Effects of Andrographis paniculata and Orthosiphon stamineus Extracts on the Glucuronidation of 4-Methylumbelliferone in Human UGT Isoforms

Abstract
:1. Introduction
2. Results and Discussion





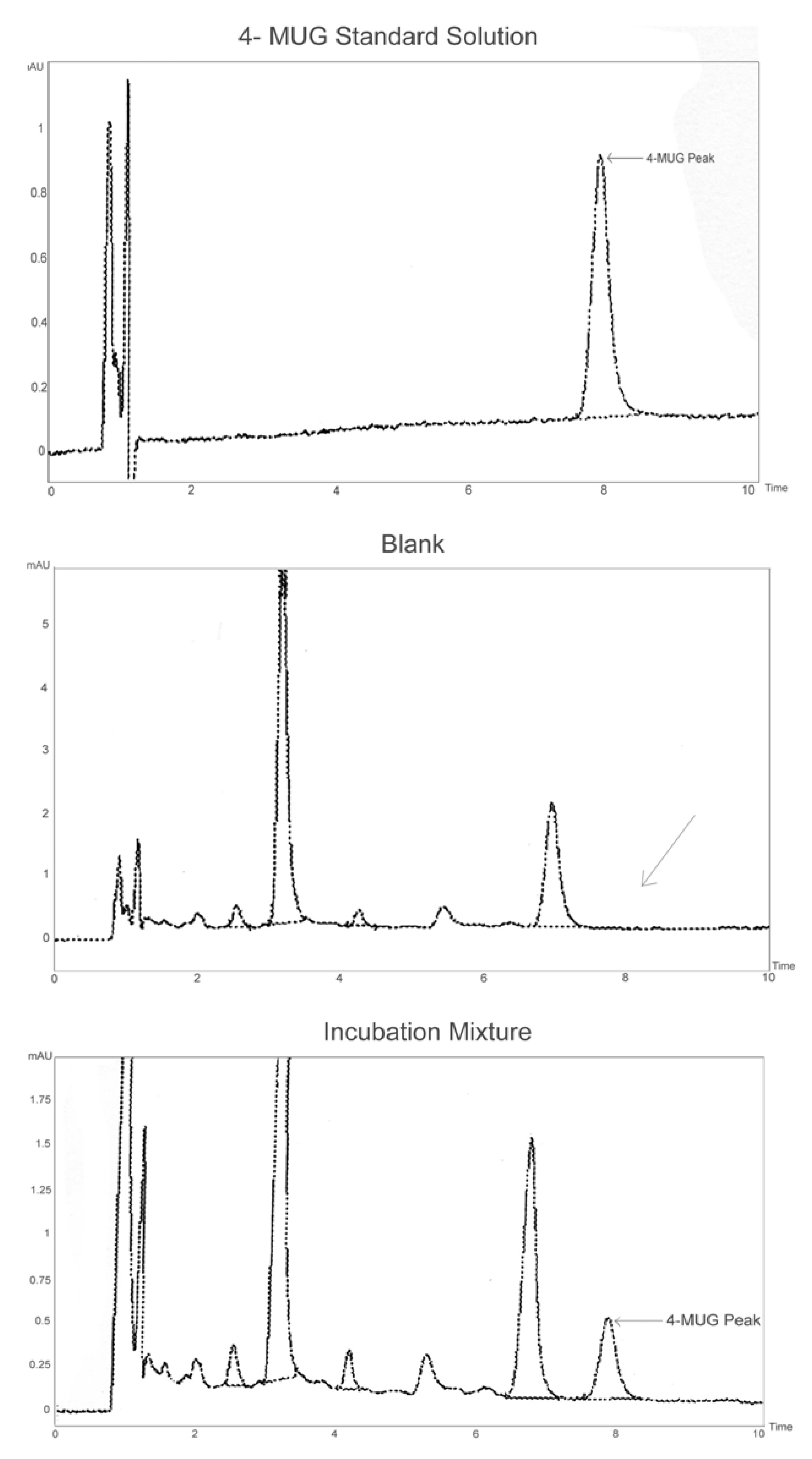{kind=link}
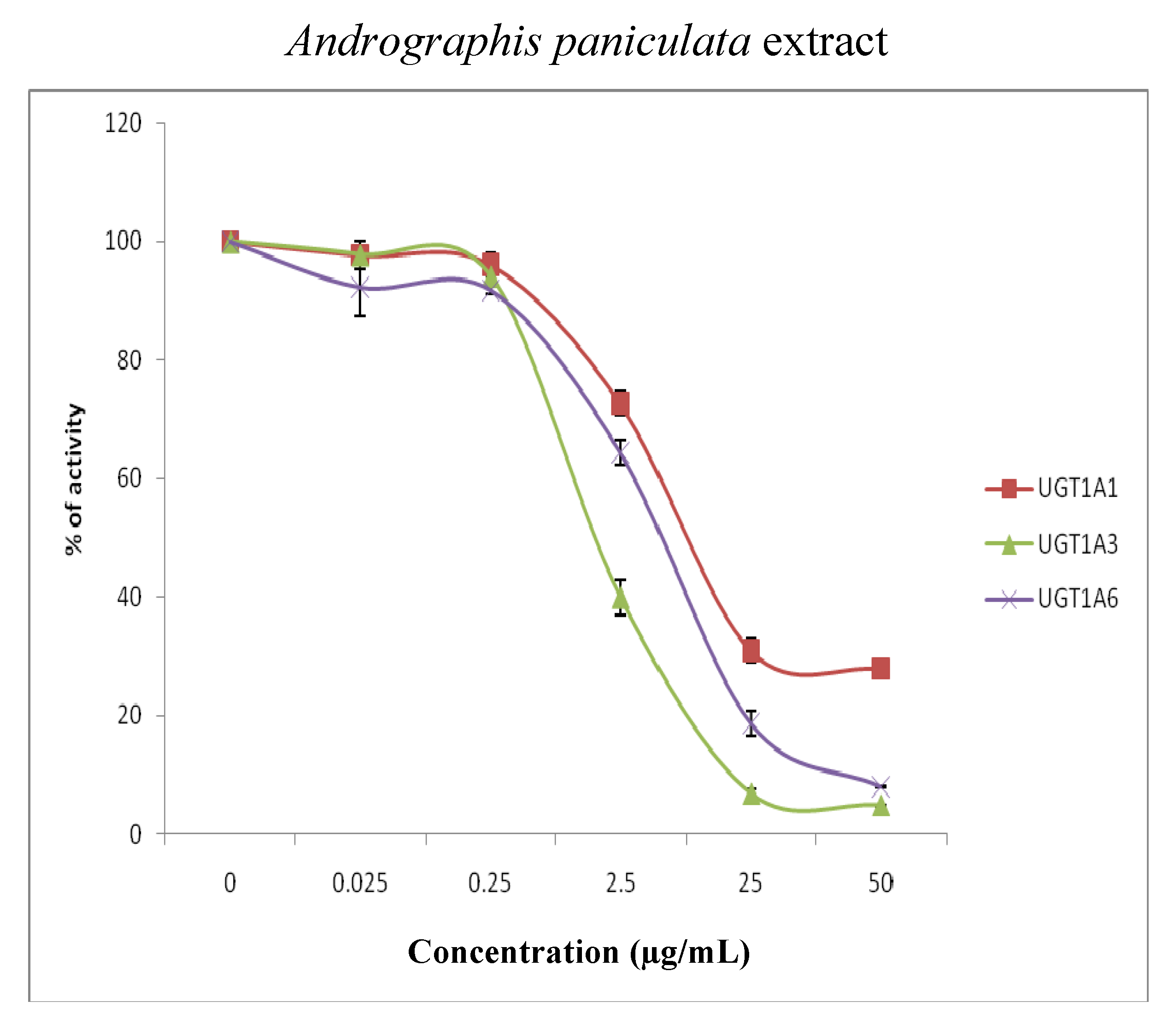{kind=link}
{kind=link}
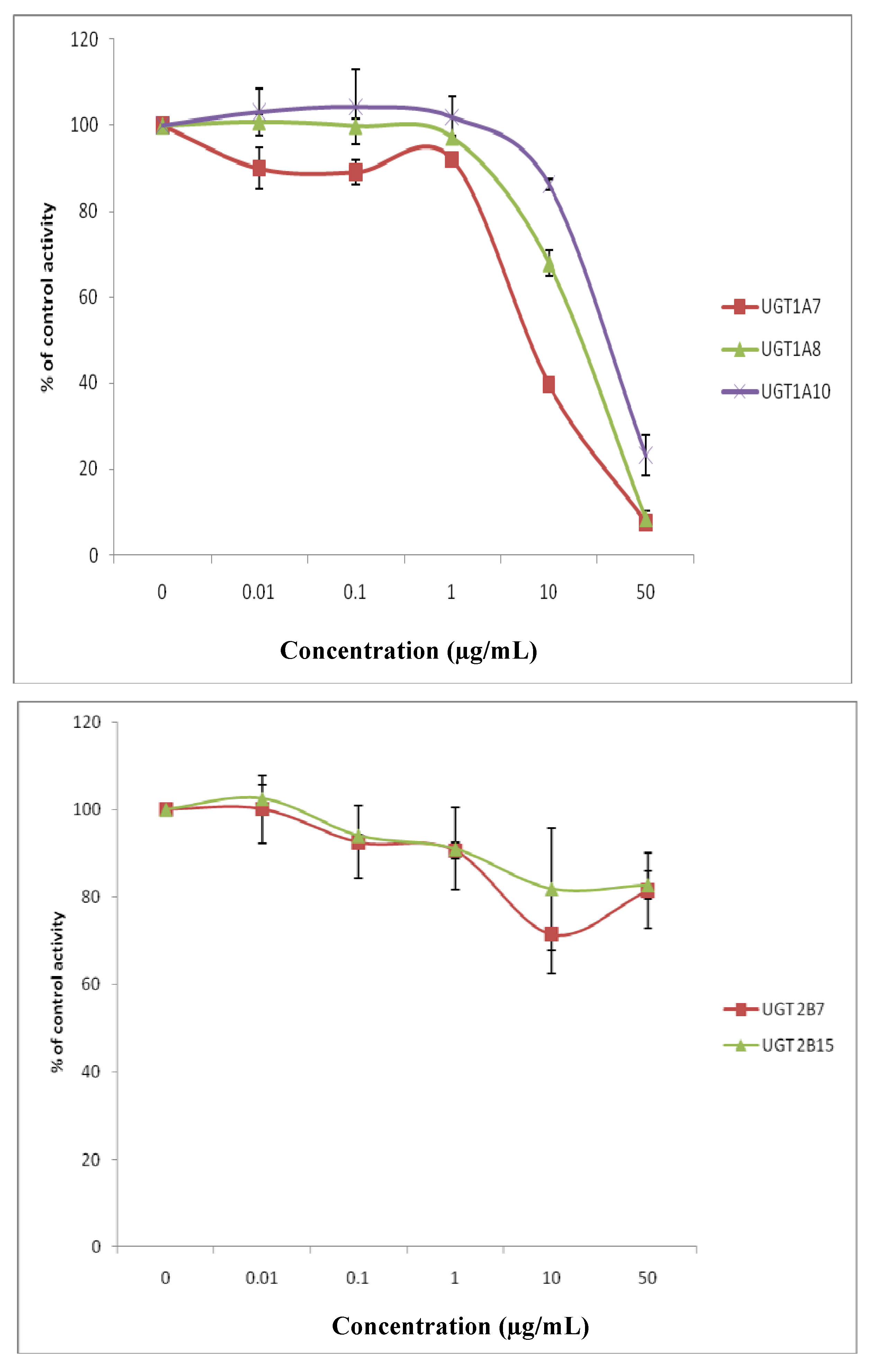{kind=link}
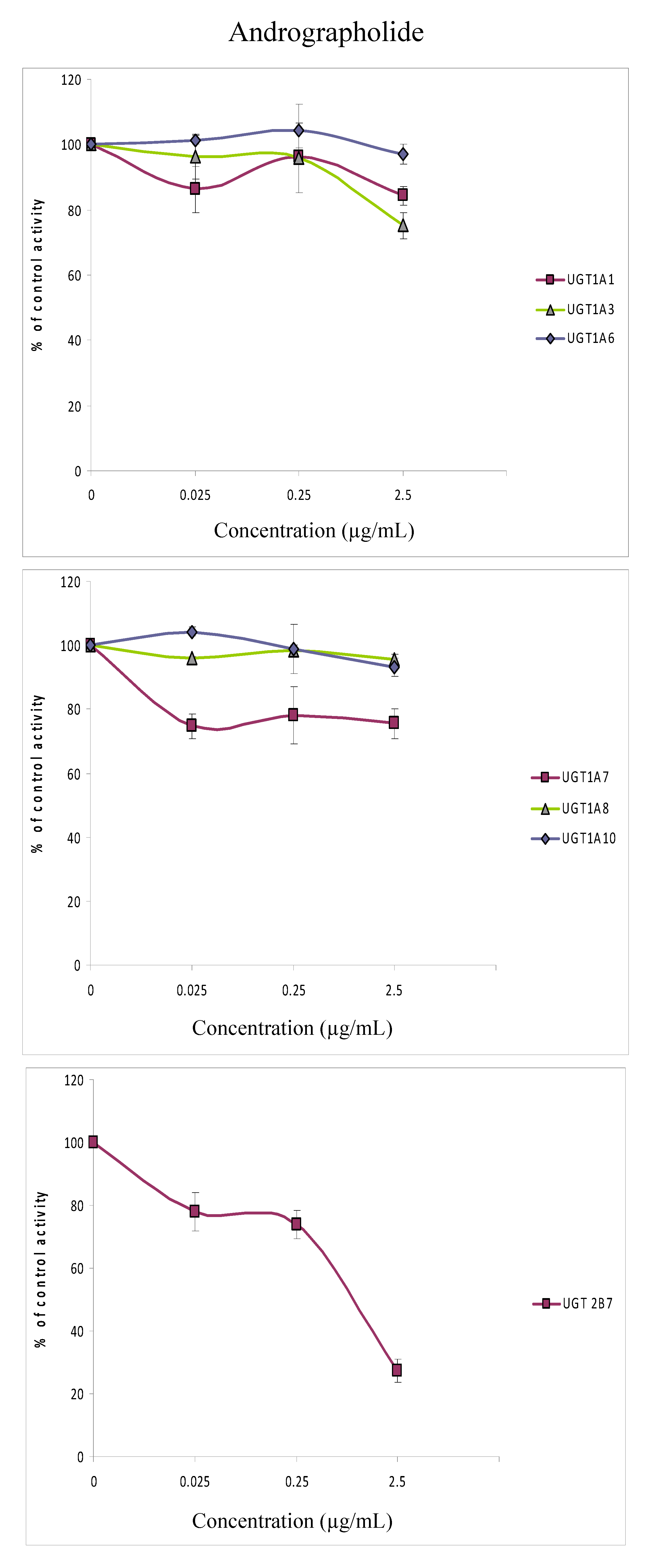{kind=link}
| Isoforms | Inhibitors (µg/mL) | |
|---|---|---|
| Andrographis paniculata | Orthosiphon stamineus | |
| UGT1A1 | 5.00 | 24.65 |
| UGT1A3 | 1.70 | >50 |
| UGT1A6 | 5.66 | 30.02 |
| UGT1A7 | 9.88 | 10.83 |
| UGT1A8 | 2.57 | 43.39 |
| UGT1A10 | 15.66 | >50 |
| UGT2B7 | 2.82 | >50 |
| UGT2B15 | >50 | >50 |

3. Experimental
3.1. Materials
3.2. Preparation of Andrographis paniculata extract
3.3. HPLC analysis for the determination of andrographolide and neoandrographolide percentages in Andrographis paniculata extract
3.4. Preparation of Orthosiphon stamineus extract
3.5. HPLC analysis for the determination of sinensetin, eupatorin, 3’-hydroxy-5,6,7,4’-tetramethoxy-flavone and rosmarinic acid percentages in Orthosiphon stamineus extract
3.6. Expression of UGT proteins
3.7. 4MU glucuronidation assays
3.8. Inhibition of 4MU glucuronidation by Andrographis paniculata and Orthosiphon stamineus extracts
3.9. Inhibition of 4MU glucuronidation by andrographolide
| Isoform | 4MU | ||
|---|---|---|---|
| Protein amount | Incubation time | Concentration | |
| µg/incubation | min | µM | |
| 1A1 | 67 | 120 | 100 |
| 1A3 | 33 | 75 | 1000 |
| 1A6 | 0.5 | 30 | 100 |
