Inter-Population Variability of Terpenoid Composition in Leaves of Pistacia lentiscus L. from Algeria: A Chemoecological Approach

Abstract
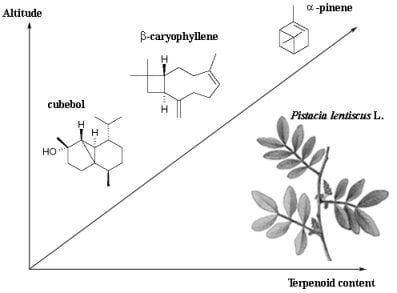
:
1. Introduction
2. Results and Discussion

{kind=link}
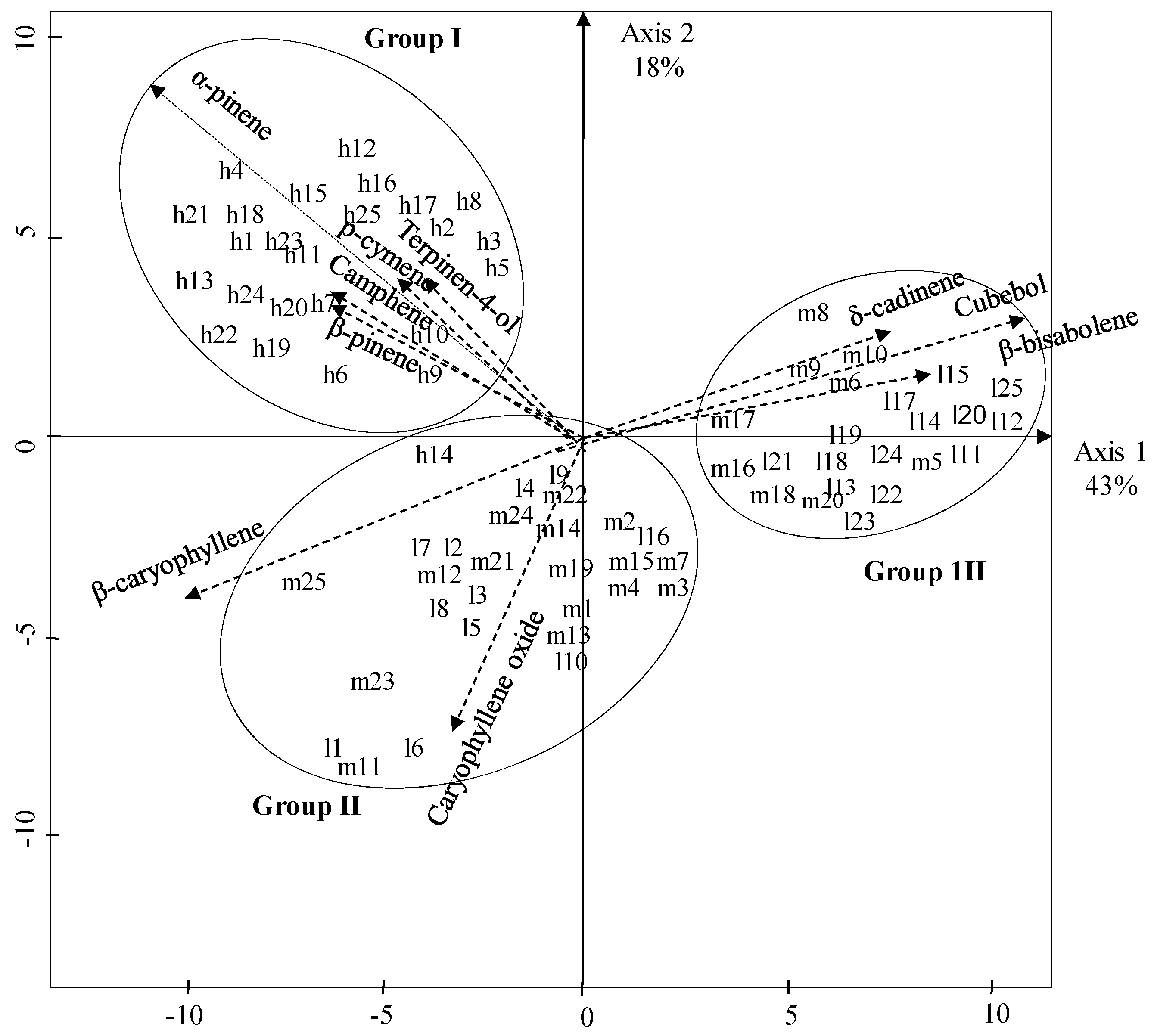{kind=link}
| Compounds | Mean concentration ± standard deviation of terpenoid contents in leaves extracts of Pistacia lentiscus (µg.g dw−1) | |||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| RI | Low altitude | Mid altitude | High altitude | |||||||||||||
| L1(n = 5) | L2(n = 5) | L3(n = 5) | L4(n = 5) | L5(n = 5) | M1(n = 5) | M2(n = 5) | M3(n = 5) | M4(n = 5) | M5(n = 5) | H1(n = 5) | H2(n = 5) | H3(n = 5) | H4(n = 5) | H5(n = 5) | ||
| α-thujene | 926 | 4.8±1.6 | 3.8±1.3 | 1.1±0.9 | 1.4±0.9 | 4.6±1.5 | 8.2±1.1 | 8.1±0.4 | 4.5±0.9 | 3.4±0.5 | 6.5±1.2 | 13.6±0.7 | 15.2±0.7 | 12.5±1.5 | 15.1±0.7 | 16.9±0.5 |
| α-pinene | 933 | 8.1±3.3 | 7.6±3.0 | 2.9±1.1 | 2.7±0.9 | 5.5±1.2 | 2.2±0.2 | 5.7±0.9 | 5.2±0.8 | 1.9±0.3 | 13.5±2.0 | 102.9±6.2 | 100.9±3.6 | 76.3±14.2 | 105.0±4.5 | 98.0±0.5 |
| camphene | 947 | 0.6±0.2 | 0.6±0.2 | 0.3±0.1 | 0.3±0.1 | 0.4±0.1 | 0.9±0.2 | 3.0±1.1 | 1.1±0.5 | 0.9±0.3 | 2.2±0.7 | 14.3±1.3 | 17.1±1.8 | 12.8±3.4 | 15.9±2.1 | 20.3±0.6 |
| β-pinene | 973 | 12.2±4.3 | 10.9±3.8 | 3.0±1.5 | 3.3±1.5 | 7.9±2.3 | 11.5±1.3 | 12.1±1.8 | 11.4±3.0 | 4.5±0.4 | 14.5±2.2 | 35.3±7.4 | 36.7±7.1 | 56.2±14.9 | 44.3±8.8 | 30.9±0.6 |
| α-terpinene | 1018 | 2.0±0.9 | 1.9±0.7 | 0.1±0.1 | 0.6±0.3 | 1.9±0.6 | 7.3±0.7 | 6.2±1.4 | 2.9±1.1 | 1.3±0.3 | 1.9±0.4 | 7.9±0.6 | 9.3±0.5 | 8.5±1.4 | 9.5±0.4 | 10.5±0.4 |
| p-cymene | 1027 | 0.9±0.4 | 0.9±0.4 | 0.4±0.1 | 0.3±0.1 | 0.7±0.1 | 1.9±0.4 | 3.5±0.9 | 1.4±0.5 | 0.7±0.3 | 1.6±0.6 | 15.7±0.7 | 17.7±0.5 | 21.3±4.8 | 18.5±0.5 | 18.8±0.4 |
| limonene | 1029 | 2.3±0.2 | 2.3±0.3 | 1.7±0.0 | 1.5±0.1 | 2.1±0.1 | 5.8±0.5 | 5.7±1.6 | 2.9±0.9 | 2.4±0.3 | 4.0±0.4 | 11.5±2.6 | 13.6±2.2 | 32.6±12.3 | 16.3±2.7 | 12.7±0.3 |
| γ-terpinene | 1062 | 2.1±0.6 | 1.7±0.6 | 0.5±0.3 | 0.8±0.4 | 2.0±0.6 | 4.7±0.4 | 5.6±1.0 | 2.5±0.8 | 1.4±0.3 | 2.4±0.4 | 7.2±0.6 | 8.9±0.7 | 7.7±0.9 | 8.8±0.7 | 10.5±0.4 |
| terpinen-4-ol | 1179 | 5.1±1.2 | 4.0±0.7 | 3.5±0.6 | 2.4±0.1 | 3.1±0.4 | 5.9±0.8 | 7.0±1.3 | 4.6±0.9 | 4.1±0.4 | 6.3±0.6 | 27.4±0.6 | 29.6±0.4 | 42.5±7.8 | 30.3±0.4 | 30.5±0.4 |
| α-terpineol | 1197 | 2.4±0.6 | 1.6±0.5 | 0.8±0.2 | 0.5±0.2 | 1.4±0.1 | 1.9±0.3 | 3.5±1.0 | 2.0±0.5 | 1.1±0.3 | 2.0±0.7 | 5.1±0.8 | 7.5±0.5 | 6.0±1.1 | 8.4±0.6 | 8.2±0.4 |
| borneol acetate | 1287 | 6.8±1.9 | 5.6±1.5 | 3.9±0.2 | 3.6±0.2 | 3.2±0.4 | 7.3±0.8 | 6.5±1.1 | 6.3±1.5 | 6.9±0.7 | 18.7±6.2 | 18.6±0.7 | 22.3±0.5 | 25.8±6.9 | 22.4±0.5 | 23.6±0.4 |
