In Vitro Action of Flavonoids in the Canine Malignant Histiocytic Cell Line DH82

Abstract
:1. Introduction
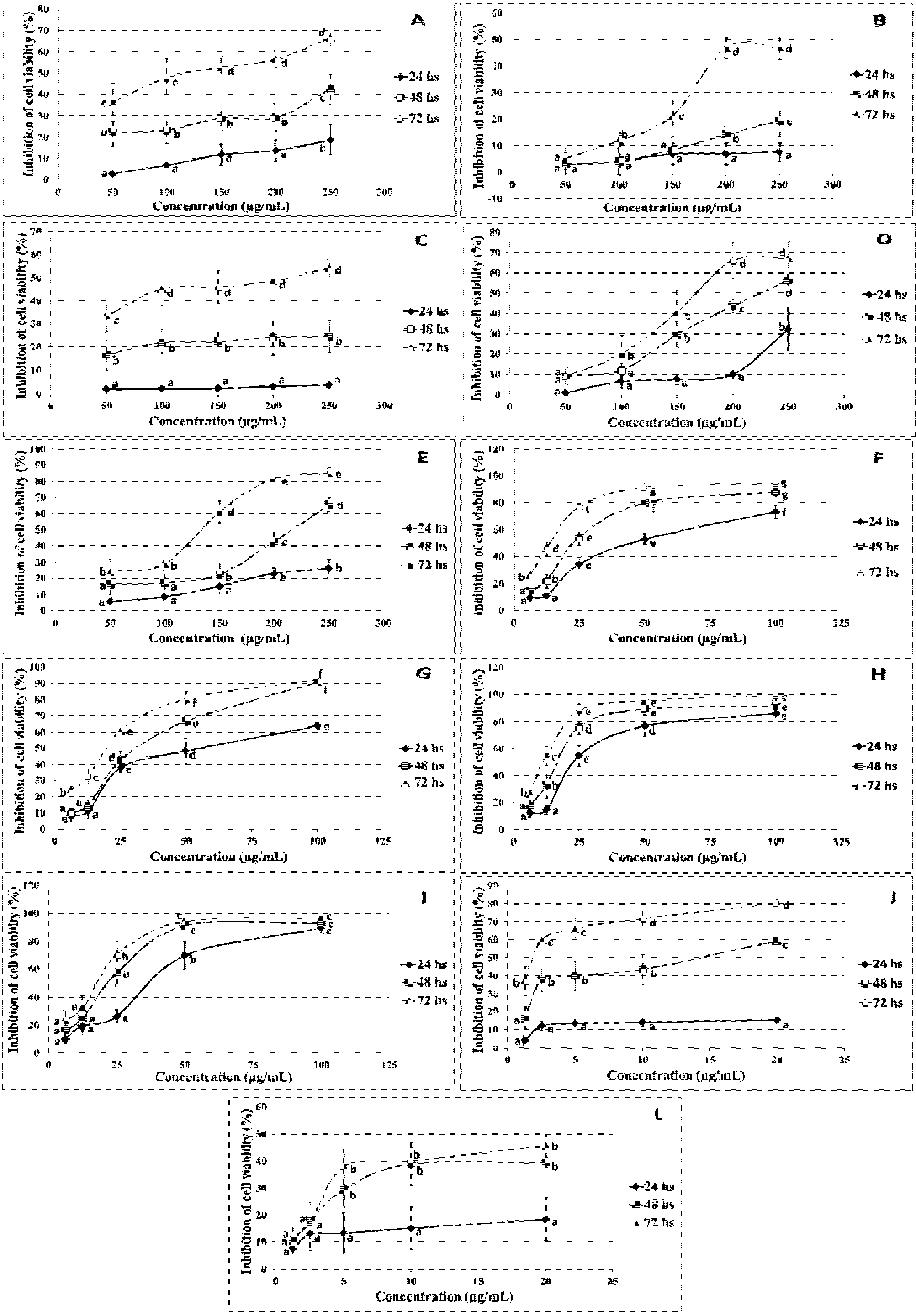
2. Results and Discussion
2.1. Cytotoxic Effect of Flavonoids

{kind=link}
{kind=link}
{kind=link}
| Flavonoids and Drugs | IC50 (µM) | ||
|---|---|---|---|
| 24 h | 48 h | 72 h | |
| Luteolin | 156.2 | 87.7 | 52.8 |
| Apigenin | 165.4 | 124.3 | 79.5 |
| 4-Methoxychalcone | 154.0 | 92.7 | 76.4 |
| trans-Chalcone | 129.2 | 89.3 | 59.5 |
| Quercetin | >992.5 | >661.7 | >302.2 |
| Genistein | >1110 | >740.1 | >370 |
| EGCG | >654.5 | >436.6 | >218.2 |
| Myricetin | >942.7 | >628.5 | >628.5 |
| Fisetin | >1048 | >1048 | >349.5 |
| Etoposide | 95.5 | 19.1 | 3.6 |
| Merbarone | 205.5 | 77.8 | 65.3 |
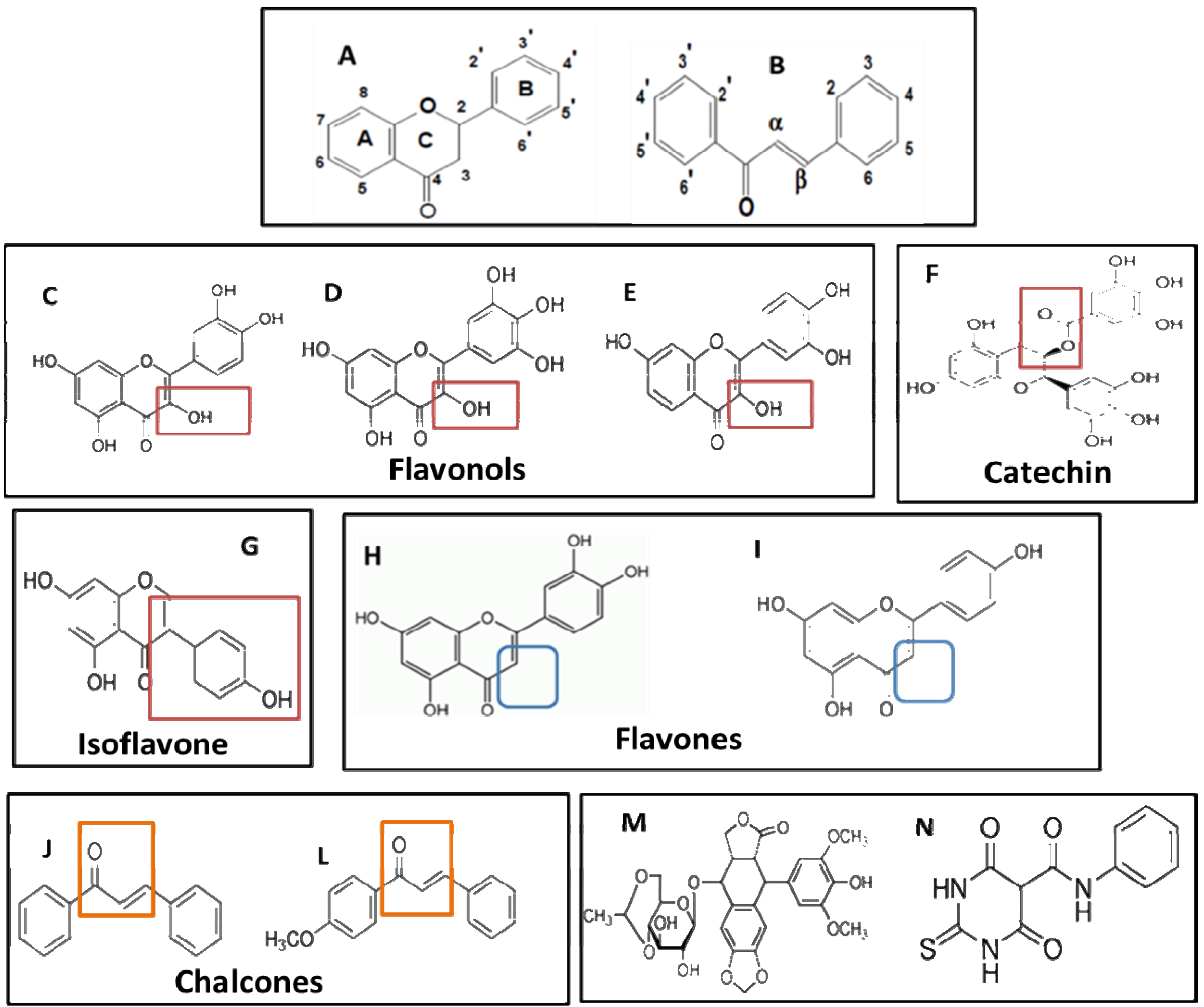
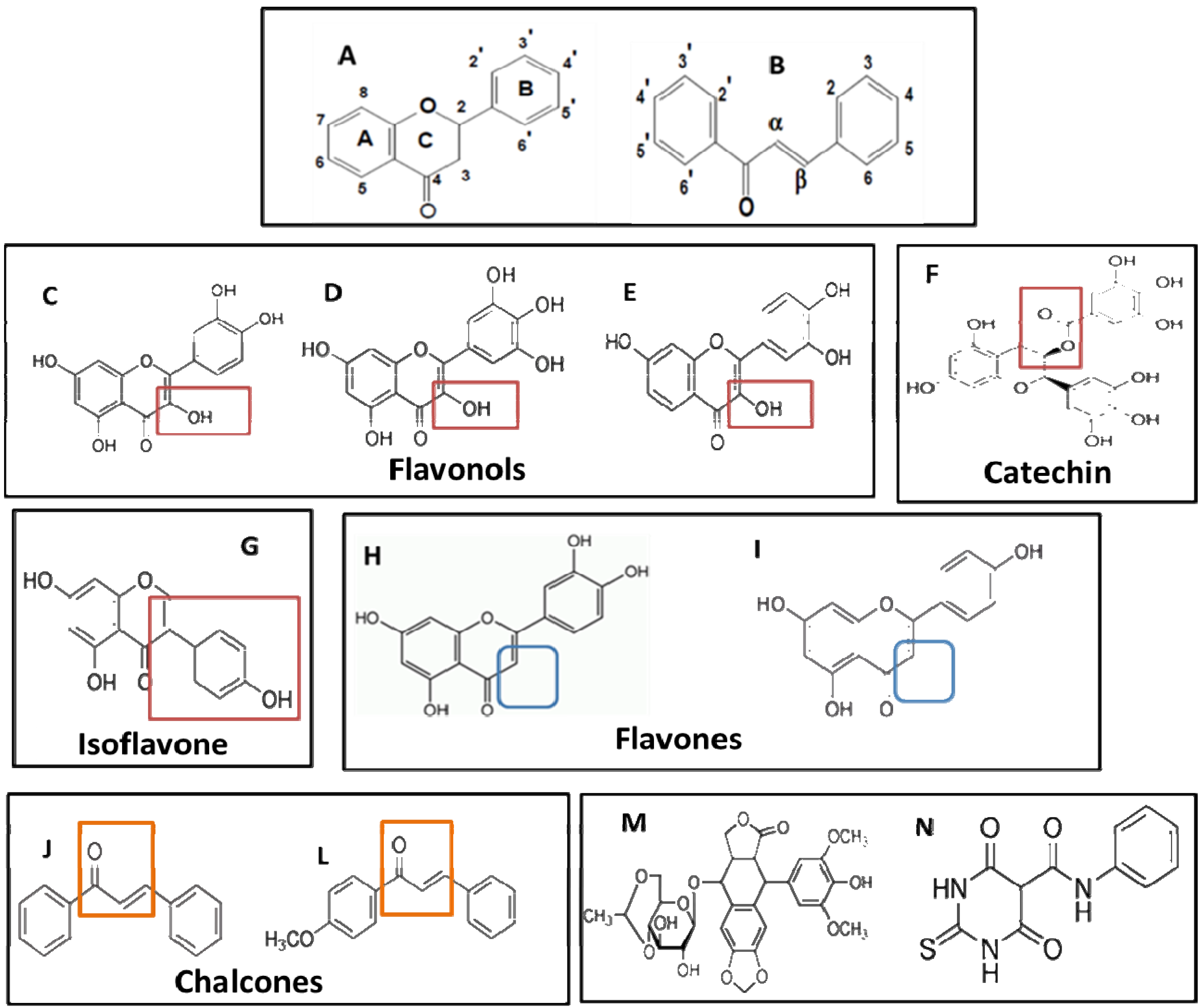
2.2. Analysis of the Relationship between Structure and Cytotoxic Effect
| Flavonoids | Class | Substituents | Total OH | Total COH3 | ||
|---|---|---|---|---|---|---|
| Chain A | Chain B | Chain C | ||||
| Cytotoxic compounds | ||||||
| trans-Chalcone | Chalcone | 0 | 0 | |||
| 4-Methoxychalcone | Chalcone | COH3-4' | 0 | 1 | ||
| Luteolin | Flavone | OH-5,7 | OH-3',4' | O-4; anel-2 | 4 | 0 |
| Apigenin | Flavone | OH-5,7 | OH-4' | O-4; anel-2 | 3 | 0 |
| Low cytotoxicity compounds | ||||||
| Quercetin | Flavonol | OH-,5,7 | OH-3',4' | O-4; OH-3; anel-2 | 5 | 0 |
| Myricetin | Flavonol | OH-,5,7 | OH-3',4',5' | O-4; OH-3; anel-2 | 6 | 0 |
| Fisetin | Flavonol | OH-7 | OH-4',5' | O-4; OH-3; anel-2 | 4 | 0 |
| EGCG | Catechin | OH-5,7 | OH-3',4',5' | galato-3; anel-2 | 8 | 0 |
| Genistein | Isoflavone | OH-5,7 | OH-4' | O-4; anel-3 | 3 | 0 |
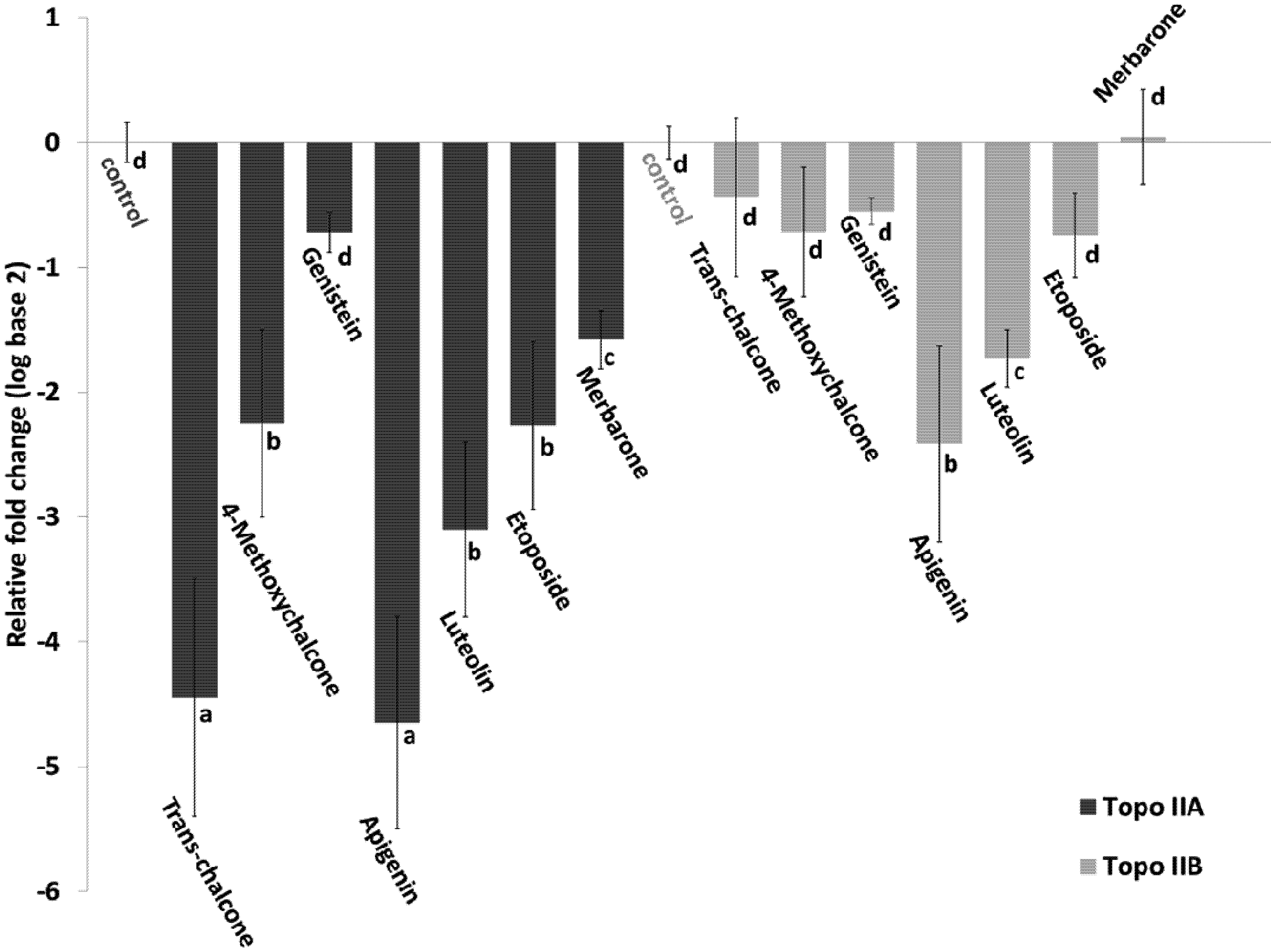
2.3. Genotoxic Effect
| Genotoxicity | |||
|---|---|---|---|
| Substances | Concentration (µg/mL) | DNA Damage Index: 0–4 | |
| 24 h | 6 h | ||
| Control (0.5% DMSO) | 0.36 ± 0.070 a | 0.4 ± 0.061 a | |
| Etoposide | 2.5 | 3.22 ± 0.065 h | 2.5 ± 0.115 f |
| Merbarone | 2.5 | 1.14 ± 0.088 d | 0.79 ± 0.052 c |
| Genistein | 200 | 2.79 ± 0.067 g | 1.84 ± 0.17 e |
| 100 | 1.80 ± 0.083 e | 1.15 ± 0.105 d | |
| 50 | 1.10 ± 0.065 d | 0.83 ± 0.095 c | |
