The Relationship of Nitrogen and C/N Ratio with Secondary Metabolites Levels and Antioxidant Activities in Three Varieties of Malaysian Kacip Fatimah (Labisia pumila Blume)

Abstract
:1. Introduction
2. Results and Discussion
2.1. Total Phenolics and Total Flavonoids Profiling

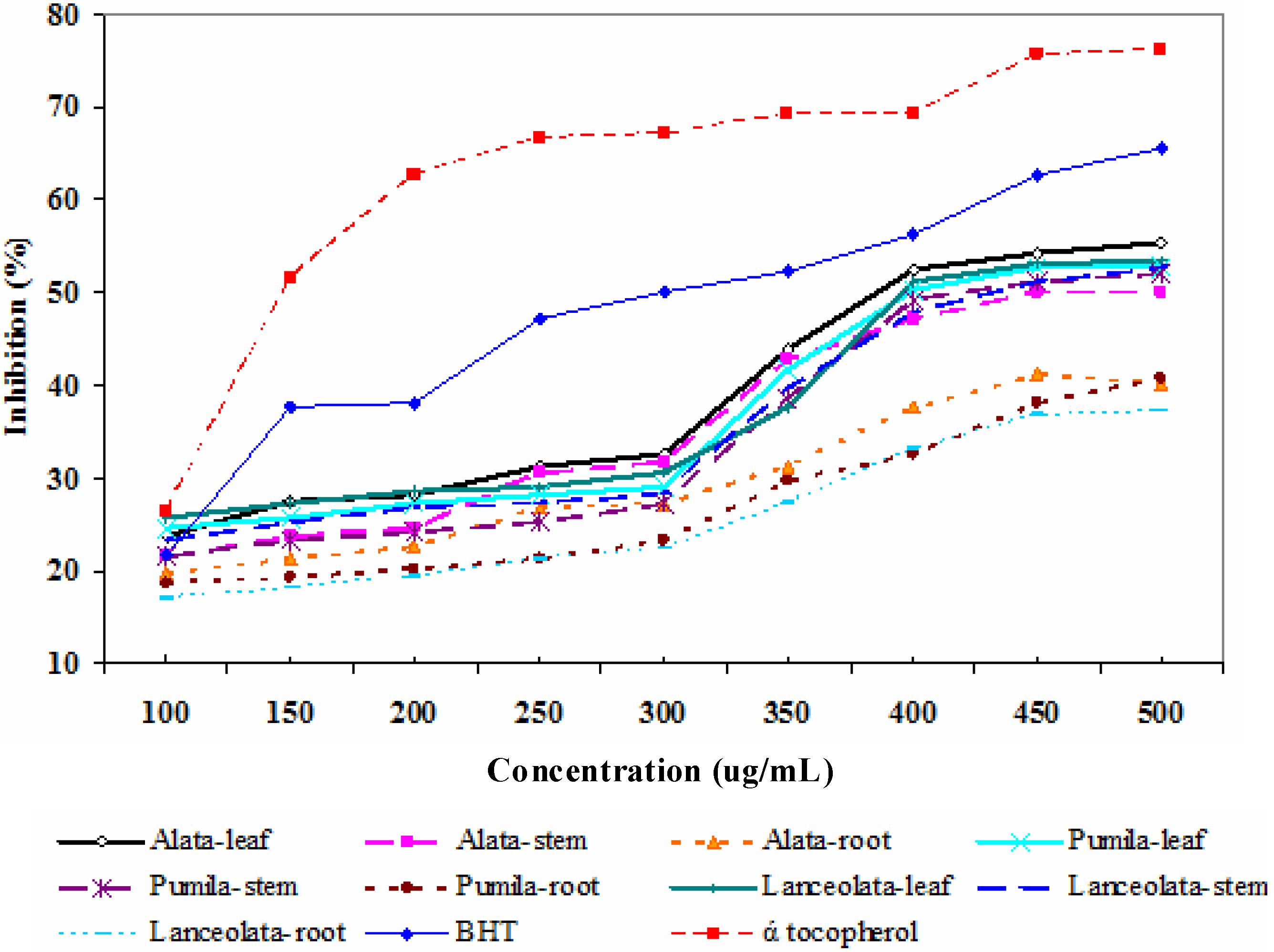{kind=link}
| Varieties | Plant parts | Total phenolics 1 | Total flavonoids 2 |
|---|---|---|---|
| Leaves | 1.64 ± 0.03 a,* | 0.91 ± 0.10 a,* | |
| Alata | Stems | 0.98 ± 0.02 b | 0.54 ± 0.05 b |
| Roots | 0.87 ± 0.02 c | 0.34 ± 0.01 c | |
| Leaves | 1.56 ± 0.02 a | 0.87 ± 0.01 a | |
| Pumila | Stems | 0.87 ± 0.04 b | 0.31 ± 0.07 b |
| Roots | 0.76 ± 0.03 c | 0.25 ± 0.05 c | |
| Leaves | 1.34 ± 0.12 a | 0.79 ± 0.02 a | |
| Lanceolata | Stems | 0.85 ± 0.02 b | 0.24 ± 0.0 b |
| Roots | 0.67 ± 0.01 c | 0.23 ± 0.03 c |
2.2. Radical Scavenging Activity
2.2.1. 1,1-Diphenyl-2-picrylhydrazyl (DPPH) assay

2.3. Reducing Ability
2.3.1. Ferric reducing antioxidant potential (FRAP)
| Varieties | Extract source | FRAP 1 |
|---|---|---|
| Leaves | 680.68 ± 34.51 c,* | |
| Alata | Stems | 537.65 ± 33.23 e |
| Roots | 435.23 ± 12.67 f | |
| Leaves | 600.45 ± 16.90 d | |
| Pumila | Stems | 512.56 ± 12.89 e |
| Roots | 401.23 ± 12.89 f | |
| Leaves | 599.87 ± 12.83 d | |
| Lanceolata | Stems | 425.67 ± 10.45 e |
| Roots | 421.61 ± 12.34 f | |
| Controls | BHT | 84.32 ± 56.34 g |
| α-tocopherol | 953.01 ± 45.67 b | |
| Vitamin C | 3301.25 ± 34.56 a |
| 1 | 2 | 3 | 4 | 5 | 6 | 7 | |
|---|---|---|---|---|---|---|---|
| 1.TP | 1.00 | ||||||
| 2.TF | 0.90 * | 1.00 | |||||
| 3. DPPH | 0.89 * | 0.78 * | 1.00 | ||||
| 4. FRAP | 0.87 * | 0.75 ** | 0.83 * | 1.00 | |||
| 5. C | 0.32 | 0.02 | 0.21 | 0.09 | 1.00 | ||
| 6. N | −0.87 * | −0.87 ** | −0.71 * | −0.67* | 0.42 | 1.00 | |
| 7. C/N | 0.71 * | 0.70 * | −0.72 * | −0.69* | 0.32 | −0.54 * | 1.00 |
2.4. Total Carbon, Nitrogen and C/N Profiling
| Varieties | Plant Parts | Carbon (%) | Nitrogen (%) | C/N |
|---|---|---|---|---|
| Leaves | 37.61 ± 0.21 a,* | 1.15 ± 0.45 c,* | 31.94 ± 0.34 a,* | |
| Alata | Stems | 38.22 ± 0.34 a | 1.87 ± 0.01 b | 20.44 ± 0.45 c |
| Roots | 37.32 ± 0.21 a | 1.95 ± 0.02 a | 19.24 ± 1.21 d | |
| Leaves | 38.74 ± 0.32 a | 1.31 ± 0.32 c | 29.36 ± 0.23 a | |
| Pumila | Stems | 37.21 ± 0.32 a | 1.72 ± 0.21 b | 21.19 ± 2.12 c |
| Roots | 36.87 ± 0.42 a | 1.85 ± 0.23 a | 20.43 ± 0.23 d | |
| Leaves | 38.13 ± 0.43 a | 1.55 ± 0.01 c | 25.31 ± 0.43 b | |