| α-terpineol acetate* | 1351 | 4.4±0.2 | 4.0±0.6 | 10.3±1.0 | 8.8±0.5 | 8.9±2.9 | 17.7±2.3 | 10.4±1.5 | 5.2±1.4 | 7.9±0.4 | 14.1±3.8 | 15.3±0.3 | 16.5±0.2 | 11.7±1.5 | 16.7±0.2 | 17.2±0.2 |
| δ-elemene* | 1338 | 0.5±0.1 | 0.5±0.1 | 0.6±0.1 | 0.4±0.0 | 0.0±0.0 | 1.2±0.3 | 2.8±0.9 | 0.7±0.4 | 1.7±0.3 | 7.0±3.1 | 0.4±0.2 | 2.6±0.5 | 1.4±0.9 | 2.7±0.4 | 3.9±0.4 |
| α-cubebene* | 1351 | 4.4±0.2 | 4.0±0.6 | 10.3±1.0 | 8.8±0.5 | 8.9±2.9 | 17.7±2.3 | 10.4±1.5 | 5.2±1.4 | 7.9±0.4 | 14.1±3.8 | 15.3±0.3 | 16.5±0.2 | 11.7±1.5 | 16.7±0.2 | 17.2±0.2 |
| α-copaene | 1377 | 7.5±1.3 | 8.3±1.0 | 10.4±1.1 | 8.5±0.6 | 13.7±3.1 | 22.9±3.7 | 10.9±0.8 | 9.4±0.3 | 8.8±0.5 | 13.9±1.0 | 34.8±2.4 | 38.3±1.5 | 23.2±4.7 | 40.3±1.9 | 37.8±0.4 |
| β-bourbonene* | 1389 | 1.7±0.2 | 1.2±0.1 | 0.7±0.1 | 0.7±0.1 | 0.8±0.3 | 1.3±0.3 | 3.0±0.9 | 1.3±0.3 | 0.8±0.3 | 5.1±1.2 | 7.2±0.9 | 9.9±1.3 | 2.4±0.3 | 6.0±1.2 | 12.2±0.4 |
| β-cubebene* | 1391 | 3.9±0.5 | 3.3±0.5 | 14.2±1.0 | 13.8±1.3 | 32.5±8.7 | 21.0±3.6 | 10.4±2.0 | 4.5±1.1 | 11.0±0.9 | 5.7±1.1 | 13.2±0.6 | 16.4±0.5 | 9.6±2.0 | 16.7±0.4 | 17.7±0.4 |
| β-elemene* | 1393 | 34.9±0.9 | 34.4±0.4 | 3.1±0.4 | 2.5±0.3 | 3.4±0.5 | 4.4±0.7 | 5.5±0.5 | 3.9±0.6 | 3.5±0.3 | 7.3±0.6 | 4.5±1.2 | 6.9±0.8 | 6.7±1.8 | 8.2±1.0 | 7.2±0.4 |
| β-caryophyllene | 1422 | 92.7±18.5 | 78.4±16.4 | 71.4±6.8 | 54.3±2.7 | 57.2±4.7 | 95.5±6.6 | 71.4±2.9 | 80.3±14.9 | 82.8±9.0 | 146.8±10.8 | 178.2±14.9 | 186.3±11.7 | 156.4±30.1 | 198.5±14.3 | 175.6±0.4 |
| β-copaene* | 1431 | 6.6±0.6 | 6.2±0.5 | 6.0±0.5 | 5.0±0.2 | 17.3±11.6 | 10.4±1.5 | 7.6±0.8 | 5.9±0.6 | 7.1±1.0 | 9.4±1.0 | 14.7±4.5 | 17.2±3.9 | 13.2±2.4 | 21.6±4.9 | 14.3±0.4 |
| γ-elemene* | 1440 | 0.5±0.1 | 0.4±0.1 | 0.2±0.1 | 0.2±0.1 | 0.4±0.1 | 1.7±0.3 | 3.4±1.1 | 1.0±0.5 | 0.7±0.3 | 0.8±0.3 | 2.3±0.2 | 4.4±0.6 | 2.4±0.4 | 4.4±0.6 | 5.9±0.4 |
| trans-muurola-3,5-diene* | 1451 | 2.2±0.6 | 3.3±0.7 | 6.4±0.8 | 5.9±0.8 | 4.4±0.9 | 7.4±1.0 | 8.3±1.6 | 3.6±0.8 | 4.9±0.2 | 5.3±1.0 | 2.9±0.6 | 4.4±0.9 | 2.8±0.4 | 3.8±1.1 | 6.3±0.3 |
| α-humulene | 1457 | 20.9±2.6 | 19.3±3.0 | 20.5±1.7 | 16.3±0.7 | 19.4±1.8 | 35.6±4.9 | 19.1±0.7 | 18.6±2.1 | 23.9±1.8 | 30.8±1.6 | 28.4±6.6 | 33.3±7.6 | 29.0±5.6 | 26.2±9.3 | 42.0±0.4 |
| allo-aromadendrene* | 1462 | 6.4±0.5 | 5.6±0.7 | 7.6±0.7 | 6.1±0.3 | 8.0±0.9 | 17.5±2.6 | 9.2±1.2 | 5.9±0.8 | 7.5±0.4 | 9.3±0.9 | 15.8±3.2 | 18.7±2.6 | 10.2±2.2 | 21.8±3.3 | 17.1±0.4 |
| cis-muurola-4(14),5-diene* | 1465 | 3.8±0.3 | 3.6±0.3 | 3.7±0.3 | 3.5±0.7 | 3.9±0.6 | 5.9±0.9 | 5.3±0.7 | 3.5±0.3 | 4.6±0.4 | 5.4±0.6 | 7.0±2.4 | 9.7±1.8 | 6.6±1.1 | 12.0±2.3 | 8.8±0.4 |
| trans-cadina-1(6),4-diene* | 1476 | 2.7±0.3 | 2.8±0.5 | 6.8±0.5 | 5.0±0.3 | 5.5±0.6 | 3.8±0.7 | 5.9±1.3 | 3.2±0.7 | 5.0±0.6 | 3.2±0.7 | 12.4±1.2 | 15.4±0.6 | 5.7±1.3 | 16.4±0.7 | 15.9±0.4 |