| Quercetin | 200 | 0.65 ± 0.094 b | - |
| EGCG | 200 | 1.03 ± 0.031 d | - |
| Myricetin | 200 | 0.88 ± 0.055 c | - |
| Fisetin | 200 | 0.83 ± 0.032 c | - |
| Luteolin | 25 | 0.74 ± 0.066 b | - |
| 12.5 | 0.68 ± 0.075 b | - | |
| trans-Chalcone | 25 | 0.64 ± 0.051 b | 0.51 ± 0.079 a |
| 12.5 | 0.57 ± 0.043 a | 0.50 ± 0.039 a | |
| 4-Methoxychalcone | 25 | 0.73 ± 0.085 b | 0.48 ± 0.062 a |
| 12.5 | 0.66 ± 0.074 b | 0.47 ± 0.056 a | |
| Apigenin | 25 | 0.80 ± 0.063 | 0.66 ± 0.096 b |
| 12.5 | 0.74 ± 0.052 b | 0.57 ± 0.107 a | |
2.4. Effect of Flavonoids on the Expression of Topoisomerase II
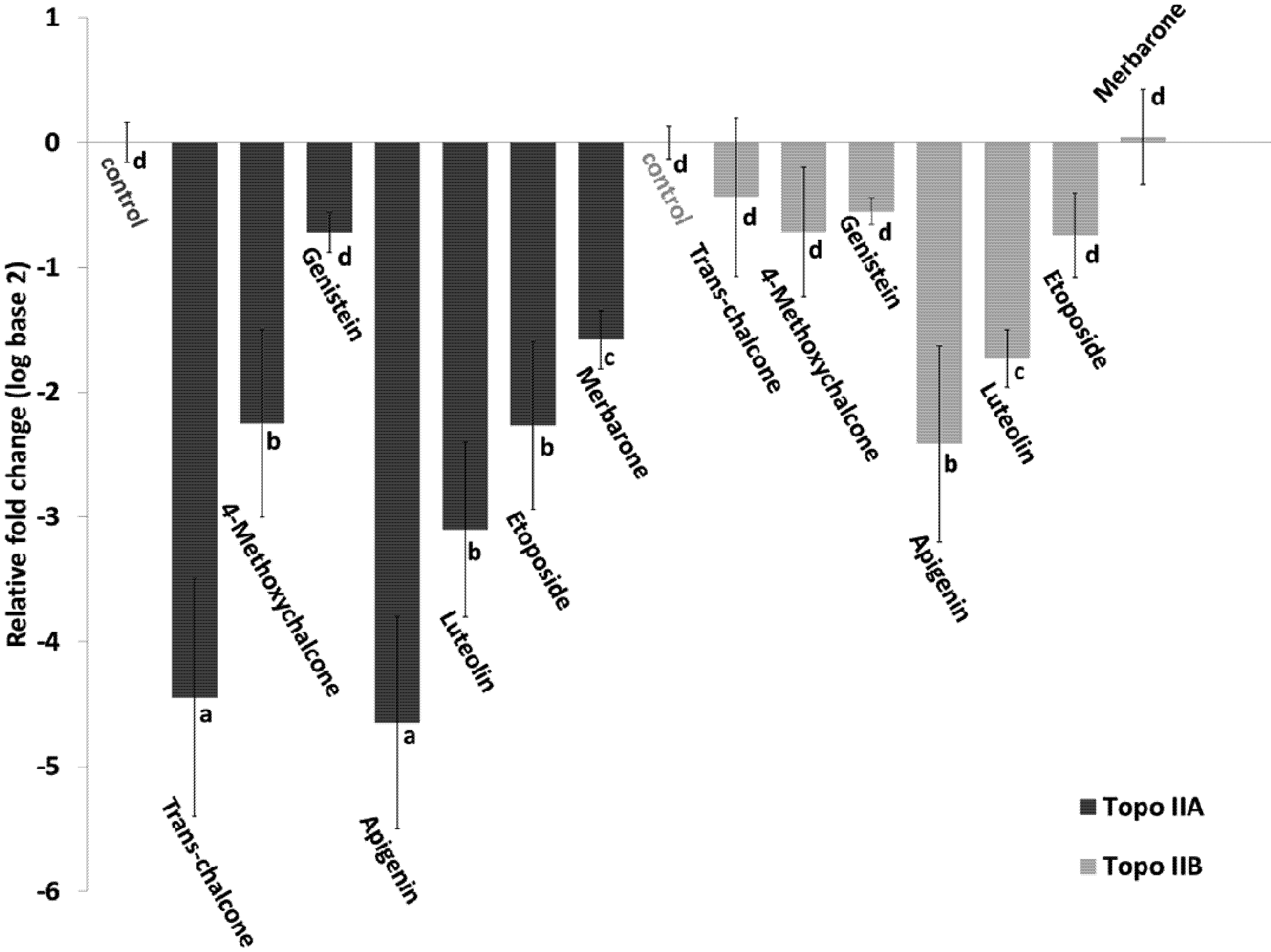
3. Experimental
3.1. Materials
3.2. Cell Culture
3.3. Cytotoxicity Assay
3.4. DNA Damage Assay
3.5. Quantification of Topoisomerase IIa and Topoisomerase IIb Messenger RNA (mRNA)
3.6. Statistical Analysis
4. Conclusions
Acknowledgments
Conflicts of Interest
References
- Baek, S.J.; McEntee, M.F.; Legendre, A.M. Review paper: Cancer chemopreventive compounds and canine cancer. Vet. Pathol. 2009, 46, 576–588. [Google Scholar] [CrossRef]
- Dobson, J.M.; Samuel, S.; Milstein, H.; Rogers, K.; Wood, J.L.N. Canine neoplasia in the UK: Estimates of incidence rates from a population of insured dogs. J. Small Anim. Pract. 2002, 43, 240–246. [Google Scholar] [CrossRef]
- Merlo, D.F.; Rossi, L.; Pellegrino, C.; Ceppi, M.; Cardellino, U.; Capurro, C.; Ratto, A.; Sambucco, P.L.; Sestito, V.; Tanara, G.; et al. Cancer incidence in pet dogs: Findings of the animal tumor registry of genoa, italy. J. Vet. Intern. Med. 2008, 22, 976–984. [Google Scholar] [CrossRef]
- Patterson, S.L.; Maresso, K.C.; Hawk, E. Cancer chemoprevention: Successes and failures. Clin. Chem. 2013, 59, 94–101. [Google Scholar] [CrossRef]
- Beecher, G.R. Overview of dietary flavonoids: Nomenclature, occurrence and intake. J. Nutr. 2003, 133, 3248S–3254S. [Google Scholar]
- De Kok, T.M.; de Waard, P.; Wilms, L.C.; van Breda, S.G. Antioxidative and antigenotoxic properties of vegetables and dietary phytochemicals: The value of genomics biomarkers in molecular epidemiology. Mol. Nutr. Food Res. 2010, 54, 208–217. [Google Scholar] [CrossRef]
