Essential Oil Composition and Internal Transcribed Spacer (ITS) Sequence Variability of Four South-Croatian Satureja Species (Lamiaceae)

Abstract
:Introduction
Results and Discussion
Essential oil composition
Essential oils variability

{kind=link}
{kind=link}
| No. | Component | RI | S. montana year (yield %) 2007 (2.8) | S. cuneifolia year (yield %) 2007 (2.6) | S. subspicata year (yield %) 2007 (2.0) | S. visianii year (yield %) 2007 (2.4) |
|---|---|---|---|---|---|---|
| 1. | α-Thujene | 924 | - | - | - | 10.9 |
| 2. | α-Pinene | 935 | 0.9 | - | 24.2 | 5.8 |
| 3. | Camphene | 947 | - | - | - | 1.7 |
| 4. | Verbenene | 961 | - | - | 0.4 | |
| 5. | Sabinene | 971 | - | - | 1.7 | 1.4 |
| 6. | 1-Octene-3-ol | 974 | 1.1 | 1.4 | 1.8 | 1.3 |
| 7. | Myrcene | 988 | 4.3 | - | 2.1 | 1.6 |
| 8 | Linalool oxide | 991 | - | 0.3 | 1.3 | 0.2 |
| 9 | α-Terpinene | 1016 | - | - | 6.2 | - |
| 10. | p-Cymene | 1021 | 11.8 | 9.8 | t | 0.3 |
| 11. | Limonene | 1028 | 9.5 | 6.2 | 7.1 | 5.1 |
| 12. | (Z)-β-Ocimene | 1032 | - | - | 4.8 | 1.7 |
| 13. | γ-Terpinene | 1057 | 10.6 | 14.8 | - | 1.6 |
| 14. | n-Octanol | 1063 | - | 2.4 | 1.4 | 2.1 |
| 15. | Sabinene hydrate | 1065 | - | - | 1.2 | 0.3 |
| 16. | Methyl benzoate | 1088 | - | - | t | |
| 17. | Linalool | 1097 | 4.6 | 6.6 | 1.2 | 2.3 |
| 18. | allo-Ocimene | 1128 | 0.6 | 2.1 | 1.7 | 0.4 |
| 19. | Camphor | 1143 | 1.2 | - | - | 18.7 |
| 20. | Camphene hydrate | 1145 | - | - | - | 0.5 |
| 21. | Menthone | 1148 | - | - | - | 0.2 |
| 22. | Isoborneol | 1155 | 1.1 | 0.6 | 1.1 | 0.4 |
| 23. | Borneol | 1165 | 5.8 | 4.2 | 1.8 | - |
| 24. | Terpinen-4-ol | 1174 | 2.7 | 2.3 | 1.3 | 1.8 |
| 25. | α -Terpineol | 1186 | 1.3 | 0.8 | 1.8 | 5.1 |
| 26. | Myrtenol | 1194 | - | 2.1 | 1.3 | 0.8 |
| 27. | Nerol | 1227 | 0.7 | - | - | 1.4 |