| Lanceolata | Stems | 36.52 ± 0.10 a | 1.73 ± 0.02 b | 21.46 ± 0.32 c |
| Roots | 37.21 ± 0.13 a | 1.93 ± 0.43 a | 19.05 ± 0.01 d |
3. Experimental
3.1. Plant Material and Maintenance
3.2. Extract Preparation
3.3. Quantification of Total Phenolics and Total Flavonoids
3.4. Determination of Antioxidant Activities
3.4.1. DPPH radical scavenging assay
3.4.2. Reducing ability (FRAP assay)
3.5. Total Carbon, Nitrogen and C:N Ratio
3.6. Statistical Analysis
4. Conclusions
Acknowledgements
References
- Devasagayam, P.A.; Tilak, J.C.; Boloor, K.K.; Sane, K.S.; Ghaskadbi, S.S.; Lele, R.D. Free radicals and antioxidants in human health: Current status and future prospects. J. Assoc. Phys. Ind. 2004, 52, 794–804. [Google Scholar]
- Serafini, M.; Bellocco, R.; Wolk, A.; Ekstrom, A.M. Total antioxidant potential of fruit and vegetables and risk of gastric cancer. Gastroenterology 2002, 123, 985–991. [Google Scholar] [CrossRef]
- Arai, Y.; Watanabe, S.; Kimira, M.; Shimoi, K.; Mochizuki, R.; Kinae, N. Dietary intakes of flavonols, flavones and isoflavones by Japanese women and the inverse correlation between quercetin intake and plasma LDL cholesterol concentration. J. Nutr. 2000, 130, 2243–2250. [Google Scholar]
- Galati, G.; O’Brien, P.J. Potential toxicity of flavonoids and other dietary phenolics: Signiflcance for thier chemoproventive and anticancer properties. Free Radic. Biol. Med. 2004, 32, 4–28. [Google Scholar]
- Bastianetto, S.; Zheng, W.H.; Quirion, R. Neuroprotective abilities of resveratrol and other red wine constituents against nitric oxide-related toxicity in cultured hippocampal neurons. Br. J. Pharmacol. 2000, 131, 711–720. [Google Scholar] [CrossRef]
- Mandel, S.; Youdim, M.B. Catechin polyphenols: Neurodegeneration and neuroprotection in neurodegenerative diseases. Free Radic. Biol. Med. 2004, 37, 304–317. [Google Scholar] [CrossRef]
- Shoskes, D.A.; Zeitlin, S.I.; Shahed, A.; Rajfer, J. Quercetin in men with category III chronic prostatitis: A preliminary prospective, double-blind, placebo-controlled trial. Urology 1999, 54, 960–963. [Google Scholar] [CrossRef]
- Schramm, D.D.; Karim, M.; Schrader, H.R.; Holt, R.R.; Cardetti, M.; Keen, C.L. Honey with high levels of antioxidants can provide protection to healthy human subjects. J. Agric. Food Chem. 2003, 51, 1732–1735. [Google Scholar]
- Shaheen, S.O.; Sterne, J.A.; Thompson, R.L.; Songhurst, C.E.; Margetts, B.M.; Burney, P.G. Dietary antioxidants and asthma in adults: Population-based case-control study. Am. J. Respir. Crit. Care Med. 2001, 164, 1823–1828. [Google Scholar]