- Christen, P.; Cuendet, M. Plants as a source of therapeutic and health products. Chimia 2012, 66, 320–323. [Google Scholar] [CrossRef]
- Russo, M.; Spagnuolo, C.; Tedesco, I.; Russo, G.L. Phytochemicals in cancer prevention and therapy: Truth or dare? Toxins 2010, 2, 517–551. [Google Scholar] [CrossRef]
- Procházková, D.; Boušová, I.; Wilhelmová, N. Antioxidant and prooxidant properties of flavonoids. Fitoterapia 2011, 82, 513–523. [Google Scholar] [CrossRef]
- Pietta, P.G. Flavonoids as antioxidants. J. Nat. Prod. 2000, 63, 1035–1042. [Google Scholar] [CrossRef]
- Hasebe, Y.; Egawa, K.; Yamazaki, Y.; Kunimoto, S.; Hirai, Y.; Ida, Y.; Nose, K. Specific inhibition of hypoxia-inducible factor (hif)-1 alpha activation and of vascular endothelial growth factor (vegf) production by flavonoids. Biol. Pharm. Bull. 2003, 26, 1379–1383. [Google Scholar] [CrossRef]
- Schindler, R.; Mentlein, R. Flavonoids and vitamin e reduce the release of the angiogenic peptide vascular endothelial growth factor from human tumor cells. J. Nutr. 2006, 136, 1477–1482. [Google Scholar]
- Hodnick, W.F.; Bohmont, C.W.; Capps, C.; Pardini, R.S. Inhibition of the mitochondrial nadh-oxidase (nadh-coenzyme q oxido-reductase) enzyme system by flavonoids: A structure-activity study. Biochem. Pharmacol. 1987, 36, 2873–2874. [Google Scholar] [CrossRef]
- Loizzo, M.R.; Said, A.; Tundis, R.; Rashed, K.; Statti, G.A.; Hufner, A.; Menichini, F. Inhibition of angiotensin converting enzyme (ace) by flavonoids isolated from ailanthus excelsa (roxb) (simaroubaceae). Phytother. Res. PTR 2007, 21, 32–36. [Google Scholar] [CrossRef]
- Bermudez-Soto, M.J.; Larrosa, M.; Garcia-Cantalejo, J.M.; Espin, J.C.; Tomas-Barberan, F.A.; Garcia-Conesa, M.T. Up-regulation of tumor suppressor carcinoembryonic antigen-related cell adhesion molecule 1 in human colon cancer caco-2 cells following repetitive exposure to dietary levels of a polyphenol-rich chokeberry juice. J. Nutr. Biochem. 2007, 18, 259–271. [Google Scholar] [CrossRef]
- Whitsett, T.G., Jr.; Lamartiniere, C.A. Genistein and resveratrol: Mammary cancer chemoprevention and mechanisms of action in the rat. Expert Rev. Anticancer Ther. 2006, 6, 1699–1706. [Google Scholar] [CrossRef]