| 28. | Geraniol | 1249 | 3.6 | 2.9 | 2.4 | 3.6 |
| 29. | Linalyl acetate | 1261 | 3.7 | 2.2 | 2.2 | 3.8 |
| 30. | Thujanol acetate | 1281 | - | - | - | 0.3 |
| 31. | Thymol | 1290 | 1.9 | 2.3 | 3.9 | 2.3 |
| 32. | Carvacrol | 1298 | 13.7 | 17.7 | 2.7 | 0.6 |
| 33. | α -Terpinyl acetate | 1316 | 1.9 | - | 1.5 | 0.3 |
| 34. | Neryl acetate | 1359 | - | 2.6 | 2.6 | 2.0 |
| 35. | α -Copaene | 1374 | - | 0.9 | 4.8 | 1.4 |
| 36. | Geranyl acetate | 1379 | 3.7 | 3.2 | 2.4 | 5.4 |
| 37. | β-Bourbonene | 1387 | 0.9 | 0.4 | 1.6 | 0.4 |
| 38. | Aromadendrene | 1439 | 0.3 | - | 0.1 | |
| 39. | α -Humulene | 1452 | 0.8 | - | - | 1.0 |
| 40. | (E)-β -Farnesene | 1454 | - | - | - | 0.4 |
| 41. | β -Caryophyllene | 1467 | 1.2 | 0.3 | - | 0.5 |
| 42. | Viridiflorene | 1496 | - | 0.3 | 2.3 | 0.2 |
| 43. | δ-Cadinene | 1522 | 2.3 | 2.9 | 4.8 | 0.1 |
| 44. | Spathulenol | 1578 | 1.0 | 0.3 | - | 0.9 |
| 45. | Caryophyllene oxide | 1582 | 1.6 | 0.2 | - | 1.6 |
| 46. | α -Eudesmol | 1652 | - | - | - | 0.6 |
| 47. | Elemol acetate | - | - | - | 0.2 | |
| 48. | Heptadecane | - | - | - | 0.5 | |
| Total: | 92.8 | 89.8 | 93.2 | 92.2 |
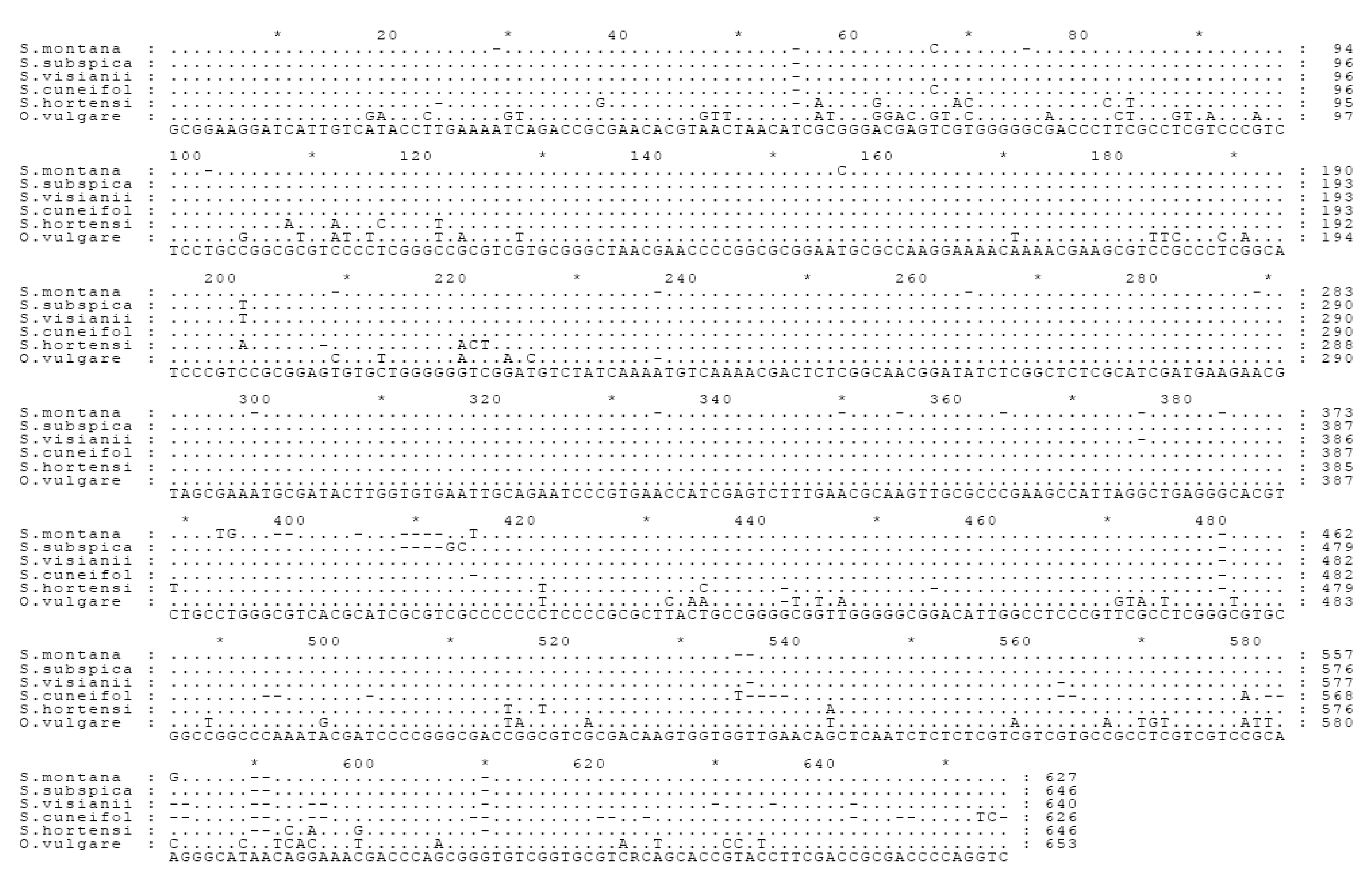
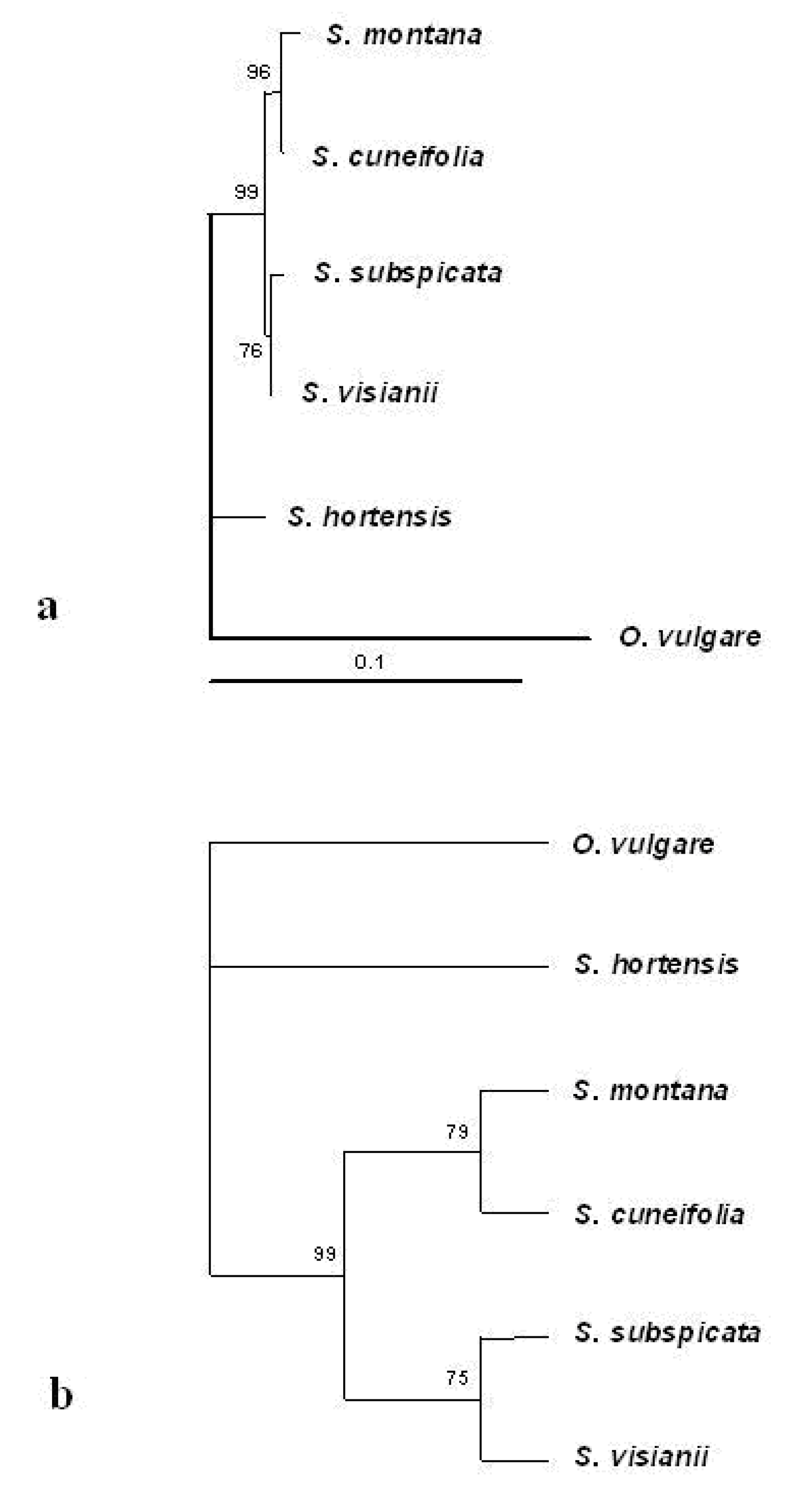
Internal transcribed spacer (ITS) sequence analysis
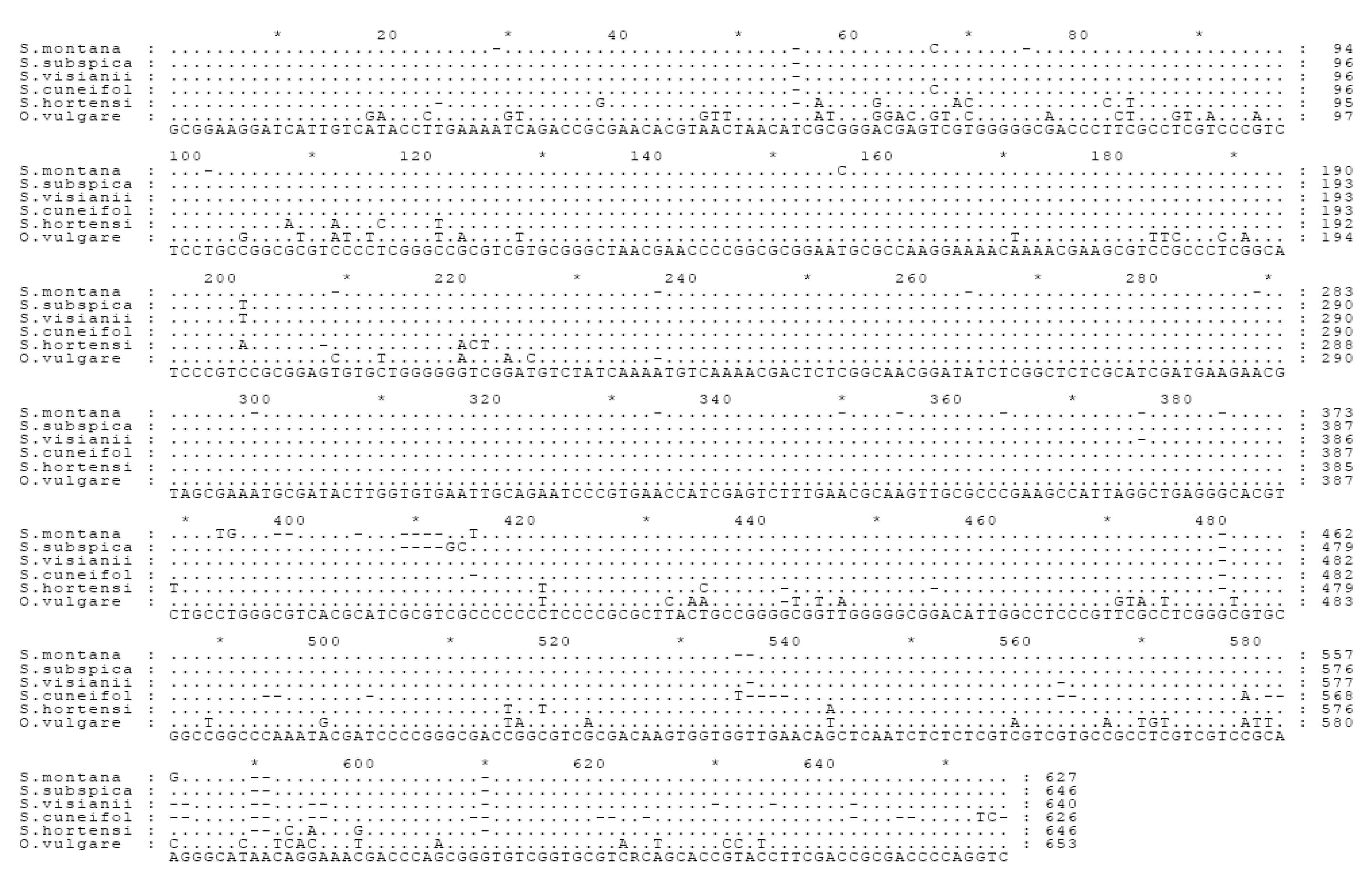

The comparison between the phytochemical and molecular DNA data
Conclusions
Experimental
Plant material
Isolation and GC-MS analysis of essential oils
DNA isolation, PCR amplification, cloning and sequencing
Sequence alignment and phenetic analysis
Acknowledgment
References
- Šilić, Č. Monographie der Gattungen Satureja L., Calamimtha Miller, Micromeria Bentham, Acinos Miller und Clinopodium L. In Der Flora Jugoslawiens; Zemaljski muzej BiH: Sarajevo, 1979; pp. 24–117. [Google Scholar]