- Halliwell, B.; Gutteridge, J.M.C. Free Radicals in Biology and Medicine, 2nd ed; Clarendon Press: Oxford, UK, 1989. [Google Scholar]
- Argolo, A.C.C.; Santa, A.; Pletsch, M.; Coelho, L.C.B. Antioxidant activity of leaf extract from Bauhinia mondora. Biosource Tech. 2004, 95, 229–233. [Google Scholar] [CrossRef]
- Rice, E.; Miller, N.J.; Pagangaa, G. Antioxidant properties of phenolics compounds. Trends Plant Sci. 1997, 2, 152–159. [Google Scholar] [CrossRef]
- Gulcin, I. The antioxidant and radical scavenging activities of black pepper seeds. Int. J. Food Sci. Nutr. 2005, 56, 491–499. [Google Scholar] [CrossRef]
- Aqil, F.; Ahmed, I.; Mehmood, Z. Antioxidant and free radical scavenging properties of twelve traditional used indian medicinal plants. Turk. J. Biol. 2006, 30, 177–183. [Google Scholar]
- Hras, A.R.; Hadolin, M.; Knez, Z.; Bauman, D. Comparison of antioxidative and synergistic effects of rosemary extract with alpha-tocopherol, ascorbyl palmitate and citric acid in sunflower oil. Food Chem. 2000, 71, 229–233. [Google Scholar] [CrossRef]
- Mccall, M.R.; Frei, B. Can antioxidant vitamins materially reduce oxidative damage in humans? Free Radic. Biol. Med. 1999, 26, 1034–1053. [Google Scholar] [CrossRef]
- Karimi, E.; Ehsan, O.; Hendra, R.; Jaafar, H.Z.E. Evaluation of Crocus sativus L. stigma phenolics and flavonoid compounds and its antioxidant activity. Molecules 2010, 15, 6244–6256. [Google Scholar] [CrossRef]
- Velioglu, Y.S.; Mazza, G.; Gao, Y.L.; Oomah, B.D. Antioxidant activity and total phenolics in selected fruits, vegetable and grain products. J. Agric. Food Chem. 1998, 46, 4113–4117. [Google Scholar] [CrossRef]
- Ibrahim, M.H.; Jaafar, H.Z.E.; Rahmat, A.; Zaharah, A.R. The relationship between phenolics and flavonoid production with total non structural carbohydrate and photosynthetic rate in Labisia pumila Benth. Under High CO2 and nitrogen fertilization. Molecules 2011, 16, 162–174. [Google Scholar]
- Jaafar, H.Z.E.; Mohamed, H.N.B.; Rahmat, A. Accumulation and partitioning of total phenols in two varieties of Labisia pumila Benth. under manipulation of greenhouse irradiance. Acta Hort. 2008, 797, 387–392. [Google Scholar]
- Tsao, R.; Deng, Z. Separation procedures for naturally occurring antioxidant photochemicals. J. Chrom. 2004, 812, 85–99. [Google Scholar]
- Zhang, Q.; Ye, M. Chemical analysis of the Chinese herbal medicine Gan-Cao (licorice). J. Chrom. 2008, 1216, 1954–1969. [Google Scholar]