| γ-muurolene* | 1479 | 32.5±1.8 | 29.7±3.1 | 37.3±1.9 | 30.8±1.0 | 34.7±4.0 | 62.5±6.7 | 36.7±1.9 | 28.6±2.4 | 29.9±2.9 | 37.7±2.4 | 59.7±5.4 | 62.6±4.4 | 40.7±5.9 | 67.3±5.4 | 58.9±0.2 |
| germacrene D* | 1484 | 23.5±3.3 | 21.5±2.0 | 16.4±0.8 | 13.9±0.5 | 14.1±0.8 | 19.0±1.1 | 19.3±2.4 | 19.0±2.4 | 24.3±6.8 | 37.6±5.0 | 33.7±14.7 | 36.9±13.9 | 44.6±9.3 | 51.4±17.1 | 24.0±0.4 |
| β-selinene* | 1489 | 7.5±0.8 | 7.1±0.5 | 5.7±0.7 | 5.4±0.2 | 8.4±0.8 | 10.7±1.2 | 8.9±0.7 | 6.1±0.6 | 6.0±0.3 | 9.7±1.1 | 8.7±2.6 | 11.3±2.1 | 10.0±1.6 | 13.8±2.6 | 10.2±0.4 |
| trans-muurola-1(14),5-diene* | 1494 | 5.5±0.4 | 5.8±0.9 | 13.6±0.8 | 10.6±0.5 | 13.1±0.9 | 14.7±2.9 | 6.8±0.5 | 4.7±0.2 | 10.5±0.8 | 7.6±1.0 | 15.3±1.4 | 18.3±0.8 | 9.7±2.1 | 19.5±0.9 | 18.6±0.4 |
| α-muurolene* | 1503 | 16.1±1.6 | 17.1±2.3 | 27.6±2.3 | 21.9±1.4 | 25.7±2.9 | 46.8±5.9 | 24.7±1.9 | 16.4±2.5 | 26.1±1.2 | 24.3±2.1 | 42.9±5.2 | 55.7±0.9 | 24.6±5.2 | 57.1±1.1 | 55.9±0.4 |
| β-bisabolene* | 1510 | 22.1±2.3 | 21.6±2.5 | 81.9±5.4 | 62.5±5.3 | 74.3±8.2 | 105.0±19.9 | 38.5±9.2 | 21.4±2.3 | 63.6±4.0 | 36.6±4.8 | 55.3±5.9 | 60.0±4.5 | 42.7±8.6 | 65.1±5.6 | 56.5±0.4 |
| γ-cadinene* | 1517 | 15.7±1.2 | 15.3±1.0 | 12.6±1.3 | 11.7±0.8 | 23.7±9.9 | 32.8±2.9 | 35.4±8.0 | 20.2±6.6 | 15.3±0.7 | 20.5±2.0 | 24.9±8.3 | 27.0±7.9 | 19.9±3.4 | 35.4±9.7 | 20.2±0.4 |
| δ-cadinene* | 1528 | 60.3±5.5 | 59.0±9.3 | 112.5±9.5 | 83.3±3.8 | 98.4±9.1 | 176.2±19.6 | 88.4±8.5 | 59.7±10.7 | 98.6±4.8 | 83.9±8.0 | 143.6±19.0 | 153.9±16.3 | 87.4±18.2 | 170.7±20.0 | 138.6±0.4 |
| cadina-1,4-diene* | 1535 | 7.1±0.8 | 5.4±0.6 | 6.3±0.5 | 6.1±0.6 | 9.8±1.2 | 11.2±0.5 | 10.4±2.0 | 5.2±1.6 | 5.0±0.5 | 3.9±2.3 | 7.1±0.5 | 8.4±0.4 | 7.1±0.5 | 8.8±0.3 | 9.5±0.4 |
| α-cadinene* | 1541 | 4.9±0.4 | 4.1±0.3 | 14.1±0.4 | 13.1±0.1 | 5.1±0.5 | 10.4±1.7 | 14.6±4.0 | 7.0±3.0 | 5.1±0.5 | 5.4±0.8 | 6.6±1.6 | 8.2±1.3 | 5.0±0.8 | 10.0±1.6 | 8.0±0.4 |
| α-calacorene* | 1546 | 1.2±0.1 | 1.2±0.0 | 1.2±0.1 | 1.3±0.0 | 1.3±0.1 | 7.9±0.5 | 8.2±0.4 | 8.7±0.8 | 3.5±0.4 | 4.1±0.2 | 1.7±0.2 | 1.3±0.0 | 1.2±0.1 | 1.3±0.0 | 1.3±0.1 |
| cubebol* | 1520 | 15.8±3.5 | 15.5±3.0 | 112.9±12.0 | 75.5±20.6 | 108.9±16.4 | 117.6±21.7 | 60.3±27.8 | 17.9±4.9 | 71.4±5.0 | 36.6±7.8 | 57.8±2.5 | 62.3±1.6 | 42.4±9.3 | 61.3±1.8 | 64.8±0.4 |
| elemol* | 1553 | 8.7±2.0 | 8.4±1.4 | 7.5±1.2 | 6.3±1.0 | 9.4±1.8 | 6.7±0.7 | 10.3±1.5 | 5.9±1.2 | 7.8±0.3 | 11.5±0.8 | 9.6±1.5 | 13.1±0.7 | 9.6±1.9 | 14.3±0.9 | 13.5±0.4 |
| germacrene D-4-ol* | 1584 | 1.2±0.0 | 1.2±0.0 | 1.1±0.1 | 1.2±0.1 | 1.1±0.1 | 1.8±0.3 | 3.6±1.0 | 1.6±0.4 | 1.1±0.1 | 1.0±0.3 | 1.1±0.0 | 1.1±0.1 | 1.2±0.0 | 1.1±0.1 | 1.6±0.1 |
