Inhibitory Effect and Mechanism on Antiproliferation of Isoatriplicolide Tiglate (PCAC) from Paulownia Coreana

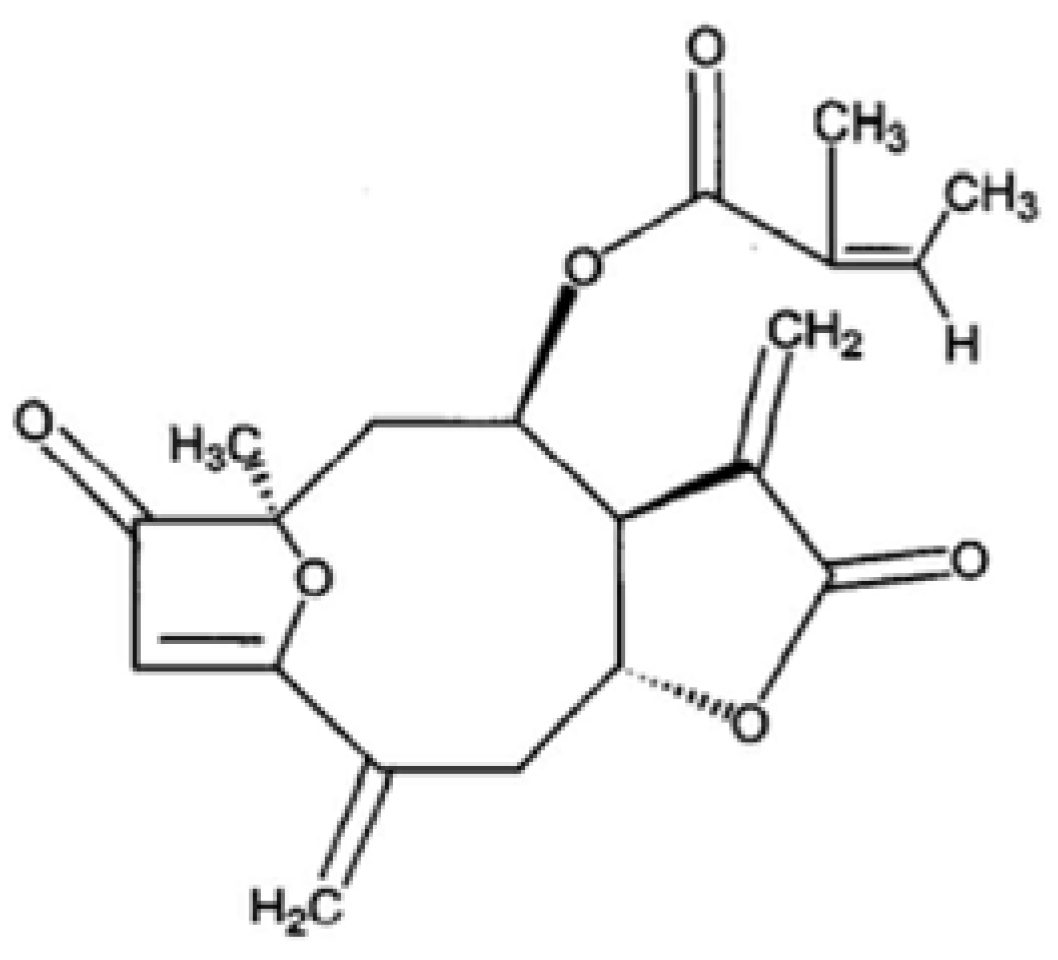{kind=link}
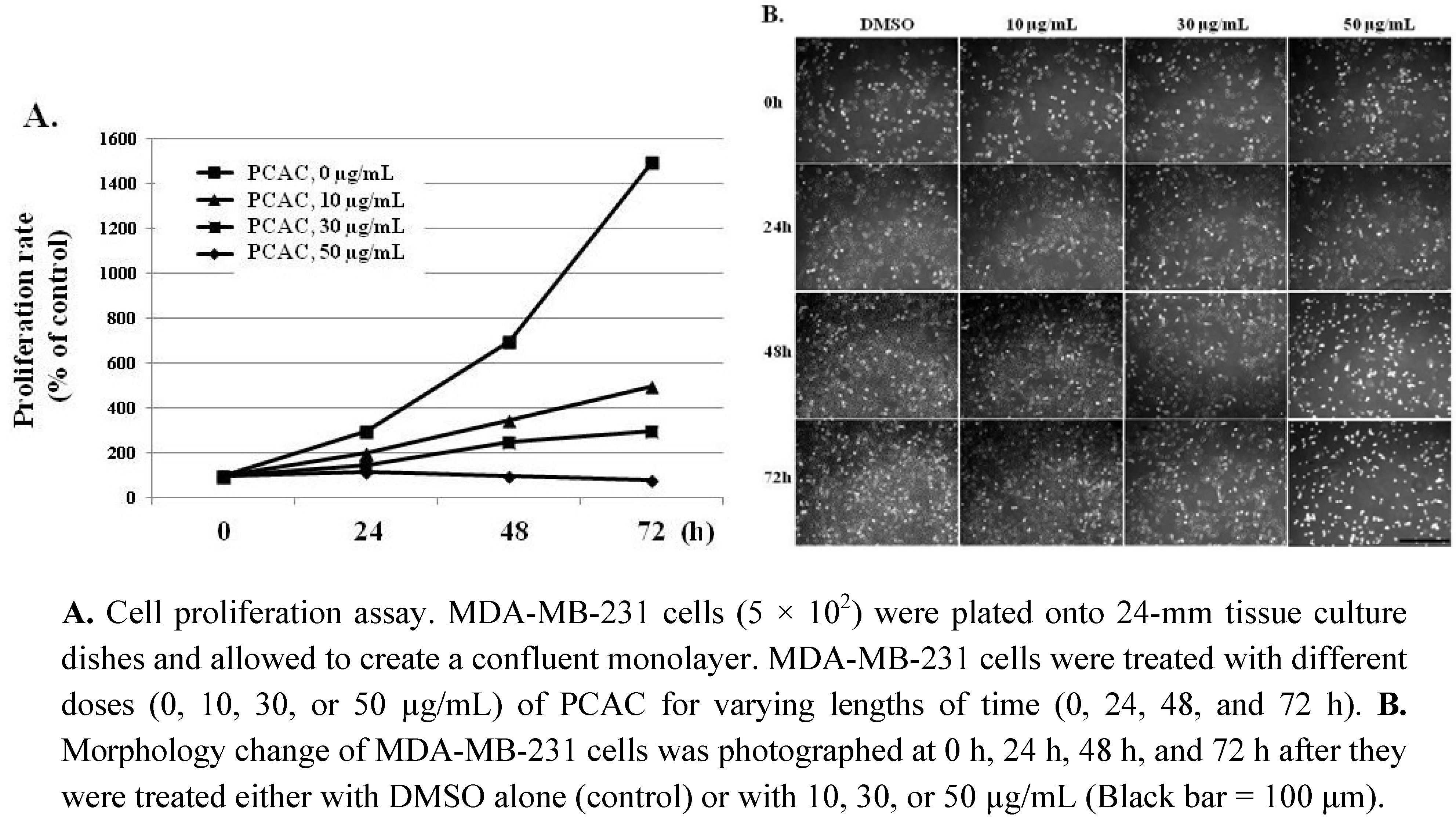{kind=link}
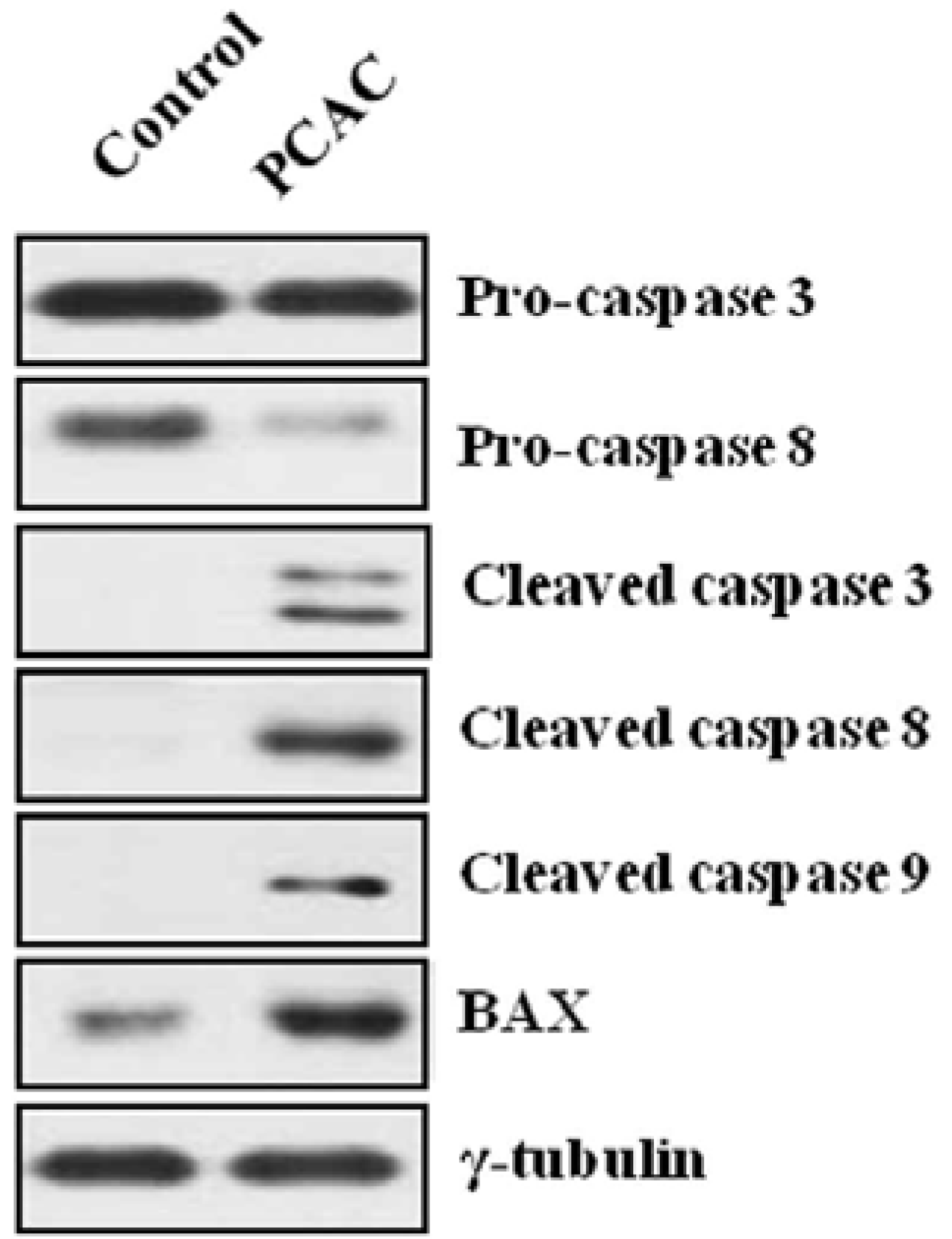{kind=link}
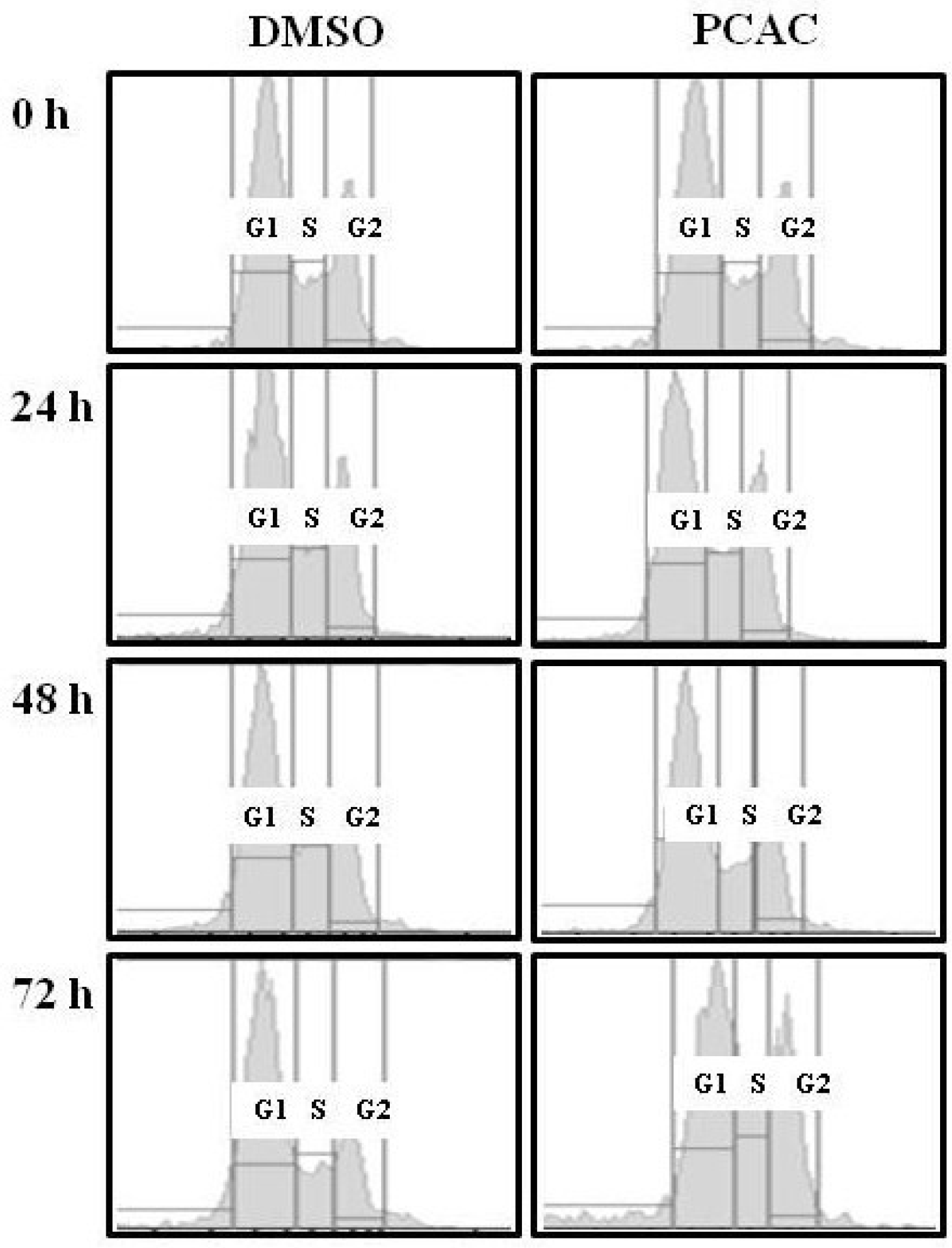{kind=link}
{kind=link}
Abstract
:1. Introduction
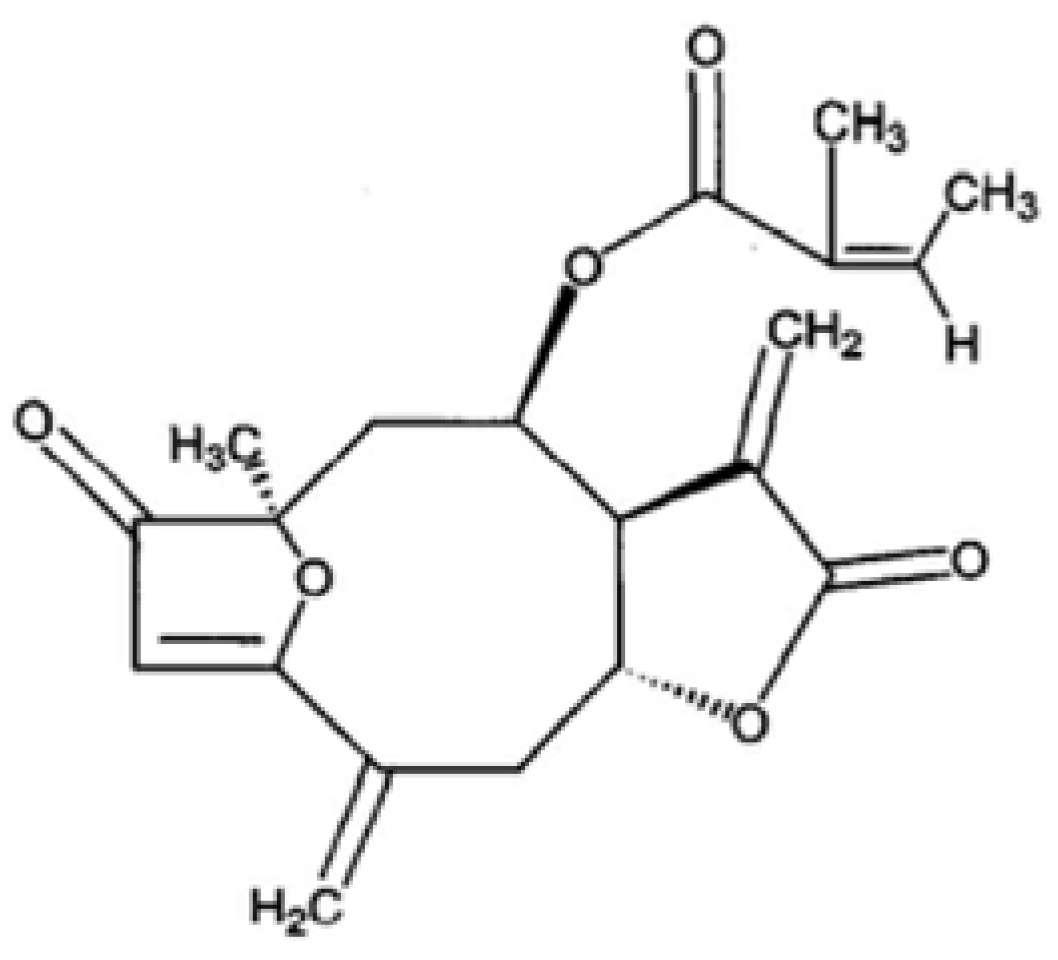
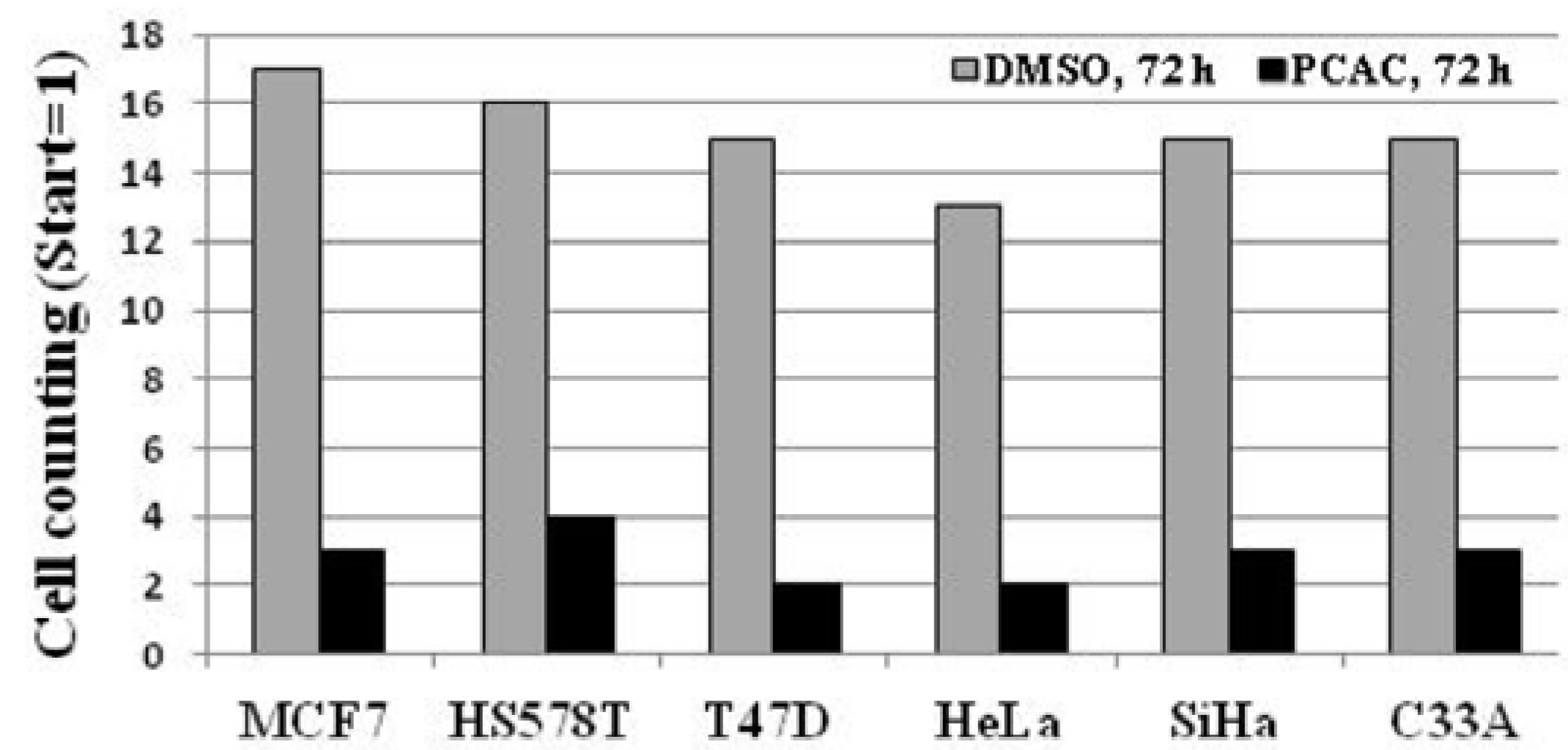