- Rozihawati, Z.; Aminah, H.; Lokman, N. Preliminary trials on the rooting ability of Labisia pumila cuttings. In Malaysia Science and Technology Congress 2003; Agricultural Sciences: Kuala Lumpur, Malaysia, 2003. [Google Scholar]
- Stone, B.C. Notes on the genus Labisia Lindyl (Myrsinaceae). Malayan Nat. J. 1988, 42, 43–51. [Google Scholar]
- Ibrahim, M.H.; Jaafar, H.Z.E.; Rahmat, A.; Abdul Rahman, Z. The relationship between phenolics and flavonoids production with total non structural carbohydrate and photosynthetic rate in Labisia pumila Benth. under high CO2 and nitrogen fertilization. Molecules 2011, 16, 162–174. [Google Scholar]
- Bryant, J.P.; Chapin, F.S.; Klein, D.R. Carbon nutrient balance of boreal plants in relation to vertebrate herbivory. Oikos 1983, 40, 357–368. [Google Scholar] [CrossRef]
- Lindroth, R.L.; Osier, T.L.; Barnhill, H.R.H.; Wood, S.A. Effects of genotype and nutrient availability on photochemistry of trembling aspen (Populus tremoides Mich.) during leaf senescence. Biochem. Sys. Ecol. 2002, 30, 297–307. [Google Scholar] [CrossRef]
- Lindroth, R.L. Impacts of eelvated atmospheric CO2 and O3 on forest: Phytochemistry, trophic interactions, and ecosystem dynamics. J. Chem. Ecol. 2010, 36, 2–21. [Google Scholar] [CrossRef]
- Norhaiza, M.; Maziah, M.; Hakiman, M. Antioxidative properties of leaf extracts of popular Malaysian herb, Labisia pumila. J. Med. Plant Res. 2009, 3, 217–223. [Google Scholar]
- Karimi, E.; Jaafar, H.Z.E.; Ahmad, S. Phytochemical analysis and antimicrobial activites of methanolics extracts of leaf, stem and root from different varieties of Labisia pumila Benth. Molecules 2011, 16, 4438–4450. [Google Scholar] [CrossRef]
- Ferry, D.R.; Smith, A.; Malkhandi, J. Phase I clinical trial of the flavonoid quercetin: Pharmacokinetics and evidence for in vivo tyrosine kinase inhibition. Clin. Cancer Res. 1996, 2, 659–668. [Google Scholar]
- Ranelletti, F.O.; Maggiano, N.; Serra, F.G. Quercetin inhibits p21-ras expression in human colon cancer cell lines and in primary colorectal tumors. Int. J. Cancer 1999, 85, 438–445. [Google Scholar]
- Pathak, S.B.; Niranjan, K.; Padh, H.; Rajani, M. TLC densitometric method for the quantification of eugenol and gallic acid in clove. Chromatographia 2004, 60, 241–244. [Google Scholar]
- Enkhmaa, B.L. Mulberry (Morus alba L.) leaves and their major flavonol quercetin 3-(6-malonylglucoside) attenuate atherosclerotic lesion development in LDL receptor-deficient mice. J. Nutr. 2005, 135, 729–734. [Google Scholar]