| spathulenol* | 1584 | 13.0±2.8 | 11.8±2.2 | 9.7±1.1 | 11.9±0.5 | 18.5±4.4 | 10.1±0.9 | 13.2±1.6 | 10.8±2.6 | 7.6±0.7 | 40.6±18.0 | 13.1±1.2 | 26.7±11.0 | 22.4±3.6 | 28.0±10.7 | 38.2±13.5 |
| caryophyllene oxide | 1587 | 56.0±18.3 | 52.0±15.9 | 53.0±8.6 | 45.6±6.4 | 67.7±11.6 | 54.8±2.2 | 53.3±2.1 | 55.1±14.3 | 48.1±1.7 | 71.8±6.6 | 58.6±2.6 | 70.4±10.2 | 49.1±9.0 | 67.9±11.0 | 83.6±10.9 |
| gleenol* | 1588 | 1.2±0.0 | 1.2±0.0 | 1.1±0.1 | 1.2±0.1 | 1.1±0.1 | 9.8±0.3 | 11.7±1.0 | 3.2±2.0 | 1.1±0.1 | 11.2±4.1 | 1.1±0.0 | 1.1±0.1 | 1.2±0.0 | 1.1±0.1 | 1.6±0.1 |
| humulene oxide II | 1613 | 12.3±2.0 | 11.5±1.7 | 16.8±1.9 | 15.2±1.5 | 24.3±3.0 | 22.6±3.2 | 13.5±1.2 | 11.1±1.5 | 16.0±1.2 | 14.7±1.5 | 17.6±2.4 | 21.3±2.3 | 12.7±2.6 | 23.7±2.2 | 22.4±2.1 |
| 1-epi-cubenol* | 1632 | 6.0±3.3 | 5.9±2.9 | 16.3±1.3 | 12.3±1.3 | 10.4±0.9 | 14.1±2.2 | 10.9±1.7 | 6.8±2.4 | 12.5±1.8 | 11.6±2.5 | 9.9±0.8 | 12.4±0.5 | 4.0±1.1 | 13.0±0.4 | 13.4±0.5 |
| α-muurolol* | 1648 | 11.5±2.3 | 11.7±2.1 | 26.9±1.5 | 23.0±1.6 | 20.3±5.1 | 19.0±2.5 | 14.8±1.2 | 10.4±2.1 | 21.6±1.2 | 12.3±2.4 | 15.9±3.6 | 21.3±3.8 | 13.8±2.8 | 24.7±3.8 | 22.2±3.8 |
| α-cadinol* | 1661 | 15.1±2.8 | 17.4±1.9 | 22.7±0.8 | 21.4±1.0 | 24.6±1.3 | 21.0±1.4 | 29.6±0.9 | 18.2±3.2 | 24.8±0.7 | 19.5±2.8 | 17.9±5.6 | 24.0±5.7 | 18.6±2.3 | 29.4±5.9 | 23.9±5.0 |
| α-bisabolol* | 1688 | 9.9±0.6 | 9.8±0.5 | 10.7±0.5 | 10.3±0.4 | 10.3±0.5 | 32.8±0.9 | 34.0±2.0 | 15.5±5.8 | 27.3±0.8 | 15.1±6.9 | 4.4±0.3 | 5.8±0.4 | 5.8±0.5 | 7.6±0.6 | 7.2±0.2 |
| eudesma-4(15),7-dien-1-beta-ol* | 1696 | 10.5±2.9 | 6.9±0.7 | 22.8±2.6 | 19.5±1.5 | 27.5±3.4 | 26.0±0.5 | 35.3±2.5 | 21.3±3.1 | 11.3±3.4 | 27.5±2.1 | 8.8±1.0 | 11.9±1.1 | 7.5±1.1 | 11.4±1.2 | 13.8±0.5 |
| Totalmonoterpenes | 39±3.9 | 65±5.0 | 299±6.8 | |||||||||||||
| Total monoterpene hydrocarbons | 21.7±3.4 | 37.5±3.2 | 221.5±5.3 | |||||||||||||
| Total oxygenated monoterpenes | 16.8±1.9 | 27.8±2.4 | 77.1±2.5 | |||||||||||||
| Total sesquiterpenes | 799±38.0 | 956±48.6 | 1181±55.0 | |||||||||||||
| Total sesquiterpene hydrocarbons | 422.18±18.0 | 509.1±32.4 | 762.5±37.1 | |||||||||||||
| Total oxygenated sesquiterpenes | 377.2±22.7 | 446.9±20.7 | 418.9±23.3 | |||||||||||||
| Total terpenes | 838±39.0 | 1021±51.6 | 1480±57.7 | |||||||||||||
| Terpenoids | factors | df | SS | MS | F | P |
|---|---|---|---|---|---|---|
| Monoterpene hydrocarbons | Site | 2 | 617137 | 308569 | 479.144 | < 0.001 |
| Site:Station | 12 | 7725 | 644 | 1.754 | 0.08 | |
| Residuals | 60 | 22007 | 367 | |||
| Oxygenated monoterpenes | Site | 2 | 51551 | 25775.3 | 108.802 | < 0.001 |
| Site:Station | 12 | 2843 | 236.9 | 2.823 | 0.004 | |
| Residuals | 60 | 5037 | 83.9 | |||
