Lichtheimia blakesleeana as a New Potencial Producer of Phytase and Xylanase

Abstract
:1. Introduction
2. Results and Discussion
2.1. Optimization of Process Parameters for Phytase Production

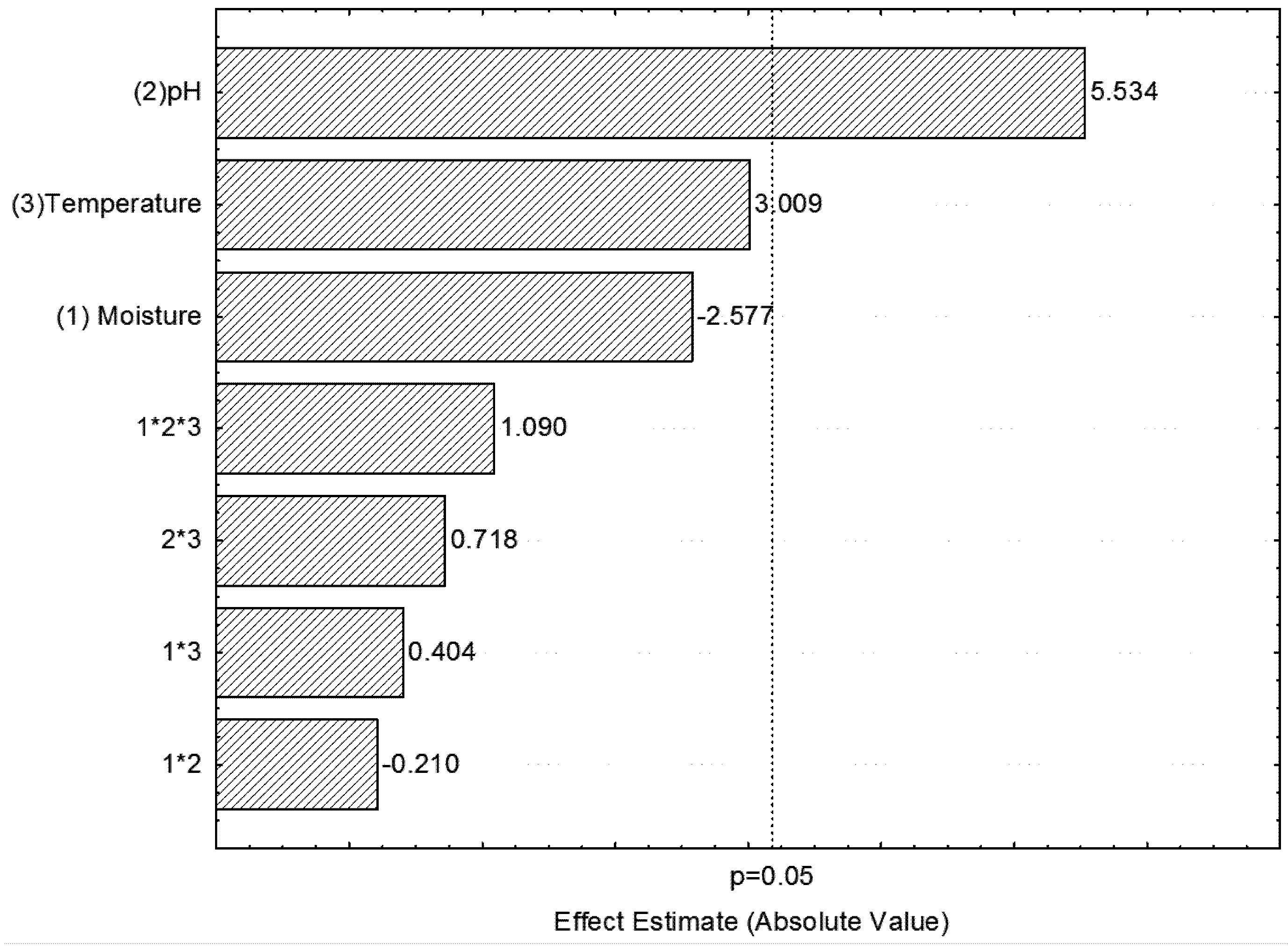{kind=link}
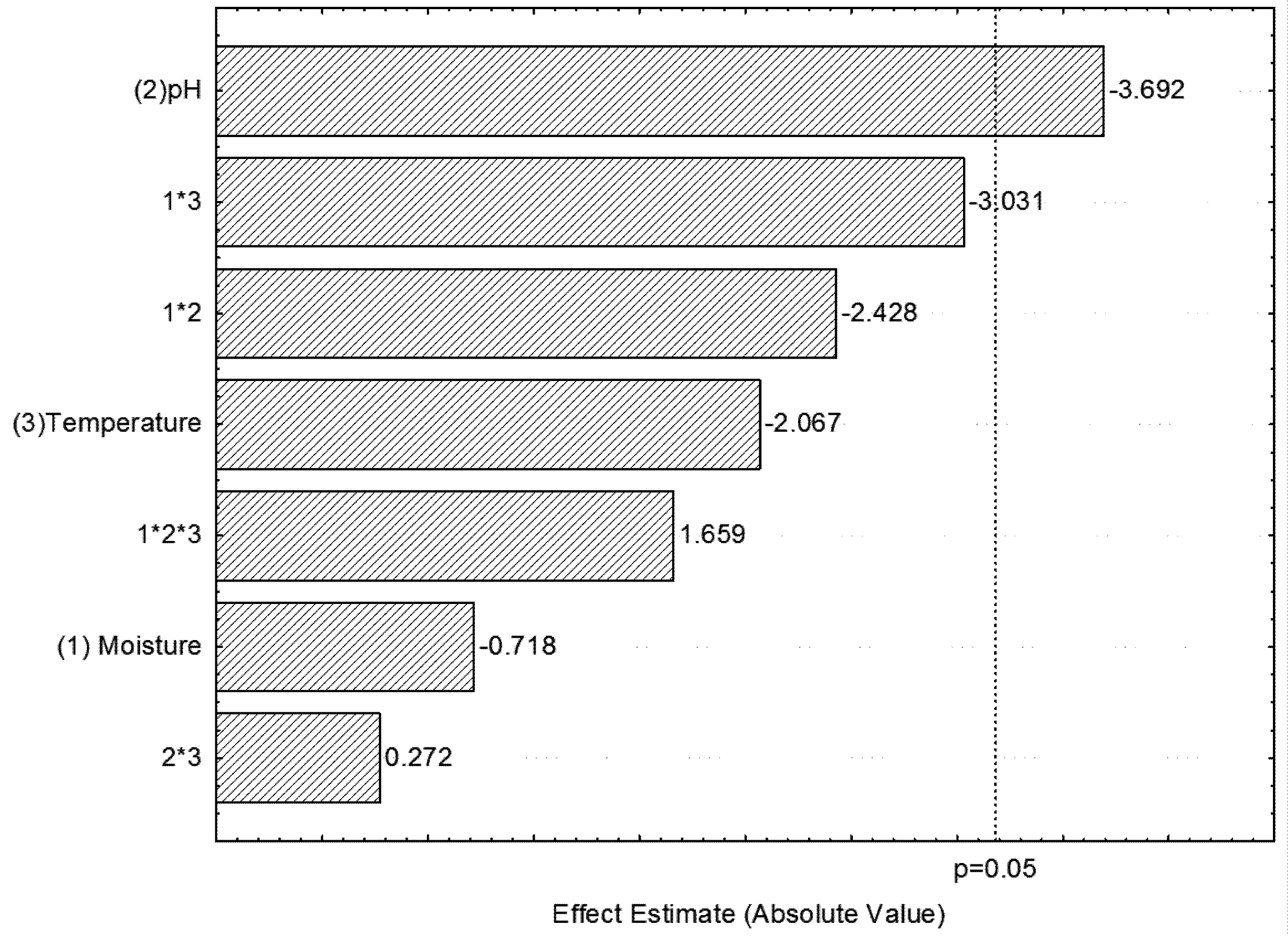{kind=link}
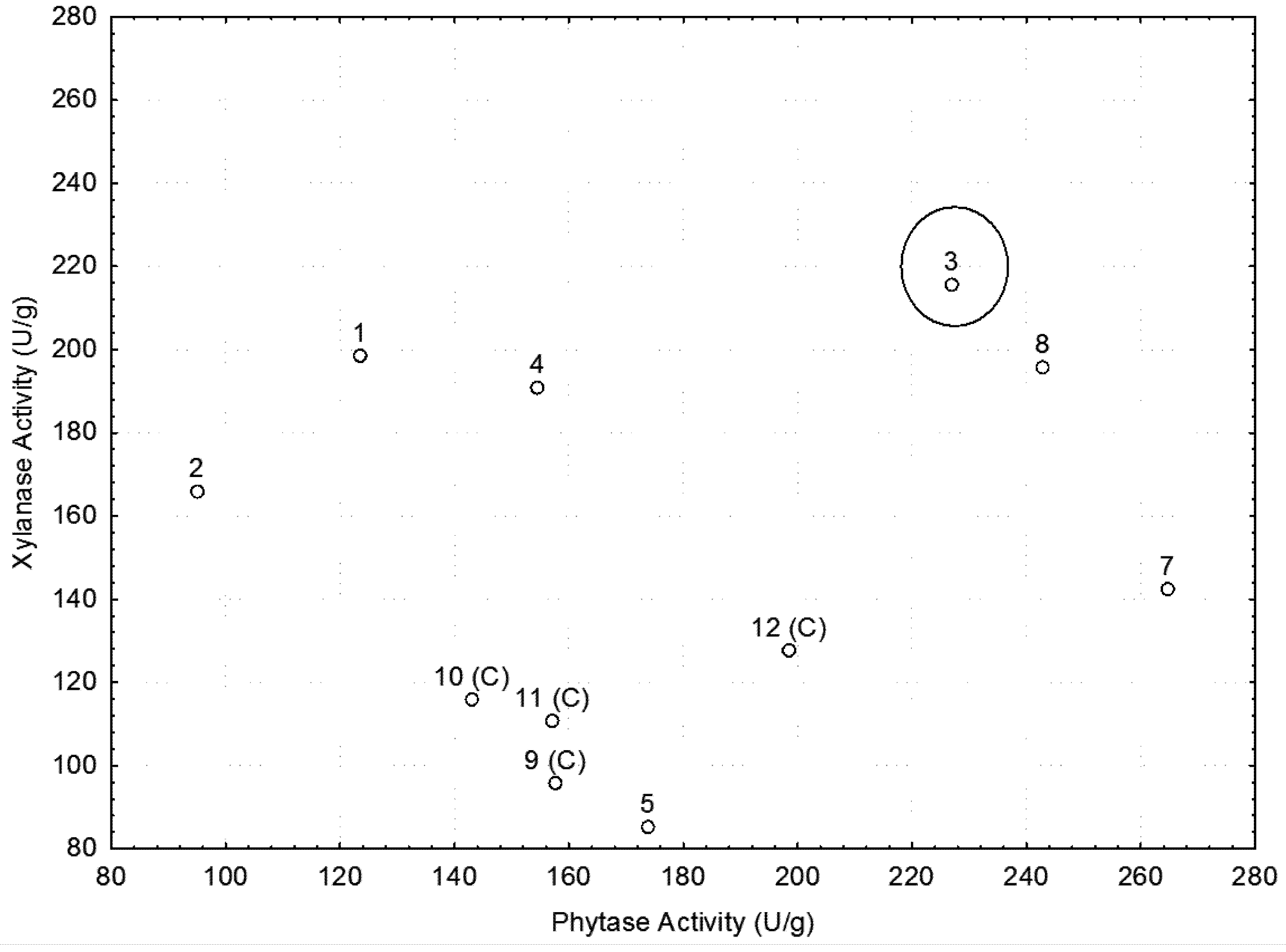{kind=link}
| Runs | Time (hours) | ||||
|---|---|---|---|---|---|
| 24 | 48 | 72 | 96 | 120 | |
| 1 | 98.16 | 123.48 | 161.77 | 126.74 | 130.76 |
| 2 | 67.36 | 95.04 | 118.43 | 121.70 | 74.24 |
| 3 | 150.88 | 226.92 | 198.82 | 195.48 | 129.79 |
| 4 | 125.28 | 154.48 | 142.80 | 112.71 | 106.84 |
| 5 | 120.23 | 173.81 | 226.05 | 103.08 | 101.65 |
| 6 | 159.83 | 122.18 | 175.43 | 69.78 | 72.01 |
| 7 | 188.50 | 264.68 | 131.82 | 132.00 | 128.07 |
| 8 | 204.29 | 242.80 | 100.31 | 93.49 | 101.03 |
| 9 | 205.22 | 157.65 | 178.79 | 95.57 | 88.45 |
| 10 | 180.12 | 143.02 | 93.50 | 72.22 | 107.03 |
| 11 | 172.16 | 157.07 | 101.86 | 87.15 | 113.67 |
| 12 | 162.98 | 198.46 | 94.74 | 99.96 | 95.39 |

2.2. Optimization of Process Parameters for Xylanase Production
| Runs | Time (hours) | ||||
|---|---|---|---|---|---|
| 24 | 48 | 72 | 96 | 120 | |
| 1 | 174.93 | 198.53 | 222.17 | 222.28 | 291.70 |
| 2 | 158.27 | 165.89 | 164.46 | 384.25 | 397.82 |
