, 82–87. [Google Scholar] [CrossRef]
- Marin, D.P.; Bolin, A.P.; Campoio, T.R.; Guerra, B.A.; Otton, R. Oxidative stress and antioxidant status response of handball athletes: Implications for sport training monitoring. Int. Immunopharmacol. 2013, 17, 462–470. [Google Scholar] [CrossRef]
- Xu, J.; Li, Y. Effects of salidroside on exhaustive exercise-induced oxidative stress in rats. Mol. Med. Rep. 2012, 6, 1195–1198. [Google Scholar]
- Djordjevic, B.; Baralic, I.; Kotur-Stevuljevic, J.; Stefanovic, A.; Ivanisevic, J.; Radivojevic, N.; Andjelkovic, M.; Dikic, N. Effect of astaxanthin supplementation on muscle damage and oxidative stress markers in elite young soccer players. J. Sports Med. Phys. Fitness 2012, 52, 382–392. [Google Scholar]
- Lobo, V.; Patil, A.; Phatak, A.; Chandra, N. Free radicals, antioxidants and functional foods: Impact on human health. Pharmacogn. Rev. 2010, 4, 118–126. [Google Scholar] [CrossRef]
- Wang, J.S.; Huang, Y.H. Effects of exercise intensity on lymphocyte apoptosis induced by oxidative stress in men. Eur. J. Appl. Physiol. 2005, 95, 290–297. [Google Scholar] [CrossRef]
- Bedard, K.; Krause, K.H. The NOX family of ROS-generating NADPH oxidases: Physiology and pathophysiology. Physiol. Rev. 2007, 87, 245–313. [Google Scholar] [CrossRef]
- Van Beek, E.M.; Zarate, J.A.; van Bruggen, R.; Schornagel, K.; Tool, A.T.; Matozaki, T.; Kraal, G.; Roos, D.; van den Berg, T.K. SIRPα controls the activity of the phagocyte NADPH oxidase by restricting the expression of gp91(phox). Cell Rep. 2012, 2, 748–755. [Google Scholar] [CrossRef] [Green Version]
- Huang, C.C.; Lo, C.P.; Chiu, C.Y.; Shyur, L.F. Deoxyelephantopin, a novel multifunctional agent, suppresses mammary tumour growth and lung metastasis and doubles survival time in mice. Br. J. Pharmacol. 2010, 159, 856–871. [Google Scholar] [CrossRef]
- Sample Availability: A sample of the I. parviflora extract is available from the co-first author, Dr. Hsiu-Mei Chiang.
© 2013 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Kan, N.-W.; Huang, W.-C.; Lin, W.-T.; Huang, C.-Y.; Wen, K.-C.; Chiang, H.-M.; Huang, C.-C.; Hsu, M.-C. Hepatoprotective Effects of Ixora parviflora Extract against Exhaustive Exercise-Induced Oxidative Stress in Mice. Molecules 2013, 18, 10721-10732. https://doi.org/10.3390/molecules180910721
Kan N-W, Huang W-C, Lin W-T, Huang C-Y, Wen K-C, Chiang H-M, Huang C-C, Hsu M-C. Hepatoprotective Effects of Ixora parviflora Extract against Exhaustive Exercise-Induced Oxidative Stress in Mice. Molecules. 2013; 18(9):10721-10732. https://doi.org/10.3390/molecules180910721
Chicago/Turabian StyleKan, Nai-Wen, Wen-Ching Huang, Wan-Teng Lin, Chih-Yang Huang, Kuo-Ching Wen, Hsiu-Mei Chiang, Chi-Chang Huang, and Mei-Chich Hsu. 2013. "Hepatoprotective Effects of Ixora parviflora Extract against Exhaustive Exercise-Induced Oxidative Stress in Mice" Molecules 18, no. 9: 10721-10732. https://doi.org/10.3390/molecules180910721