| 1A7 | 1.67 | 10 | 15 |
| 1A8 | 10 | 30 | 750 |
| 1A10 | 10 | 30 | 30 |
| 2B7 | 50 | 120 | 350 |
| 2B15 | 167 | 120 | 300 |
4. Conclusions
Acknowledgements
References
- Bensky, D.; Gamble, A. Chinese Herbal Medicine Materia Medica; Eastland Press: Seattle, WA, USA, 1993; p. 95, revised edition. [Google Scholar]
- Caceres, D.D.; Hancke, J.L.; Burgos, R.A.; Wikman, G.K. Prevention of common colds with Andrographis paniculata dried extract: a pilot double-blind trial. Phytomedicine 1997, 4, 101–104. [Google Scholar] [CrossRef]
- Puri, A.; Saxena, R.; Saxena, R.P.; Saxena, K.C.; Srivastava, V.; Tandon, J.S. Immunostimulant agents from Andrographis paniculata. J. Nat. Prod. 1993, 56, 995–999. [Google Scholar] [CrossRef]
- Chang, R.S.; Yeung, H.W. Inhibition of growth of human immunodeficiency virus in vitro by crude extracts of Chinese medicinal herbs. Antivir. Res. 1988, 9, 163–175. [Google Scholar]
- Choudhury, B.R.; Haque, S.J.; Poddar, M.K. In vivo and in vitro effects of Kalmegh (Andrographis paniculata) extract and andrographolide on hepatic microsomal drug metabolizing enzymes. Planta Med. 1987, 53, 135–140. [Google Scholar] [CrossRef]
- Singh, R.P.; Banerjee, S.; Rao, A.R. Modulatory influence of Andrographis paniculata on mouse hepatic and extrahepatic carcinogen metabolizing enzymes and antioxidant status. Phytother. Res. 2001, 15, 382–390. [Google Scholar] [CrossRef]
- Jarukamjorn, K.; Don-in, K.; Makejaruskul, C.; Laha, T.; Daodee, S.; Pearaksa, P.; Sripanidkulchai, B. Impact of Andrographis paniculata crude extract on mouse hepatic cytochrome P450 enzymes. J. Ethnopharmacol. 2006, 24, 464–467. [Google Scholar]