| 3 | 221.31 | 215.59 | 229.23 | 225.38 | 214.57 |
| 4 | 173.76 | 190.85 | 189.38 | 180.49 | 177.24 |
| 5 | 48.51 | 85.25 | 174.67 | 281.76 | 288.51 |
| 6 | 31.11 | 74.92 | 107.29 | 206.35 | 273.62 |
| 7 | 62.05 | 142.44 | 180.60 | 214.69 | 185.30 |
| 8 | 118.05 | 195.73 | 140.22 | 100.34 | 96.50 |
| 9 | 58.54 | 95.80 | 155.65 | 176.45 | 226.69 |
| 10 | 87.95 | 115.90 | 136.53 | 205.05 | 195.85 |
| 11 | 56.96 | 110.75 | 130.11 | 229.88 | 261.11 |
| 12 | 63.43 | 127.71 | 134.19 | 147.08 | 179.72 |

2.3. The Best Conditions for Phytase and Xylanase Production

3. Experimental
3.1. Microorganism and Inoculum Preparation
3.2. Substrate
3.3. Experimental Design of Solid-State Fermentation
| Runs * | Variables | ||
|---|---|---|---|
| Moisture (%) | pH | Temperature (°C) | |
| 1 | 50 | 4.0 | 26 |
| 2 | 70 | 4.0 | 26 |
| 3 | 50 | 6.0 | 26 |
| 4 | 70 | 6.0 | 26 |
| 5 | 50 | 4.0 | 34 |
| 6 | 70 | 4.0 | 34 |
| 7 | 50 | 6.0 | 34 |
| 8 | 70 | 6.0 | 34 |
| 9 (C) | 60 | 5.0 | 30 |
| 10 (C) | 60 | 5.0 | 30 |
| 11 (C) | 60 | 5.0 | 30 |
| 12 (C) | 60 | 5.0 | 30 |
3.4. Extraction of Enzymes
3.5. Phytase Activity
3.6. Xylanase Activity
4. Conclusions
Acknowledgements
References