| Sesquiterpene hydrocarbons | Site | 2 | 1563493 | 781746 | 11.700 | 0.001 |
| Site:Station | 12 | 801748 | 66812 | 4.718 | < 0.001 | |
| Residuals | 60 | 849581 | 14160 | |||
| Oxygenated sesquiterpenes | Site | 2 | 61397 | 30698 | 0.688 | 0.52 |
| Site:Station | 12 | 535103 | 44592 | 7.484 | < 0.001 | |
| Residuals | 60 | 357500 | 5958 | |||
| Total terpenes | Site | 2 | 5470660 | 2735330 | 13.782 | < 0.001 |
| Site:Station | 12 | 2381658 | 1984472 | 5.601 | < 0.001 | |
| Residuals | 60 | 2126026 | 35434 |

3. Experimental
3.1. Plant material
3.2. Terpenoids extraction
| Sites | Stations | Alt.(m) | Geographical coordinates | Habitat | P (mm) | T0 Max (C0) | Q2 |
|---|---|---|---|---|---|---|---|
| Low-altitude | Cherchel (L1) | 15 | 36°36’N/2°11’E | Phillyrea latifolia | 502.24±25.08 | 31.75±0.42 | 73.54 |
| Azzefoun (L2) | 14 | 36°53’N/4°25’E | Phillyrea latifolia | 775.78±50.04 | 31.50±0.48 | 122.06 | |
| Zemmouri Bahri (L5) | 9 | 36°48’N/3°35’E | Erica arborea | 741.14±45.34 | 29.10±0.51 | 125.85 | |
| Tigzirt (L3) | 6 | 36°53’N/4°08’E | Arbutus unedo | 923.09±67.98 | 30.30±0.44 | 150.77 | |
| Ziama Mansouriah (L4) | 8 | 36°39’N/5°28’E | Erica arborea | 968.11±60.82 | 30.80±0.38 | 148.24 | |
| Mid-altitude | Thenia (M1) | 168 | 36°43’N/3°32’E | Eucalyptus radiata | 728.53±58.43 | 35.00±0.37 | 88.61 |
| Beni Amrane (M4) | 121 | 36°39’N/5°14’E | Erica arborea | 729.58±55.22 | 35.50±0.38 | 87.50 | |
| Tizi Ouzou (M5) | 182 | 36°43’N/4°03’E | Pinus halepensis | 731.92±57.42 | 36.10±0.41 | 83.68 | |
| El Kseur (M3) | 167 | 36°41’N/4°51’E | Olea europea | 758.39±59.33 | 30.80±0.35 | 112.12 | |
| Lakhdaria (M2) | 307 | 36°34’N/3°35’E | Olea europea | 513.85±54.14 | 35.60±0.39 | 55.25 | |
| High- altitude | Ait Khelifa (H3) | 1042 | 36°31’N/4°20’E | Olea europea | 1039.30±56.57 | 23.20±0.32 | 168.95 |
| Tala Moumene (H4) | 1008 | 36°33’N/4°48’E | Olea europea | 1041.27±67.45 | 24.46±0.35 | 190.18 | |
| Beni Mendes (H1) | 809 | 36°30’N/3°59’E | Olea europea | 924.56±64.54 | 31.75±0.38 | 112.74 | |
| Aourir N’Athjelil (H2) | 868 | 36°34’N/4°48’E | Olea europea | 1019.62±69.33 | 25.44±0.31 | 182.15 | |
| Tizi-Oumalou (H5) | 942 | 36°30’N/4°20’E | Olea europea | 939.06±78.75 | 30.82±0.36 | 116.11 |
3.3. Quantitative and qualitative analysis of terpenoids
3.4. Statistical analyses
4. Conclusions
Acknowledgements
References and Notes
- Naghdi Badi, H.; Yazdani, D.; Mohammad, A.S.; Nazari, F. Effects of spacing and harvesting on herbage yield and quality/quantity of oil in thyme, Thymus vulgaris L. Ind. Crop. Prod. 2004, 19, 231–236. [Google Scholar] [CrossRef]
- Yu, F.N.A.; Utsumi, R. Diversity, regulation, and genetic manipulation of plant mono-and sesquiterpenoid biosynthesis. Cell. Mol. Life Sci. 2009, 66, 3043–3052. [Google Scholar] [CrossRef]