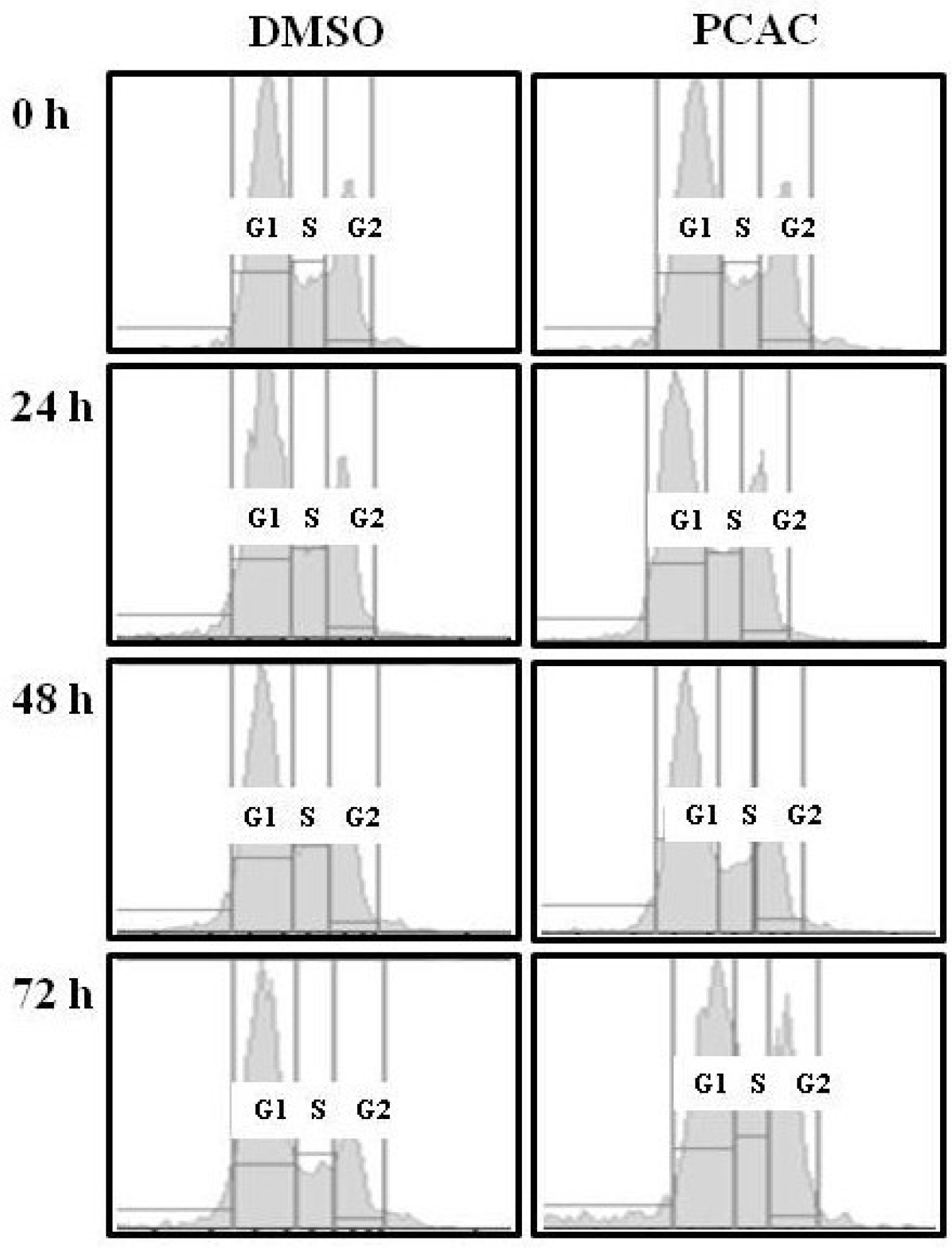
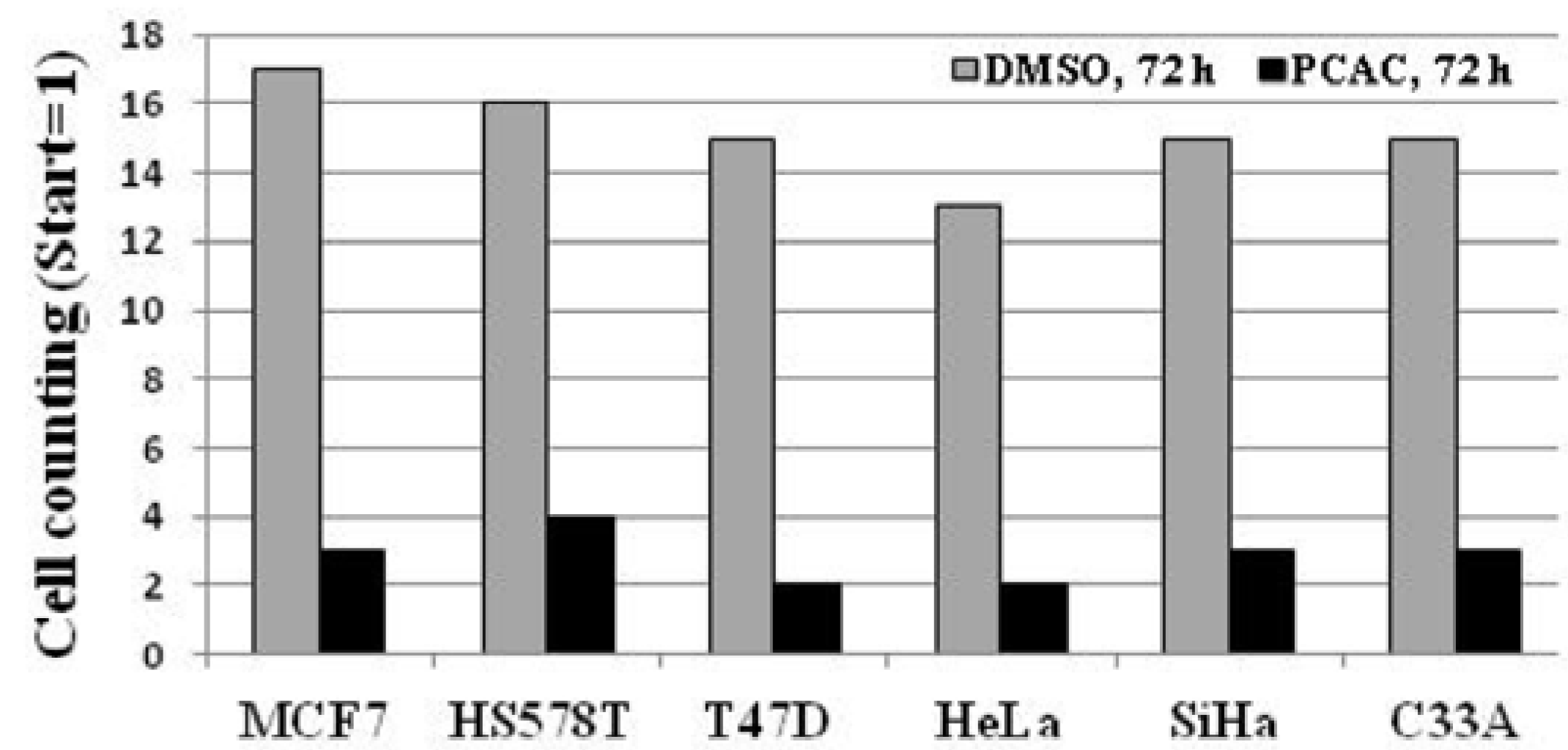
2. Results and Discussion
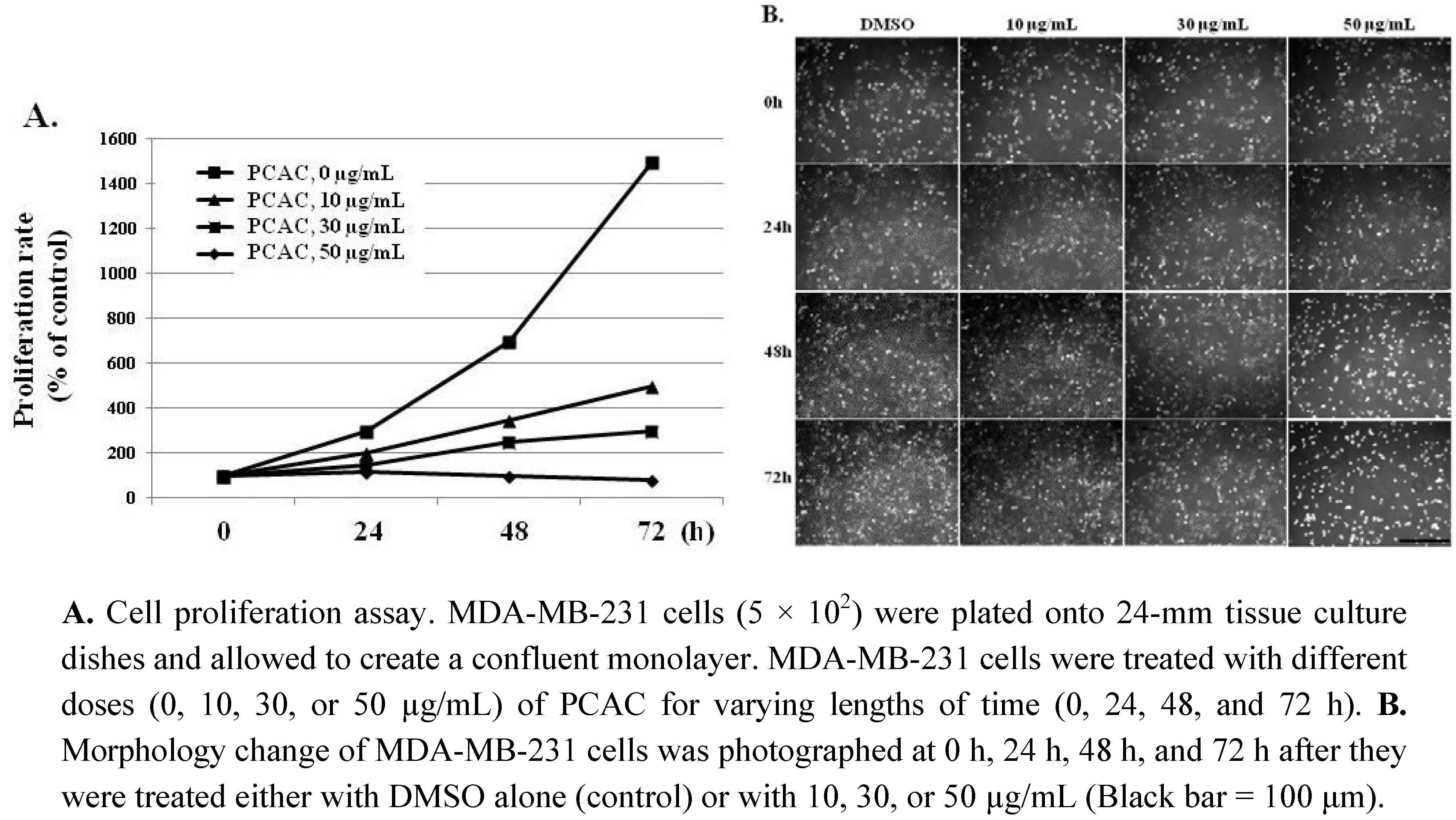
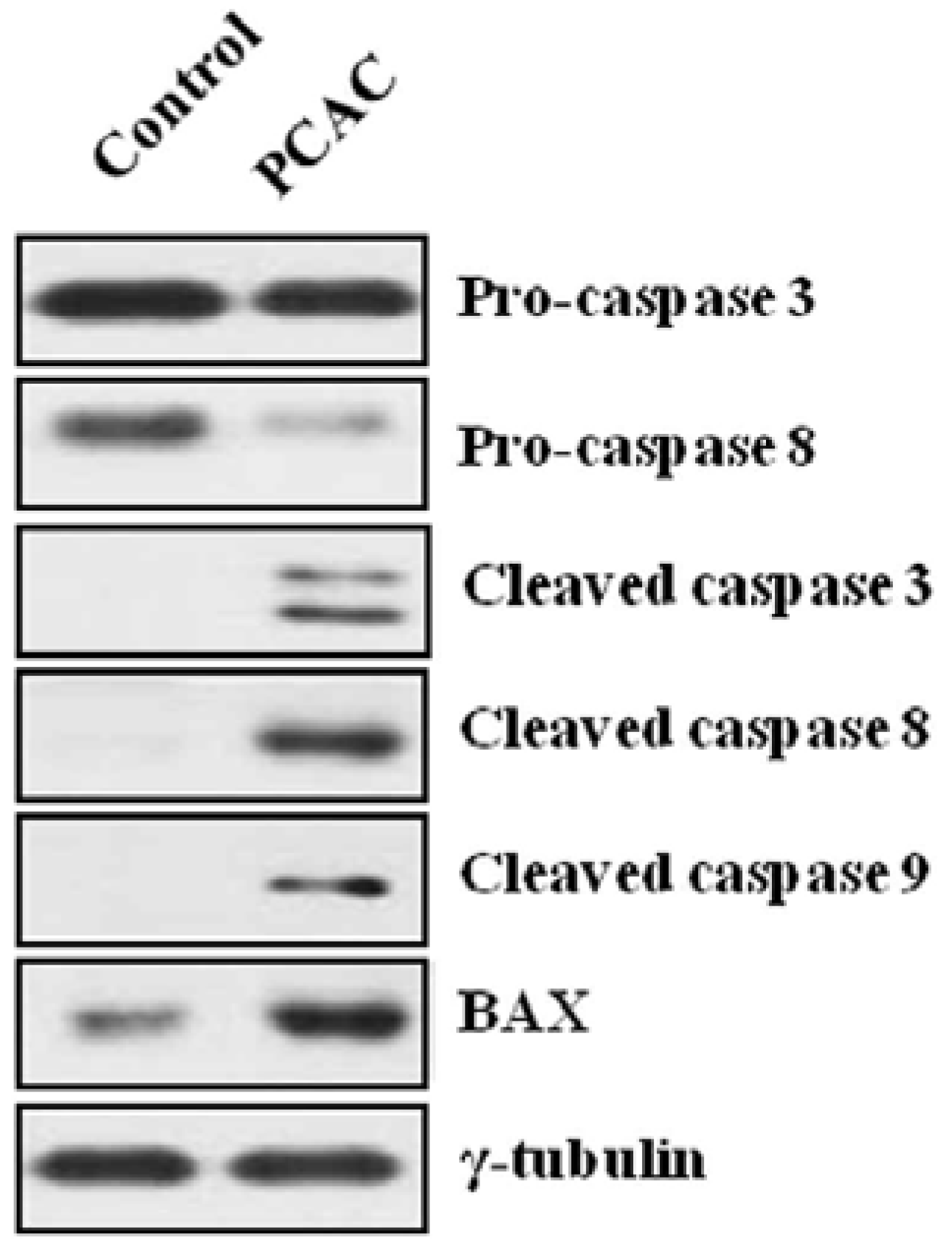
3. Experimental
4. Conclusions
Acknowledgments
Conflict of Interest
- Samples Availability: Samples of the compounds in mg scale are available from the authors.
References and Notes