- Pekthong, D.; Martin, H.; Abadie, C.; Bonet, A.; Heyd, B.; Mantion, G.; Richert, L. Differential inhibition of rat and human hepatic cytochrome P450 by Andrographis paniculata extract and andrographolide. J. Ethnopharmacol. 2008, 115, 432–440. [Google Scholar] [CrossRef]
- WHORegional Office for the Western Pacific ManilaInstitute of Material Medica HanoiMedicinal Plants in Vietnam; Science and Technology Publishing House: Hanoi, Vietnam, 1990; p. 271.
- Stampoulis, P.; Tezuka, Y.; Banskota, A.H.; Tran, K.Q.; Saiki, I.; Kadota, S.; Staminol, A. a novel diterpene from Orthosiphon Stamineus. Tetrahedron Lett. 1999, 40, 4239–4242. [Google Scholar]
- Akowuah, G.A.; Zhari, I.; Norhayati, I.; Sadikun, A.; Khamsah, S.M. Sinensetin, eupatorin, 3’-hydroxy-5, 6, 7, 4’-tetramethoxyflavone and rosmarinic acid contents and antioxidative effect of Orthosiphon stamineus from Malaysia. Food Chem. 2004, 87, 559–566. [Google Scholar] [CrossRef]
- Mackenzie, P.I.; Bock, K.W.; Burchell, B.; Guillemette, C.; Ikushiro, S.; Iyanagi, T.; Miners, J.O.; Owens, I.S.; Nebert, D.W. Nomenclature update for the mammalian UDP glycosyltransferase (UGT) gene superfamily. Pharmacogenet. Genomics 2005, 15, 677–685. [Google Scholar] [CrossRef]
- Fisher, M.B.; Paine, M.F.; Strelevitz, T.J.; Wrighton, S.A. The role of hepatic and extrahepatic UDP-glucuronosyltransferase in human drug metabolism. Drug Metab. Rev. 2001, 33, 273–297. [Google Scholar] [CrossRef]