- Kofidis, G.; Bosabalidis, A.; Moustakas, M. Chemical composition of essential oils from leaves and twigs of Pistacia lentiscus, Pistacia lentiscus var. chia, and Pistacia terebinthus from Turkey. Pharm. Biol. 2003, 42, 360–366. [Google Scholar]
- Tkachev, A.V.; Korolyuk, E.A.; Letchamo, W. Chemical screening of volatile oil- bearing flora of Siberia IX. Variations in chemical composition of the essential oil of Heteropappus altaicus Willd. (Novopokr.) growing wild at different altitudes of Altai Region, Russia. J. Essent. Oil Res. 2006, 18, 149–151. [Google Scholar] [CrossRef]
- Agnihotri, V.K.; Lattoo, S.K.; Thappa, R.K.; Kaul, P.; Qazi, G.N.; Dhar, A.K.; Saraf, A.; Kapahi, B.K.; Saxena, R.K.; Agarwal, S.G. Chemical variability in the essential oil components of Achillea millefolium Agg. from different Himalayan habitats (India). Planta Med. 2005, 71, 280–283. [Google Scholar] [CrossRef]
- Munne-Bosch, S.; Penuelas, J. Photo- and antioxidative protection during summer leaf senescence in Pistacia lentiscus L. grown under Mediterranean field conditions. Ann. Bot. 2003, 92, 385–391. [Google Scholar] [CrossRef]
- Castro-Diez, P.; Villar-Salvador, P.; Perez-Rontome, C.; Maestro-Martinez, M.; Montserrat-Marti, G. Leaf morphology, leaf chemical composition and stem xylem characteristics in two Pistacia (Anacardiaceae) species along a climatic gradient. Flora 1998, 193, 195–202. [Google Scholar]
- Ozturk, M.; Dogan, Y.; Sakcali, M.S.; Doulis, A.; Karam, F. Ecophysiological responses of some maquis (Ceratonia siliqua L., Olea oleaster Hoffm. & Link, Pistacia lentiscus and Quercus coccifera L.) plant species to drought in the east Mediterranean ecosystem. J. Environ. Biol. 2010, 31, 233–245. [Google Scholar]
- Barazani, O.; Golan-Goldhirsh, A. Salt-driven interactions between Pistacia lentiscus and Salsola inermis. Environ. Sci. Poll. Res. 2009, 16, 855–861. [Google Scholar] [CrossRef]
- Verdu, M.; Garcia-Fayos, P. Reproductive ecology of Pistacia lentiscus L. (Anacardiaceae): an evolutionary anachronism in the Mediterranean shrubland. Rev. Chil. Hist. Nat. 2002, 75, 57–65. [Google Scholar]
- Charef, M.; Yousfi, M.; Saidi, M.; Stocker, P. Determination of the fatty acid composition of acorn (Quercus), Pistacia lentiscus seeds growing in Algeria. J. Am. Oil Chem. Soc. 2008, 85, 921–924. [Google Scholar] [CrossRef]
- Janakat, S.; Al-Merie, H. Evaluation of hepatoprotective effect of Pistacia lentiscus, Phillyrea latifolia and Nicotiana glauca. J. Ethnopharmacol. 2002, 83, 135–138. [Google Scholar] [CrossRef]