- Chung, I.M.; Kim, E.H.; Jeon, H.S.; Moon, H.I. Protective effects of isoatriplicolide tiglate from Paulownia coreana against glutamate-induced neurotoxicity in primary cultured rat cortical cells. Nat. Prod. Commun. 2010, 5, 851–852. [Google Scholar]
- Steller, H. Mechanisms and Genes of Cellular Suicide. Science 1995, 267, 1445–1449. [Google Scholar]
- Jacobson, M.D.; Weil, M.; Raff, M.C. Programmed cell death in animal development. Cell 1997, 88, 347–354. [Google Scholar] [CrossRef]
- Degterev, A.; Boyce, M.; Yuan, J. A decade of caspases. Oncogene 2003, 22, 8543–8567. [Google Scholar] [CrossRef]
- Chen, M.; Wang, J. Initiator caspases in apoptosis signaling pathways. Apoptosis 2002, 7, 313–319. [Google Scholar] [CrossRef]
- Ashkenazi, A.; Dixit, V.M. Death receptors: Signaling and modulation. Science 1998, 281, 1305–1308. [Google Scholar] [CrossRef]
- Zamzami, N.; Kroemer, G. The mitochondrion in apoptosis: How Pandora’s box opens. Nat. Rev. Mol. Cell Biol. 2001, 2, 67–71. [Google Scholar] [CrossRef]
- Li, H.; Zhu, H.; Xu, C.J.; Yuan, J. Cleavage of BID by caspase 8 mediates the mitochondrial damage in the Fas pathway of apoptosis. Cell 1998, 94, 491–501. [Google Scholar] [CrossRef]
- Festjens, N.; van Gurp, M.; van Loo, G.; Saelens, X.; Vandenabeele, P. Bcl-2 family members as sentinels of cellular integrity and role of mitochondrial intermembrane space proteins in apoptotic cell death. Acta Haematol. 