- Pedersen, J. Distribution and taxonomic implications of some phenolics in the family Lamiaceae determined by ESR spectroscopy. Biochem. Syst. Ecol. 2000, 28, 229–253. [Google Scholar] [CrossRef]
- Güllüce, M.; Sökmen, M.; Daferera, D.; Ağar, G.; Ozkan, H.; Kartal, N.; Polissiou, M.; Sökmen, A.; Sahin, F. In vitro antibacterial, antifungal, and antioxidant activities of the essential oil and methanol extracts of herbal parts and callus cultures of Satureja hortensis L. J. Agric. Food Chem. 2003, 51, 3958–3965. [Google Scholar] [CrossRef]
- Gören, A.; Topçu, G.; Bilsel, G.; Bilsel, M.; Wilkinson, J.; Cavanagh, H. Analysis of essential oil of Satureja thymbra by hydrodistillation, thermal desorber, and headspace GC/MS techniques and its antimicrobial activity. Nat. Prod. Res. 2004, 18, 189–195. [Google Scholar] [CrossRef]
- Sahin, F.; Karaman, I.; Güllüce, M.; Oğütçü, H.; Sengül, M.; Adigüzel, A.; Oztürk, S.; Kotan, R. Evaluation of antimicrobial activities of Satureja hortensis L. J. Ethnopharmacol. 2003, 87, 61–65. [Google Scholar] [CrossRef]
- Skočibušić, M.; Bezić, N. Chemical Composition and Antimicrobial Variability of Satureja montana L. Essential Oils Produced During Ontogenesis. J. Essent. Oil Res 2004 a, 16, 387–391. [Google Scholar] [CrossRef]
- Skočibušić, M.; Bezić, N. Phytochemical analysis and in vitro antimicrobial activity of two Satureja species essential oils. Phytotherapy Res. 2004 b, 18, 967–970. [Google Scholar]
- Skočibušić, M.; Bezić, N.; Dunkić, V. Phytochemical composition and antimicrobial activities of the essential oils from Satureja subspicata Vis. growing in Croatia. Food Chem. 2006, 96, 20–28. [Google Scholar] [CrossRef]
- Cetojević-Simin, D.; Canadanović-Brunet, J.; Bogdanović, G.; Cetković, G.; Tumbas, V.; Djilas, S. Antioxidative and antiproliferative effects of Satureja montana L. extracts. JBUON. 2004, 9, 443–449. [Google Scholar]