- Fini, L.; Selgrad, M.; Fogliano, V.; Graziani, G.; Romano, M.; Hotchkiss, E.; Daoud, Y.A.; de Vol, E.B.; Boland, C.R.; Ricciardiello, L. Annurca apple polyphenols have potent demethylating activity and can reactivate silenced tumor suppressor genes in colorectal cancer cells. J. Nutr. 2007, 137, 2622–2628. [Google Scholar]
- Fresco, P.; Borges, F.; Marques, M.P.; Diniz, C. The anticancer properties of dietary polyphenols and its relation with apoptosis. Curr. Pharm. Des. 2010, 16, 114–134. [Google Scholar] [CrossRef]
- Azarova, A.M.; Lin, R.K.; Tsai, Y.C.; Liu, L.F.; Lin, C.P.; Lyu, Y.L. Genistein induces topoisomerase iibeta- and proteasome-mediated DNA sequence rearrangements: Implications in infant leukemia. Biochem. Biophys. Res. Commun. 2010, 399, 66–71. [Google Scholar] [CrossRef]
- Barjesteh van Waalwijk van Doorn-Khosrovani, S.; Janssen, J.; Maas, L.M.; Godschalk, R.W.; Nijhuis, J.G.; van Schooten, F.J. Dietary flavonoids induce mll translocations in primary human cd34+ cells. Carcinogenesis 2007, 28, 1703–1709. [Google Scholar] [CrossRef]
- Lin, C.N.; Chen, H.L.; Yen, M.H. Flavonoids with DNA strand-scission activity from rhus javanica var. Roxburghiana. Fitoterapia 2008, 79, 32–36. [Google Scholar] [CrossRef]
- Varela-Barca, F.N.; Agnez-Lima, L.F.; de Medeiros, S.R. Base excision repair pathway is involved in the repair of lesions generated by flavonoid-enriched fractions of pepper tree (schinus terebinthifolius, raddi) stem bark. Environ. Mol. Mutagen. 2007, 48, 672–681. [Google Scholar] [CrossRef]
- Gilbert, E.R.; Liu, D. Flavonoids influence epigenetic-modifying enzyme activity: Structure—function relationships and the therapeutic potential for cancer. Curr. Med. Chem. 2010, 17, 1756–1768. [Google Scholar] [CrossRef]
- Landis-Piwowar, K.R.; Dou, Q.P. Polyphenols: Biological activities, molecular targets, and the effect of methylation. Curr. Mol. Pharmacol. 2008, 1, 233–243. [Google Scholar]
- Li, Y.; Tollefsbol, T.O. Impact on DNA methylation in cancer prevention and therapy by bioactive dietary components. Curr. Med. Chem. 2010, 17, 2141–2151. [Google Scholar] [CrossRef]