- Luo, Y.L. Inhibition of cell growth and VEGF expression in ovarian cancer cells by flavonoids. Nut. Cancer 2008, 60, 800–809. [Google Scholar] [CrossRef]
- Wong, C.C.; Li, H.B.; Cheng, K.W.; Chen, F. A systematic survey of antioxidant activity of 30 Chinese medicinal plants using the ferric reducing antioxidant power assay. Food Chem. 2006, 97, 705–711. [Google Scholar] [CrossRef]
- Miliauskas, G.; Venskutonis, P.R.; Van Beek, T.A. Screening of radical scavenging activity of some medicinal and aromatic plant extracts. Food Chem. 2004, 85, 231–237. [Google Scholar] [CrossRef]
- Tamura, H.; Yamagami, A. Antioxidative activity of monoacylated anthocyanins isolated from Muscat bailey A grape. J. Agric. Food Chem. 1994, 42, 1612–1615. [Google Scholar] [CrossRef]
- Bravo, L. Polyphenols: Chemistry, dietary sources, metabolism, and nutritional significance. Nutr. Rev. 1998, 56, 317–333. [Google Scholar] [CrossRef]
- Chanwitheesuk, A.; Teerawutgulrag, A.; Rakariyatham, N. Screening of antioxidant activity and antioxidant compounds of some edible plants of Thailand. Food Chem. 2004, 92, 491–497. [Google Scholar]
- Luximon-Ramma, A.; Bahorun, T.; Soobrattee, A.M.; Aruoma, O.I. Antioxidant activities of phenolic, proanthocyanidin and flavonoid components in extracts of Acacia fistula. J. Agric. Food Chem. 2005, 50, 5042–5047. [Google Scholar]
- Agnaniet, H.; Menut, C.; Bessie, J.M. Aromatic plants of tropical central Africa. Part XLIX: Chemical composition of essential oils of the leaf and rhizomes of Aframomum giganteum from Gabon. Flavour Fragrance 2004, 19, 205–209. [Google Scholar]
- Yen, G.C.; Duh, P.D. Scavenging effects of methanolic extract of peanut hulls on free-radical and active oxygen species. J. Agric. Food Chem. 1994, 42, 629–632. [Google Scholar] [CrossRef]
- Gil, M.I.; Thomas-Barberan, F.T.; Hess-Pirce, B.; Kader, A.A. Antioxidant capacities, phenolic compounds, carotenoids and vitamin C contents of nectarine, peach and plum cultivars from California. J. Agric. Food Chem. 2002, 50, 4976–4982. [Google Scholar] [CrossRef]
- Ruan, J.; Haerdter, R.; Gerendás, J. Impact of nitrogen supply on carbon/nitrogen allocation: A case study on amino acids and catechins in green tea [Camellia sinensis (L.) O. Kuntze]. Plant Biol. 2010, 12, 724–734. [Google Scholar] [CrossRef]