- Singh, B.; Satyanarayana, T. Phytase production by a thermophilic mould Sporotrichum thermophile in solid state fermentation and its potential applications. Bioresour. Technol. 2008, 99, 2824–2830. [Google Scholar] [CrossRef]
- Roopesh, K.; Ramachandran, S.; Nampoothiri, K.N.; Szakacs, G.; Pandey, A. Comparison of phytase production on wheat bran and oilcakes in solid-state fermentation by Mucor racemosus. Bioresour. Technol. 2006, 97, 506–511. [Google Scholar] [CrossRef]
- Pandey, A.; Larroche, C.; Soccol, C.R.; Dussap, C.G. Advances in Fermentation Technology; Asiatech Publishers, Inc.: New Delhi, India, 2008; p. 672. [Google Scholar]
- Pandey, A.; Selvakumar, P.; Soccol, C.R.; Nigam, P. Solid state fermentation for the production of industrial enzymes. Curr. Sci. 1999, 77, 149–162. [Google Scholar]
- Gunashree, B.S.; Venkateswaran, G. Effect of different cultural conditions for phytase production by Aspergillus niger CFR 335 in submerged and solid-state fermentations. J. Ind. Microbiol. Biotechnol. 2008, 35, 1587–1596. [Google Scholar] [CrossRef]
- Martin, N.; Guez, M.A.U.; Sette, L.D.; Da Silva, R.; Gomes, E. Pectinase production by a brazilian thermophilic fungus Thermomucor indicae-seudaticae N31 in solid state and submerged fermentation. Microbiology 2010, 79, 306–313. [Google Scholar] [CrossRef]