- Hadi, S.M.; Bhat, S.H.; Azmi, A.S.; Hanif, S.; Shamim, U.; Ullah, M.F. Oxidative breakage of cellular DNA by plant polyphenols: A putative mechanism for anticancer properties. Semin. Cancer Biol. 2007, 17, 370–376. [Google Scholar] [CrossRef]
- Shamim, U.; Hanif, S.; Ullah, M.F.; Azmi, A.S.; Bhat, S.H.; Hadi, S.M. Plant polyphenols mobilize nuclear copper in human peripheral lymphocytes leading to oxidatively generated DNA breakage: Implications for an anticancer mechanism. Free Radic. Res. 2008, 42, 764–772. [Google Scholar] [CrossRef]
- Nitiss, J.L. Targeting DNA topoisomerase ii in cancer chemotherapy. Nat. Rev. Cancer 2009, 9, 338–350. [Google Scholar] [CrossRef]
- Jamadar-Shroff, V.; Papich, M.G.; Suter, S.E. Soy-derived isoflavones inhibit the growth of canine lymphoid cell lines. Clin. Cancer Res. Off. J. Am. Assoc. Cancer Res. 2009, 15, 1269–1276. [Google Scholar] [CrossRef]
- Wellman, M.; Krakowka, S.; Jacobs, R.; Kociba, G. A macrophage-monocyte cell line from a dog with malignant histiocytosis. In Vitro Cell Dev. Biol. 1988, 24, 223–229. [Google Scholar] [CrossRef]
- Fulmer, A.K.; Mauldin, G.E. Canine histiocytic neoplasia: An overview. Can. Vet. J. La Revue Vet. Can. 2007, 48, 1041–1043, 1046–1050. [Google Scholar]
- Chahar, M.K.; Sharma, N.; Dobhal, M.P.; Joshi, Y.C. Flavonoids: A versatile source of anticancer drugs. Pharmacogn. Rev. 2011, 5, 1–12. [Google Scholar] [CrossRef]
- Kobayashi, T.; Nakata, T.; Kuzumaki, T. Effect of flavonoids on cell cycle progression in prostate cancer cells. Cancer Lett. 2002, 176, 17–23. [Google Scholar] [CrossRef]
- Matsuo, M.; Sasaki, N.; Saga, K.; Kaneko, T. Cytotoxicity of flavonoids toward cultured normal human cells. Biol. Pharm. Bull. 2005, 28, 253–259. [Google Scholar] [CrossRef]
- Yadegarynia, S.; Pham, A.; Ng, A.; Nguyen, D.; Lialiutska, T.; Bortolazzo, A.; Sivryuk, V.; Bremer, M.; White, J.B. Profiling flavonoid cytotoxicity in human breast cancer cell lines: Determination of structure-function relationships. Nat. Prod. Commun. 2012, 7, 1295–1304. [Google Scholar]
- Boots, A.W.; Li, H.; Schins, R.P.; Duffin, R.; Heemskerk, J.W.; Bast, A.; Haenen, G.R. The quercetin paradox. Toxicol. Appl. Pharmacol. 2007, 222, 89–96. [Google Scholar] [CrossRef]