- Uchaipichat, V.; Mackenzie, P.I.; Guo, X.H.; Gardner-Stephen, D.; Galetin, A.; Houston, J.B.; Miners, J.O. Human UDP-glucuronosyltransferases: Isoform selectivity and kinetics of 4-methylumbelliferone and 1-naphthol glucuronidation, effects of organic solvents, and inhibition by diclofenac and probenecid. Drug Metab. Dispos. 2004, 32, 413–423. [Google Scholar] [CrossRef]
- Tezuka, Y.; Stampoulis, P.; Banskota, A.J.; Awale, S.; Kadota, S.; Saiki, I. Constituents of the Vietnamese medicinal plant Orthosiphon staminues. Chem. Pharm.. Bull. 2000, 48, 1711–1719. [Google Scholar] [CrossRef]
- Alkharfy, K.M.; Frye, R.F. Effect of valerian, valerian/hops extracts, and valerenic acid on glucuronidation in vitro. Xenobiotica 2007, 37, 113–123. [Google Scholar] [CrossRef]
- Sridar, C.; Goosen, T.C.; Kent, U.M.; Williams, J.A.; Hollenberg, P.F. Silybin inactivates cytochromes P450 3A4 and 2C9 and inhibits major hepatic glucuronosyltransferases. Drug Metab. Dispos. 2004, 32, 587–594. [Google Scholar] [CrossRef]
- Katoh, M.; Yoshioka, Y.; Nakagawa, N.; Yokoi, T. Effects of Japanese herbal medicine, Kampo, on human UGT1A1 activity. Drug Metab. Pharmacokinet. 2009, 24, 226–234. [Google Scholar] [CrossRef]
- Soars, M.G.; Ring, B.J.; Wrighton, S.A. The effect of incubation conditions on the enzyme kinetics of UDP-glucuronosyltransferases. Drug Metab. Dispos. 2003, 31, 762–767. [Google Scholar] [CrossRef]
- Cheng, Z.; Radominska-Pandya, A.; Tephly, T.R. Cloning and expression of human UDP-glucuronosyltransferases 1A8. Arch. Biochem. Biophys. 1999, 356, 301–305. [Google Scholar]