- Nielsen, M.M.; Damstrup, M.L.; Hansen, A. An optimised micro-titer plate method for characterisation of endogenous rye phytase under industrial rye bread making conditions. Eur. Food Res. Technol. 2008, 227, 1009–1015. [Google Scholar] [CrossRef]
- Pandey, A.; Soccol, C.R.; Rodriguez-Leon, J.A.; Nijam, P. Solid state Fermentation in Biotechnology: Fundamentals and Applications; Asiatech Publishers, Inc.: New Delhi, India, 2001; p. 221. [Google Scholar]
- Bogar, B.; Szakacs, G.; Linden, J.C.; Pandey, A.; Tengerdy, R.P. Optimization of phytase production by solid substrate fermentation. J. Ind. Microbiol. Biotechnol. 2003, 30, 183–189. [Google Scholar]
- Cao, L.; Wang, W.; Yang, C.; Yang, Y.; Diana, J.; Yakupitiyage, A.; Luo, Z.; Li, D. Application of microbial phytase in fish feed. Enzyme Microb. Technol. 2007, 40, 497–507. [Google Scholar] [CrossRef]
- Bhat, M.K. Cellulases and related enzymes in biotechnology. Biotechnol. Adv. 2000, 18, 355–383. [Google Scholar] [CrossRef]
- Alberton, L.R.; Vandenberghe, L.P.S.; Assman, R.; Munchen, L.; Ciffoni, E.M.G.; Pachaly, J.R.; Soccol, C.R. Effect of the Streptomyces viridosporus T7A crude fermentation extract on the in vitro digestibility of dry matter for ruminants. Arq. Ciên. Vet. Zool. 2006, 9, 29–36. [Google Scholar]