2004, 111, 7–27. [Google Scholar] [CrossRef]
- Luo, X.; Budihardjo, I.; Zou, H.; Slaughter, C.; Wang, X. Bid, a Bcl2 interacting protein, mediates cytochrome c release from mitochondria in response to activation of cell surface death receptors. Cell 1998, 94, 481–490. [Google Scholar] [CrossRef]
© 2012 by the authors; licensee MDPI, Basel, Switzerland. This article is an open-access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Jung, S.; Moon, H.-I.; Ohk, J.; Lee, S.; Li, C.; Kim, S.-K.; Lee, M.-S. Inhibitory Effect and Mechanism on Antiproliferation of Isoatriplicolide Tiglate (PCAC) from Paulownia Coreana. Molecules 2012, 17, 5945-5951. https://doi.org/10.3390/molecules17055945
Jung S, Moon H-I, Ohk J, Lee S, Li C, Kim S-K, Lee M-S. Inhibitory Effect and Mechanism on Antiproliferation of Isoatriplicolide Tiglate (PCAC) from Paulownia Coreana. Molecules. 2012; 17(5):5945-5951. https://doi.org/10.3390/molecules17055945
Chicago/Turabian StyleJung, Samil, Hyung-In Moon, Jiyeon Ohk, Soonduck Lee, Chengping Li, Soo-Ki Kim, and Myeong-Sok Lee. 2012. "Inhibitory Effect and Mechanism on Antiproliferation of Isoatriplicolide Tiglate (PCAC) from Paulownia Coreana" Molecules 17, no. 5: 5945-5951. https://doi.org/10.3390/molecules17055945