- Amanlou, M.; Dadkhah, F.; Salehnia, A.; Farsam, H.; Dehpour, A. An anti-inflammatory and anti-nociceptive effects of hydroalcoholic extract of Satureja khuzistanica Jamzad extract. J. Pharm. Sci. 2005, 8, 102–106. [Google Scholar]
- Michaelakis, A.; Theotokatos, S.A.; Koliopoulos, G.; Chorianopoulos, N.G. Essential OIls of Satureja species: Insecticidal Effect on Culex pipiens Larvae (Diptera: Culicidae). Molecules 2007, 12, 2567–2578. [Google Scholar]
- Yamasaki, K.; Nakano, M.; Kawahata, T.; Mori, H.; Otake, T.; Ueba, N.; Oishi, I.; Inami, R.; Yamane, M.; Nakamura, M.; Murata, H.; Nakanishi, T. Anti-HIV-1 activity of herbs in Labiatae. Biol. Pharm. Bull. 1998, 21, 829–833. [Google Scholar] [CrossRef]
- Smith-Palmer, A.; Stewart, J.; Fyfe, L. Antimicrobial properties of plant essential oils and essences against five important food-borne pathogens. Lett. Appl. Microbiol. 1998, 26, 118–122. [Google Scholar]
- Razzaghi-Abyaneh, M.; Shams-Ghahfarokhi, M.; Yoshinari, T.; Rezaee, M.; Jaimand, K.; Nagasawa, H.; Sakuda, S. Inhibitory effects of Satureja hortensis L. essential oil on growth and aflatoxin production by Aspergillus parasiticus. Int. J. Food Microbiol. 2008, 123, 228–233. [Google Scholar] [CrossRef]
- Rota, C.; Carramiñana, J.; Burillo, J.; Herrera, A. In vitro antimicrobial activity of essential oils from aromatic plants against selected foodborne pathogens. J. Food Prot. 2004, 67, 1252–1256. [Google Scholar]
- Wink, M. Evolution of secondary metabolites from an ecological and molecular phylogenetic perspective. Phytochemistry 2003, 64, 3–19. [Google Scholar] [CrossRef]