- Benhammou, N.; Bekkara, F.A.; Panovska, T.K. Antioxidant and antimicrobial activities of the Pistacia lentiscus and Pistacia atlantica extracts. Afr. J. Pharm. Pharmacol. 2008, 2, 022–028. [Google Scholar]
- Atmani, D.; Chaher, N.; Berboucha, M.; Ayouni, K.; Lounis, H.; Boudaoud, H.; Debbache, N. Antioxidant capacity and phenol content of selected Algerian medicinal plants. Food Chem. 2009, 112, 303–309. [Google Scholar] [CrossRef]
- Fahn, A. Secretory Tissues in Plants; Academic Press: London, UK, 1979. [Google Scholar]
- Llusià, J.; Peñuelas, J. Changes in terpene content and emission in potted Mediterranean woody plants under severe drought. Can. J. Bot. Rev. Can. Bot. 1998, 76, 1366–1373. [Google Scholar]
- Zrira, S.; Elamrani, A.; Benjilali, B. Chemical composition of the essential oil of Pistacia lentiscus L. from Morocco - a seasonal variation. Flavour Fragr. J. 2003, 18, 475–480. [Google Scholar] [CrossRef]
- Duru, M.E.; Cakir, A.; Kordali, S.; Zengin, H.; Harmandar, M.; Izumi, S.; Hirata, T. Chemical composition and antifungal properties of essential oils of three Pistacia species. Fitoterapia 2003, 74, 170–176. [Google Scholar] [CrossRef]
- Barra, A.; Coroneo, V.; Dessi, S.; Cabras, P.; Angioni, A. Characterization of the volatile constituents in the essential oil of Pistacia lentiscus L. from different origins and its antifungal and antioxidant activity. J. Agr. Food Chem. 2007, 55, 7093–7098. [Google Scholar] [CrossRef]
- Gardeli, C.; Vassiliki, P.; Athanasios, M.; Kibouris, T.; Komaitis, M. Essential oil composition of Pistacia lentiscus L. and Myrtus communis L.: Evaluation of antioxidant capacity of methanolic extracts. Food Chem. 2008, 107, 1120–1130. [Google Scholar] [CrossRef]
- Fernandez, A.; Camacho, A.; Fernandez, C.; Altarejos, A.; Perez, P. Composition of the essential oils from galls and aerial parts of Pistacia lentiscus L. J. Essent. Oil Res. 2000, 12, 19–23. [Google Scholar] [CrossRef]
- Tzakou, O.; Bazos, I.; Yannitsaros, A. Volatile metabolites of Pistacia atlantica Desf. from Greece. Flavour Fragr. J. 2007, 22, 358–362. [Google Scholar] [CrossRef]
- Haider, F.; Kumar, N.; Banerjee, S.; Naqvi, A.A.; Bagchi, G.D. Effect of Altitude on the essential Oil Constituents of Artemisia roxburghiana Besser var. purpurascens (Jacq.) Hook. J. Essent. Oil Res. 2009, 21, 303–304. [Google Scholar] [CrossRef]
- Vokou, D.; Kokkini, S.; Bessiere, J.M. Geographic-Variation of Greek Oregano (Origanum-Vulgare ssp Hirtum) Essential Oils. Biochem. Syst. Ecol. 1993, 21, 287–295. [Google Scholar] [CrossRef]
- Bilger, W.; Rolland, M.; Nybakken, L. UV screening in higher plants induced by low temperature in the absence of UV-B radiation. Photochem. Photobiol. Sci. 2007, 6, 190–195. [Google Scholar] [CrossRef]