- Araujo, K.C.; de, M.B.C.E.M.; Pazini, F.; Valadares, M.C.; de Oliveira, V. Bioconversion of quercetin and rutin and the cytotoxicity activities of the transformed products. Food Chem. Toxicol. Int. J. Publ. Bri. Ind. Biol. Res. Assoc. 2013, 51, 93–96. [Google Scholar]
- Metodiewa, D.; Jaiswal, A.K.; Cenas, N.; Dickancaite, E.; Segura-Aguilar, J. Quercetin may act as a cytotoxic prooxidant after its metabolic activation to semiquinone and quinoidal product. Free Radic. Biol. Med. 1999, 26, 107–116. [Google Scholar] [CrossRef]
- Plochmann, K.; Korte, G.; Koutsilieri, E.; Richling, E.; Riederer, P.; Rethwilm, A.; Schreier, P.; Scheller, C. Structure-activity relationships of flavonoid-induced cytotoxicity on human leukemia cells. Arch. Biochem. Biophys. 2007, 460, 1–9. [Google Scholar] [CrossRef]
- Kawaii, S.; Tomono, Y.; Katase, E.; Ogawa, K.; Yano, M. Antiproliferative activity of flavonoids on several cancer cell lines. Biosci. Biotechnol. Biochem. 1999, 63, 896–899. [Google Scholar] [CrossRef]
- Chang, H.; Mi, M.; Ling, W.; Zhu, J.; Zhang, Q.; Wei, N.; Zhou, Y.; Tang, Y.; Yuan, J. Structurally related cytotoxic effects of flavonoids on human cancer cells in vitro. Arch. Pharmacal Res. 2008, 31, 1137–1144. [Google Scholar] [CrossRef]
- Kang, K.; Oh, S.H.; Yun, J.H.; Jho, E.H.; Kang, J.H.; Batsuren, D.; Tunsag, J.; Park, K.H.; Kim, M.; Nho, C.W. A novel topoisomerase inhibitor, daurinol, suppresses growth of hct116 cells with low hematological toxicity compared to etoposide. Neoplasia 2011, 13, 1043–1057. [Google Scholar]
- Fortune, J.M.; Osheroff, N. Merbarone inhibits the catalytic activity of human topoisomerase iialpha by blocking DNA cleavage. J. Biol. Chem. 1998, 273, 17643–17650. [Google Scholar] [CrossRef]
- Bandele, O.J.; Osheroff, N. The efficacy of topoisomerase ii-targeted anticancer agents reflects the persistence of drug-induced cleavage complexes in cells. Biochemistry 2008, 47, 11900–11908. [Google Scholar] [CrossRef]
- Bandele, O.J.; Osheroff, N. Bioflavonoids as poisons of human topoisomerase ii alpha and ii beta. Biochemistry 2007, 46, 6097–6108. [Google Scholar] [CrossRef]
- Chowdhury, A.R.; Sharma, S.; Mandal, S.; Goswami, A.; Mukhopadhyay, S.; Majumder, H.K. Luteolin, an emerging anti-cancer flavonoid, poisons eukaryotic DNA topoisomerase i. Biochem. J. 2002, 366, 653–661. [Google Scholar] [CrossRef]