- Oussalah, M.; Caillet, S.; Lacroix, M. Mechanism of action of Spanish oregano, Chinese cinnamon, and savory essential oils against cell membranes and walls of Escherichia coli O157:H7 and Listeria monocytogenes. J. Food Prot. 2006, 69, 1046–1055. [Google Scholar]
- Di Pasqua, R.; Betts, G.; Hoskins, N.; Edwards, M.; Ercolini, D.; Mauriello, G. Membrane toxicity of antimicrobial compounds from essential oils. J. Agric. Food Chem. 2007, 55, 4863–4870. [Google Scholar] [CrossRef]
- Turner, G.; Croteau, R. Organization of monoterpene biosynthesis in Mentha. Immunocytochemical localizations of geranyl diphosphate synthase, limonene-6-hydroxylase, isopiperitenol dehydrogenase, and pulegone reductase. Plant Physiol. 2004, 136, 4215–4227. [Google Scholar] [CrossRef]
- Turner, G.; Gershenzon, J.; Nielson, E.; Froehlich, J.; Croteau, R. Limonene synthase, the enzyme responsible for monoterpene biosynthesis in peppermint, is localized to leucoplasts of oil gland secretory cells. Plant Physiol. 1999, 120, 879–886. [Google Scholar] [CrossRef]
- Turner, G.; Gershenzon, J.; Croteau, R. Development of peltate glandular trichomes of peppermint. Plant Physiol. 2000, 124, 665–680. [Google Scholar]
- Dunkić, V.; Bezić, N.; Ljubešić, N.; Bočina, I. Glandular hair ultrastructure and essential oils in Satureja subspicata Vis. ssp. subspicata and ssp. liburnica Šilić. Acta Biol. Cracov. Series Bot. 2007, 49, 45–51. [Google Scholar]
- Hanlidou, E.; Kokkini, S.; Bosobalidis, A.M.; Bessiere, J.M. Glandular trichomes and essential oil constituents of Calamintha menthifolia (Lamiaceae). Plant Syst. Evol. 1990, 177, 17–26. [Google Scholar]
- Bezić, N.; Dunkić, V.; Radonić, A. Glandular apparatus structure and essential oil constituents of Satureja cuneifolia Ten. Acta Biol. Cracov. Ser. Bot. 2001, 43, 65–68. [Google Scholar]