- Williams, J.A.; Hyland, R.; Jones, B.C.; Smith, D.A.; Hurst, S.; Goosen, T.C.; Peterkin, V.; Koup, J.R.; Ball, S.E. Drug-drug interactions for UDP-glucuronosyltransferase substrates: A pharmacokinetic explanation for typically observed low exposure (AUCi/AUC) ratios. Drug Metab. Dispos. 2004, 32, 1201–1208. [Google Scholar] [CrossRef]
- Senafi, S.B.; Clarke, D.J.; Burchell, B. Investigation of the substrate specificity of a cloned expressed human bilirubin UDP-glucuronosyltransferase: UDP-sugar specificity and involvement in steroid and xenobiotic glucuronidation. Biochem. J. 1994, 303, 233–240. [Google Scholar]
- Liston, H.L.; Markowitz, J.S.; Devane, C.L. Drug glucuronidation in clinical psychopharmacology. J. Clin. Psychopharm. 2001, 21, 500–515. [Google Scholar] [CrossRef]
- Bernard, O.; Guillemette, C. The main role of UGT1A9 in the hepatic metabolism of mycophenolic acid and the effects of naturally occurring variants. Drug Metab. Dispos. 2004, 32, 775–778. [Google Scholar] [CrossRef]
- Calabrese, C.; Merman, S.H.; Babish, J.G.; Ma, X.; Shinto, L.; Dorr, M.; Wells, K.; Wenner, C.A.; Standish, L.J. A phase I trial of andrographolide in HIV posiive patients and normal volunteers. Phytother. Res. 2000, 14, 333–338. [Google Scholar] [CrossRef]
- Chang, J.; Zhang, R.M.; Zhang, Y.; Chen, Z.B.; Zhang, Z.M.; Xu, Q. Andrographolide drop-pill in treatment of acute upper respiratory tract infection with external wind-heat syndrome: a multicenter and randomized control trial. J. Chin. Integr. Med. 2008, 6, 1238–1245. [Google Scholar] [CrossRef]