- Pompelli, M.F.; Martins, S.C.V.; Antunes, W.C.; Chaves, A.R.M.; DaMatta, F.M. Photosynthesis and photoprotection in coffee leaves is affected by nitrogen and light availabilities in winter conditions. J. Plant Physiol. 2010, 167, 1052–1060. [Google Scholar] [CrossRef]
- Guo, W.; Liu, R.; Zhou, Z.; Chen, B.; Xu, N. Waterlogging of cotton calls for caution with N fertilization. Acta Agri. Scand. 2010, 60, 450–459. [Google Scholar]
- Okano, K.; Totsuka, T. Absorption of nitrogen dioxide by sunflower plants grown at various levels of nitrate. New Phytol. 1986, 102, 551–562. [Google Scholar] [CrossRef]
- Jaafar, H.Z.E.; Ibrahim, M.H.; Por, L.S. Effects of CO2 enrichment on accumulation of total phenols, flavonoid and chlorophyll content in two varieties of Labisia pumila Benth. exposed to different shade levels. In Proceedings of International Conference on Balanced Nutrient Management for Tropical Agriculture, Kuantan, Pahang, Malaysia, 15–22 February 2010; UPM: Kuala Lumpur, Malaysia, 2010; pp. 112–114. [Google Scholar]
- Benzie, I.F.F.; Strain, J.J. The ferric reducing ability of plasma (FRAP) as a measure of ‘Antioxidant Power’: The FRAP assay. Anal. Biochem. 1996, 239, 70–76. [Google Scholar]
- Ibrahim, M.H.; Jaafar, H.Z.E.; Haniff, M.; Yusop, R. Changes in the growth and photosynthetic patterns of oil palm (Elaeis guineensis Jacq.) seedlings exposed to short term CO2 enrichment in a Closed Top Chamber. Acta Physiol. Plant 2010, 32, 305–313. [Google Scholar] [CrossRef]
- Sample Availability: Not Available.
© 2011 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license ( http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Ibrahim, M.H.; Jaafar, H.Z.E. The Relationship of Nitrogen and C/N Ratio with Secondary Metabolites Levels and Antioxidant Activities in Three Varieties of Malaysian Kacip Fatimah (Labisia pumila Blume). Molecules 2011, 16, 5514-5526. https://doi.org/10.3390/molecules16075514
Ibrahim MH, Jaafar HZE. The Relationship of Nitrogen and C/N Ratio with Secondary Metabolites Levels and Antioxidant Activities in Three Varieties of Malaysian Kacip Fatimah (Labisia pumila Blume). Molecules. 2011; 16(7):5514-5526. https://doi.org/10.3390/molecules16075514
Chicago/Turabian StyleIbrahim, Mohd Hafiz, and Hawa Z.E. Jaafar. 2011. "The Relationship of Nitrogen and C/N Ratio with Secondary Metabolites Levels and Antioxidant Activities in Three Varieties of Malaysian Kacip Fatimah (Labisia pumila Blume)" Molecules 16, no. 7: 5514-5526. https://doi.org/10.3390/molecules16075514