- Kaufmann, M.; Wink, M. Molecular systematics of the nepetoideae (family Labiatae): phylogenetic implications from rbcL gene sequences. Z. Naturforsch. C 1994, 49, 635–645. [Google Scholar]
- Cantino, P.; Harley, R.; Wagstaff, S. Genera of Labiatae status and classification. In Advances in Labiatae Science; Harley, R., Reynolds, J., Eds.; Royal Botanical Gardens: Kew, Landon, UK, 1992; pp. 511–522. [Google Scholar]
- Sefidkon, F.; Jamazad, Z. Essential oil analysis of Iranian Satureja edmondi and S. isophylla. Flavour Fragr. J. 2006, 21, 230–233. [Google Scholar] [CrossRef]
- Kuštrak, D.; Kulftinec, J.; Blažević, N.; Maffei, M. Comparison of the Essential Oil Composition of Two Subspecies of Satureja montana. J. Essent. Oil Res. 1996, 8, 7–13. [Google Scholar] [CrossRef]
- Tümen, G.; Kirimer, N.; Ermin, N.; Başer, K. The essential oil of Satureja cuneifolia. Planta Med. 1998, 64, 81–83. [Google Scholar] [CrossRef]
- Azaz, D.; Demirci, F.; Satil, F.; Kürkçüoğlu, M.; Başer, K. Antimicrobial Activity of Some Satureja Essential Oils. Z. Naturforsch. 2002, 57, 817–821. [Google Scholar]
- Radonić, A.; Milos, M. Chemical composition and in vitro evaluation of antioxidant effect of free volatile compounds from Satureja montana L. Free Radic. Res. 2003, 37, 673–679. [Google Scholar] [CrossRef]
- Baydar, H.; Sağdic, O.; Özkan, G.; Karadoğan, T. Antibacterial activity and composition of essential oils from Origanum, Thymbra and Satureja species with commercial importance in Turkey. Food Contr. 2004, 15, 169–172. [Google Scholar] [CrossRef]
- Angelini, L.; Carpanese, G.; Cioni, P.; Morelli, I.; Macchia, M.; Flamini, G. Essential oils from Mediterranean Lamiaceae as weed germination inhibitors. J. Agric. Food Chem. 2003, 51, 6158–6164. [Google Scholar] [CrossRef]