- Aguilar, C.N.; Gutiérrez-Sánchez, G.; Rado-Barragán, P.A.; Rodríguez-Herrera, R.; Martínez-Hernandez, J.L.; Contreras-Esquivel, J.C. Perspectives of solid state fermentation for production of food enzymes. Am. J. Biochem. Biotechnol. 2008, 4, 354–366. [Google Scholar] [CrossRef]
- Marín, F.R.; Soler-Rivas, C.; Benavente-García, O.; Castillo, J.; Pérez-Alvarez, J.A. By-products from different citrus processes as a source of customized functional fibres. Food Chem. 2007, 100, 736–741. [Google Scholar] [CrossRef]
- Spier, M.R.; Greiner, R.; Rodriguez-León, J.A.; Woiciechowski, A.L.; Pandey, A.; Soccol, V.T.; Soccol, C.R. Phytase production using citric pulp and other residues of the agroindustry in SSF by fungal isolates. Food Techol. Biotechnol. 2008, 46, 178–182. [Google Scholar]
- Izquierdo, L.; Sendra, J.M. Citrus fruits composition and characterization. In Encyclopedia of Food Sciences and Nutrition; Caballero, B., Trugo, L., Finglas, P., Eds.; Academic Press: Oxford, UK, 2003; pp. 1335–1341. [Google Scholar]
- Pascual, J.; Carmona, J.F. Composition of citrus pulp. Anim. Feed Sci. Technol. 1980, 5, 1–10. [Google Scholar] [CrossRef]
- Orange By-Products: ABE Citrus. The Brazilian Association for Citrus Exporters: São Paulo, Brazil, 2007.