- Congiu, R.; Falconieri, D.; Marongiu, B.; Piras, A.; Porcedda, S. Extraction and isolation of Pistacia lentiscus L. essential oil by supercritical CO2. Flavour Fragr. J. 2002, 17, 239–244. [Google Scholar] [CrossRef]
- Mecherara-Idjeri, S.; Hassani, A.; Castola, V.; Casanova, J. Composition and Chemical Variability of the Essential oil from Pistacia lentiscus L. Growing Wild in Algeria Part: Leaf Oil. J. Essent. Oil Res. 2008, 20, 32–38. [Google Scholar] [CrossRef]
- Tiwari, M.; Kakkar, P. Plant derived antioxidants - Geraniol and camphene protect rat alveolar macrophages against t-BHP induced oxidative stress. Toxicol. In Vitro 2009, 23, 295–301. [Google Scholar] [CrossRef]
- Hudaib, M.; Aburjai, T. Volatile components of Thymus vulgaris L. from wild-growing and cultivated plants in Jordan. Flavour Fragr. J. 2007, 22, 322–327. [Google Scholar] [CrossRef]
- Adams, R.P. Identification of Essential Oil Components by Gas Chromatography/Mass Spectrometry, 4th ed; Allured Publishing Corporation: Carol Stream, IL, USA, 2007. [Google Scholar]
- R Development Core Team (2009). R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2009. ISNB 3-900051-07-0. Available online: http://www.R-project.org.
- Dray, S.; Dufour, A.B.; Chessel, D. The ade4 package-II: Two-table and K-table methods. R News 2007, 7, 47–52. [Google Scholar]
- De Mendiburu, F. Agricolae: Statistical Procedures for Agricultural Research, R package version 1.0-9. Available online: http://CRAN.R-project.org/package=agricolae,accessed on 24 February 2010.
- Sample Availability: Samples of the compounds are available from the authors.
© 2011 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license ( http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Said, S.A.; Fernandez, C.; Greff, S.; Torre, F.; Derridj, A.; Gauquelin, T.; Mevy, J.-P. Inter-Population Variability of Terpenoid Composition in Leaves of Pistacia lentiscus L. from Algeria: A Chemoecological Approach. Molecules 2011, 16, 2646-2657. https://doi.org/10.3390/molecules16032646
Said SA, Fernandez C, Greff S, Torre F, Derridj A, Gauquelin T, Mevy J-P. Inter-Population Variability of Terpenoid Composition in Leaves of Pistacia lentiscus L. from Algeria: A Chemoecological Approach. Molecules. 2011; 16(3):2646-2657. https://doi.org/10.3390/molecules16032646
Chicago/Turabian StyleSaid, Samir Ait, Catherine Fernandez, Stéphane Greff, Franck Torre, Arezki Derridj, Thierry Gauquelin, and Jean-Philippe Mevy. 2011. "Inter-Population Variability of Terpenoid Composition in Leaves of Pistacia lentiscus L. from Algeria: A Chemoecological Approach" Molecules 16, no. 3: 2646-2657. https://doi.org/10.3390/molecules16032646