- Akgul, A.; Ozcan, M.; Chialva, F.; Monguzzi, F. Essential oils of four Turkish wild-growing Labiatae herbs: Salvia cryptantha Montbr. Et Auch., Satureja cuneifolia Ten., Thymbra spicata L. and Thymus cilicicus Boiss. Et Bal. J. Essent. Oil Res. 1999, 11, 209–214. [Google Scholar] [CrossRef]
- Biavati, B.; Ozcan, M.; Piccaglia, R. Composition and antimicrobial properties of Satureja cuneifolia Ten. and Thymbra sintenisii Bornm. Et. Aznav. subsp. isaurica P.H. Davis essential oils. Ann. Microbiol. 2004, 54, 393–401. [Google Scholar]
- Bruneton, J. Pharmacognosy Phytochemistry Medical Plants.; Lavoiser Publishing Inc.: Paris, France, 1995; pp. 474–510. [Google Scholar]
- Glambiel, J.; Croteau, R. Biosynthesis of (±)-α-pinene and (-)-β-pinene from geranylpyrophsophate by a soluble enzyme system from sage (SaMa officinalis). J. Biol. Chem. 1982, 257, 2335–2342. [Google Scholar]
- Vigushin, D.; Poon, G.; Boddy, A.; English, J.; Halbert, G.; Pagonis, C.; Jarman, M.; Coombes, R. Phase I and pharmacokinetic study of D-limonene in patients with advanced cancer. Cancer Research Campaign Phase I/II Clinical Trials Committee. Cancer Chem. Pharmacol. 1998, 42, 111–117. [Google Scholar] [CrossRef]
- Hebert, P.; Ratnasingham, S.; deWaard, J. Barcoding animal life: cytochrome c oxidase subunit 1 divergences among closely related species. Proc. R. Soc. B 2003, 270 Suppl 1, S96–S99. [Google Scholar]
- Bakker, F.T.; Culham, A.; Pankhurst, C.E.; Gibby, M. Mitochondrial and chloroplast DNA-based phylogeny of Pelargonium (Geraniaceae). Am. J. Bot. 2000, 87, 727–734. [Google Scholar] [CrossRef]
- Alvarez, I.; Wendel, J. Ribosomal ITS sequences and plant phylogenetic inference. Mol. Phylog. Evol. 2003, 29, 417–434. [Google Scholar] [CrossRef]
- Kress, W.; Wurdack, K.; Zimmer, E.; Weigt, L.; Janzen, D. Use of DNA barcodes to identify flowering plants. Proc. Nat. Acad. Sci. USA 2005, 102, 8369–8374. [Google Scholar] [CrossRef]