- Lanoue, L.; Green, K.K.; Kwik-Uribe, C.; Keen, C.L. Dietary factors and the risk for acute infant leukemia: Evaluating the effects of cocoa-derived flavanols on DNA topoisomerase activity. Exp. Biol. Med. 2010, 235, 77–89. [Google Scholar] [CrossRef]
- Lopez-Lazaro, M.; Calderon-Montano, J.M.; Burgos-Moron, E.; Austin, C.A. Green tea constituents (−)-epigallocatechin-3-gallate (egcg) and gallic acid induce topoisomerase i- and topoisomerase ii-DNA complexes in cells mediated by pyrogallol-induced hydrogen peroxide. Mutagenesis 2011, 26, 489–498. [Google Scholar] [CrossRef]
- O’Prey, J.; Brown, J.; Fleming, J.; Harrison, P.R. Effects of dietary flavonoids on major signal transduction pathways in human epithelial cells. Biochem. Pharmacol. 2003, 66, 2075–2088. [Google Scholar] [CrossRef]
- Ramos, S. Cancer chemoprevention and chemotherapy: Dietary polyphenols and signalling pathways. Mol. Nutr. Food Res. 2008, 52, 507–526. [Google Scholar] [CrossRef]
- Zhou, N.; Yan, Y.; Li, W.; Wang, Y.; Zheng, L.; Han, S.; Yan, Y.; Li, Y. Genistein inhibition of topoisomerase iialpha expression participated by sp1 and sp3 in hela cell. Int. J. Mol. Sci. 2009, 10, 3255–3268. [Google Scholar] [CrossRef]
- Deweese, J.E.; Osheroff, N. The DNA cleavage reaction of topoisomerase ii: Wolf in sheep’s clothing. Nucleic Acids Res. 2009, 37, 738–748. [Google Scholar] [CrossRef]
- Azarova, A.M.; Lyu, Y.L.; Lin, C.P.; Tsai, Y.C.; Lau, J.Y.; Wang, J.C.; Liu, L.F. Roles of DNA topoisomerase ii isozymes in chemotherapy and secondary malignancies. Proc. Natl. Acad. Sci. USA 2007, 104, 11014–11019. [Google Scholar] [CrossRef]
- Lyu, Y.L.; Kerrigan, J.E.; Lin, C.P.; Azarova, A.M.; Tsai, Y.C.; Ban, Y.; Liu, L.F. Topoisomerase iibeta mediated DNA double-strand breaks: Implications in doxorubicin cardiotoxicity and prevention by dexrazoxane. Cancer Res. 2007, 67, 8839–8846. [Google Scholar] [CrossRef]
- Mandraju, R.; Chekuri, A.; Bhaskar, C.; Duning, K.; Kremerskothen, J.; Kondapi, A.K. Topoisomerase iibeta associates with ku70 and parp-1 during double strand break repair of DNA in neurons. Arch. Biochem. Biophys. 2011, 516, 128–137. [Google Scholar] [CrossRef]