- Jain, D.C.; Gupta, M.M.; Saxena, S.; Kumar, S. LC analysis of hepatoprotective diterpenoids from Andrographis paniculata. J. Pharm. Biomed. 2000, 22, 705–709. [Google Scholar] [CrossRef]
- Jin, C.-J.; Miners, J.O.; Lillywhite, K.J.; Mackenzie, P.I. Complementary deoxyribonucleic acid cloning and expression of a human liver uridine diphosphate –glucuronosyltransferase glucuronidating carboxylic acid-containing drugs. J. Pharmacol. Exp. Ther. 1993, 264, 475–479. [Google Scholar]
- Mojarrabi, B.; Butler, R.; Mackenzie, P.I. cDNA cloning and characterization of the human UDP glucuronosyltransferase UGT1A3. Biochem. Bioph. Res. Co. 1996, 225, 785–790. [Google Scholar] [CrossRef]
- Miners, J.O.; Valente, L.; Lillywhite, K.J.; Mackenzie, P.I.; Burchell, B.; Baguley, B.C.; Kestell, P. Preclinical prediction of factors influencing the elimination of 5,6-dimethylxanthenone-4-acetic acid, a new anticancer drug. Cancer Res. 1997, 57, 284–289. [Google Scholar]
- Mojarrabi, B.; Mackenzie, P.I. The human UDP glucuronosyltransferase UGT1A10 glucuronidates mycophenolic acid. Biochem. Bioph. Res. Co. 1997, 238, 775–778. [Google Scholar] [CrossRef]
- Mojarrabi, B.; Mackenzie, P.I. Characterization of two UDP- glucuronosyltransferases that are predominantly expressed in human colon. Biochem. Bioph. Res. Co. 1998, 240, 704–709. [Google Scholar] [CrossRef]
- Sorich, M.J.; Smith, P.A.; McKinnon, R.A.; Miners, J.O. Pharmacophore and quantitative structure-activity relationship modelling of UDP-glucuronosyltransferase 1A1 (UGT1A1) substrates. Pharmacogenetics 2002, 12, 635–645. [Google Scholar] [CrossRef]