- Adams, R.P. Identification of essential oil components by gas chromatography/ mass spectroscopy; Fourth ed. Allured Publishing Corp.: Carol Stream IL, USA, 2007. [Google Scholar]
- Hall, T.; Ibis Biosciences, North Carolina State University, NC, USA. BioEdit v5.0.9, 1999.
- Swofford, D. PAUP, Phylogenetic Analysis Using Parsimony (*and or Other Methods), Version 4. Sinauer Assoc.: Sunderland, MA, USA, 1998. [Google Scholar]
- Kimura, M. Rare variant alleles in the light of the neutral theory. Mol. Biol. Evol. 1983, 1, 84–93. [Google Scholar]
- Sample Availability: Available from the authors.
© 2009 by the authors; licensee Molecular Diversity Preservation International, Basel, Switzerland. This article is an open-access article distributed under the terms and conditions of the Creative Commons Attribution license ( http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Bezić, N.; Šamanić, I.; Dunkić, V.; Besendorfer, V.; Puizina, J. Essential Oil Composition and Internal Transcribed Spacer (ITS) Sequence Variability of Four South-Croatian Satureja Species (Lamiaceae). Molecules 2009, 14, 925-938. https://doi.org/10.3390/molecules14030925
Bezić N, Šamanić I, Dunkić V, Besendorfer V, Puizina J. Essential Oil Composition and Internal Transcribed Spacer (ITS) Sequence Variability of Four South-Croatian Satureja Species (Lamiaceae). Molecules. 2009; 14(3):925-938. https://doi.org/10.3390/molecules14030925
Chicago/Turabian StyleBezić, Nada, Ivica Šamanić, Valerija Dunkić, Višnja Besendorfer, and Jasna Puizina. 2009. "Essential Oil Composition and Internal Transcribed Spacer (ITS) Sequence Variability of Four South-Croatian Satureja Species (Lamiaceae)" Molecules 14, no. 3: 925-938. https://doi.org/10.3390/molecules14030925