- El-Batal, A.I.; Karem, H.A. Phytase production and phytic acid reduction in rapeseed meal by Aspergillus niger during solid state fermentation. Food Res. Int. 2001, 34, 715–720. [Google Scholar] [CrossRef]
- Botella, C.; Diaz, A.; Ory, I.; Webb, C.; Blandino, A. Xylanase and pectinase production by Aspergillus awamori on grape pomace in solid state fermentation. Process Biochem. 2007, 42, 98–101. [Google Scholar] [CrossRef]
- Chadha, B.S.; Harmeet, G.; Mandeep, M.; Saini, H.S.; Singh, N. Phytase production by the thermophilic fungus Rhizomucor pusillus. World J. Microbiol. Biotechnol. 2004, 20, 105–109. [Google Scholar] [CrossRef]
- Vassilev, N.; Vassileva, M.; Bravo, V.; Fernández-Serrano, M.; Nikolaeva, I. Simultaneous phytase production and rock phosphate solubilization by Aspergillus niger grown on dry olive wastes. Ind. Crop Prod. 2007, 26, 332–336. [Google Scholar] [CrossRef]
- Gervais, P.; Molin, P. The role of water in solid-state fermentation. Biochem. Eng. J. 2003, 13, 85–101. [Google Scholar] [CrossRef]
- Shah, A.R.; Madamwar, D. Xylanase production by a newly isolated Aspergillus foetidus strain and its characterization. Process Biochem. 2005, 40, 1763–1771. [Google Scholar] [CrossRef]
- Camassola, M.; Dillon, A.J.P. Production of cellulases and hemicellulases by Penicillium echinulatum grown on pretreated sugar cane bagasse and wheat bran in solid-state fermentation. J. Appl. Microbiol. 2007, 103, 2196–2204. [Google Scholar] [CrossRef]
- Zenebon, O.; Pascuet, N.S. Métodos Físico-químicos para Análise de Alimentos, 4th ed; Instituto Adolfo Lutz: São Paulo, Brazil, 2005. [Google Scholar]
- Bruns, R.E.; Scarminio, I.S.; Neto, B.B. Statistical Design-Chemometrics, 1st ed; Elsevier: Amsterdam, The Netherlands, 2006; p. 422. [Google Scholar]
- Spier, M.R.; Woiciechowski, A.L.; Letti, L.A.; Scheidt, G.N.; Sturm, W.; Rodriguez-León, J.A.; Carvalho, J.C.; Dergint, D.E.; Soccol, C.R. Monitoring fermentation parameters during phytase production in column-type bioreactor using a new data acquisition system. Bioprocess Biosyst. Eng. 2010, 33, 1033–1041. [Google Scholar] [CrossRef]
- Bailey, M.J.; Biely, P.; Poutanen, K. Interlaboratory testing of methods for assay of xilanase activity. J. Biotechnol. 1992, 23, 257–270. [Google Scholar]
- Miller, G.L. Use of dinitrosalicylic acid reagent for determination of reducing sugars. Anal. Chem. 1959, 31, 426–428. [Google Scholar] [CrossRef]
- Sample Availability: Not available.
© 2011 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license ( http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Neves, M.L.C.; Silva, M.F.d.; Souza-Motta, C.M.; Spier, M.R.; Soccol, C.R.; Porto, T.S.; Moreira, K.A.; Porto, A.L.F. Lichtheimia blakesleeana as a New Potencial Producer of Phytase and Xylanase. Molecules 2011, 16, 4807-4817. https://doi.org/10.3390/molecules16064807
Neves MLC, Silva MFd, Souza-Motta CM, Spier MR, Soccol CR, Porto TS, Moreira KA, Porto ALF. Lichtheimia blakesleeana as a New Potencial Producer of Phytase and Xylanase. Molecules. 2011; 16(6):4807-4817. https://doi.org/10.3390/molecules16064807
Chicago/Turabian StyleNeves, Maria Luiza Carvalho, Milena Fernandes da Silva, Cristina Maria Souza-Motta, Michele Rigon Spier, Carlos Ricardo Soccol, Tatiana Souza Porto, Keila Aparecida Moreira, and Ana Lúcia Figueiredo Porto. 2011. "Lichtheimia blakesleeana as a New Potencial Producer of Phytase and Xylanase" Molecules 16, no. 6: 4807-4817. https://doi.org/10.3390/molecules16064807