- Stone, A.N.; Mackenzie, P.I.; Galetin, A.; Houston, J.B.; Miners, J.O. Isoform selectivity and kinetics of morphine 3- and 6-glucuronidation by human UDP-glucuronosyltransferases: Evidence of atypical glucuronidation kinetics by UGT2B7. Drug Metab. Dispos. 2003, 31, 1086–1089. [Google Scholar] [CrossRef]
- Miners, J.O.; Lillywhite, K.J.; Matthews, A.P.; Jones, M.E.; Birkett, D.J. Kinetic and inhibitor studies of 4-methylumbelliferone and 1-naphthol glucuronidation in human liver microsomes. Biochem. Pharmacol. 1988, 37, 665–671. [Google Scholar]
- Sample Availability: Samples of the compounds are available from the authors.
© 2010 by the authors;
Share and Cite
Ismail, S.; Aziah Hanapi, N.; Ab Halim, M.R.; Uchaipichat, V.; Mackenzie, P.I. Effects of Andrographis paniculata and Orthosiphon stamineus Extracts on the Glucuronidation of 4-Methylumbelliferone in Human UGT Isoforms. Molecules 2010, 15, 3578-3592. https://doi.org/10.3390/molecules15053578
Ismail S, Aziah Hanapi N, Ab Halim MR, Uchaipichat V, Mackenzie PI. Effects of Andrographis paniculata and Orthosiphon stamineus Extracts on the Glucuronidation of 4-Methylumbelliferone in Human UGT Isoforms. Molecules. 2010; 15(5):3578-3592. https://doi.org/10.3390/molecules15053578
Chicago/Turabian StyleIsmail, Sabariah, Nur Aziah Hanapi, Mohd Rohaimi Ab Halim, Verawan Uchaipichat, and Peter I. Mackenzie. 2010. "Effects of Andrographis paniculata and Orthosiphon stamineus Extracts on the Glucuronidation of 4-Methylumbelliferone in Human UGT Isoforms" Molecules 15, no. 5: 3578-3592. https://doi.org/10.3390/molecules15053578