- Mandraju, R.K.; Kannapiran, P.; Kondapi, A.K. Distinct roles of topoisomerase ii isoforms: DNA damage accelerating alpha, double strand break repair promoting beta. Arch. Biochem. Biophys. 2008, 470, 27–34. [Google Scholar] [CrossRef]
- Larsen, A.K.; Escargueil, A.E.; Skladanowski, A. Catalytic topoisomerase ii inhibitors in cancer therapy. Pharmacol. Ther. 2003, 99, 167–181. [Google Scholar] [CrossRef]
- Mosmann, T. Rapid colorimetric assay for cellular growth and survival: Application to proliferation and cytotoxicity assays. J. Immunol. Methods 1983, 65, 55–63. [Google Scholar] [CrossRef]
- McKelvey-Martin, V.J.; Green, M.H.L.; Schmezer, P.; Pool-Zobel, B.L.; de Méo, M.P.; Collins, A. The single cell gel electrophoresis assay (comet assay): A european review. Mutat. Res./Fundam. Mol. Mech. Mutagen. 1993, 288, 47–63. [Google Scholar] [CrossRef]
- Collins, A.R.; Ma, A.G.; Duthie, S.J. The kinetics of repair of oxidative DNA damage (strand breaks and oxidised pyrimidines) in human cells. Mutat. Res. 1995, 336, 69–77. [Google Scholar] [CrossRef]
- Silva, C.E.V.; Camacho, A.A. Alterações ecocardiográficas em cães sob tratamento prolongado com doxorrubicina. Arq. Bras. Med. Vet. Zootec. 2005, 57, 300–306. [Google Scholar] [CrossRef]
- Sample Availability: Samples of the compounds are available from the authors.
© 2013 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Silva, G.; Fachin, A.L.; Beleboni, R.O.; França, S.C.; Marins, M. In Vitro Action of Flavonoids in the Canine Malignant Histiocytic Cell Line DH82. Molecules 2013, 18, 15448-15463. https://doi.org/10.3390/molecules181215448
Silva G, Fachin AL, Beleboni RO, França SC, Marins M. In Vitro Action of Flavonoids in the Canine Malignant Histiocytic Cell Line DH82. Molecules. 2013; 18(12):15448-15463. https://doi.org/10.3390/molecules181215448
Chicago/Turabian StyleSilva, Gabriel, Ana Lúcia Fachin, Renê O. Beleboni, Suzelei C. França, and Mozart Marins. 2013. "In Vitro Action of Flavonoids in the Canine Malignant Histiocytic Cell Line DH82" Molecules 18, no. 12: 15448-15463. https://doi.org/10.3390/molecules181215448